| ← |
«Секс на R» |
→ |
| Секс на R–08: «Стать – королева проблем еволюційної біології» (Опис проблеми II) | Секс на R–09: Моделювання парадокса Мейнарда Сміта (Завдання I) |
Секс на R–10: Моделювання циклічно змінюваного добору за Стертевантом та іншими (Завдання II) |
Моделювання парадокса Мейнарда Сміта
Крок 1
В першій лекції, присвяченій проблемі еволюції сексу, пояснено проблему, яку можна назвати «парадоксом Мейнарда Сміта» (слайди 30–32). Цей парадокс полягає в тому, що поява партеногенетичних самиць в популяціях роздільностатевих організмів мала б призвести до швидкої відмови від сексуального відтворення. Чому ж це не відбувається?
Завдання 1: побудова моделі парадокса Мейнарда Сміта I типу. Побудуйте у R модель, яка дослідить динаміку популяції, яка складається з самиць, самців і клональних особин (прикладом яких можуть бути партеногенетичні самиці). Вхідними параметрами цієї моделі мають бути:
— початковий склад популяції (окремо чисельність кожної категорії особин: самиць, самців та партеногенетиків);
— кількість і склад потомства у тих особин, що його безпосередньо залишають;
— верхня межа чисельності популяції.
Можливі різні механізми скорочення чисельності цієї популяції (у разі, якщо вона перевищує верхню межу); на першому кроці можна обрати найпростіший з них. На кожному циклі цієї моделі має відбуватися розмноження, а потім - скорочення чисельності особин. Виходом має стати динаміка чисельності різних форм з часом.
Цю модель можна зробити детерміністською (наприклад, завжди потомство самиці — парне, наполовину самиці, наполовину самці), а можна — ймовірнісним. Цікавіше зробити модель ймовірнісною. У такому разі краще обрати механізм ймовірнісного округлення чисельності.
Наступна вимога не є обов'язковою, але усе ж таки краще зробити модель відповідно до типової структури моделей у курсі. Відповідну чернетку R-скрипту можна завантажити тут. Зрозуміло, що починати краще з моделі I типу, тобто такої моделі, яка проводить розрахунок одного причинно-наслідкового ланцюжка, отримує певний варіант динаміки складу модельної популяції за заданих умов.
Спробуємо описати один з варіантів побудови такої моделі (тобто розробити концептуальну модель). Можна обрати один з двох варіантів. За першого варіанта партеногенетики є в модельній популяції з початку (але, скоріше за все, у невеликій чисельності). За другого варіанта на початку імітації партеногенетиків в популяції нема, але вони можуть з невеликою ймовірністю в ній з'явитися. Цей варіант цілком відповідає наявним даним, що були отримані при дослідженні дійсних популяцій багатьох видів тварин. Цікавішим є другий варіант; оберемо саме його.
Концептуальна модель парадокса Мейнарда Сміта (типу I):
— модельна популяція складається з трьох груп особин: самиць (їх у моделі можна позначити 1), самців (2) та партеногенетиків (3);
— початкову (α) чисельність (N) самиць (fe) на циклі t можна позначити αtNfe; аналогічно можна використати позначення αtNma та αtNpg;
— кількості особин певної статі на початку імітації належить до вхідних параметрів; ці кількості можна позначити α0Nfe, α0Nma та α0Npg;
— внаслідок розмноження відбувається перехід від альфа-популяції до бета-популяції;
— у самиць (при наявності в популяції самців) з'являється потомство, що складається з самиць та самців; у партеногенетиків — потомство з партеногенетиків;
— слід припустити, що з невеликою ймовірністю у потомстві самиці може випадково з'явитися партеногенетик;
— кількість потомків певної статі у певних батьків належить до вхідних параметрів; так, кількість самиць у потомстві самиці можна позначити feofFe (перша f позначає, що самиця має бути заплідненою, фертилізованою, і для цього необхідною є наявність самців);
— інші позначення вхідних параметрів, що характеризують плодючість: maofFe, pgofFe, pgoPg; партеногенетики не потребують самців для запліднення;
— бета-чисельність кожної групи вираховується як сума альфа-чисельності та приплоду (з ймовірнісним округленням до цілих): βtNpg = αtNpg + αtNfe×pgofFe + αtNpg×pgoPg;
— верхня межа чисельності популяції (K) є одним з вхідних параметрів моделі;
— якщо сумарна бета-чисельність популяції (βtN = βtNfe + βtNma +βtNpg) перевищує верхню межу, відбувається скорочення чисельності при переході до омега-чисельності (якщо не перевищує — омега-склад популяції є ідентичним бета-складу);
— скорочення відбувається так: кожна особина виживає з ймовірністю K / βtN та гине з ймовірністю 1 - K / βtN;
— альфа-склад популяції (альфа-чисельність кожної групи) наступного циклу визначається омега-складом популяції на попередньому етапі: αt+1Nfe = ωtNfe, αt+1Nma = ωtNma, αt+1Npg = ωtNpg.
Одним з варіантів початкових параметрів та умов, що відповідає описаній концептуальній моделі, є такий.
# I.2. Initial state of the system — початковий стан модельованої системи
al_0_N_fe <- 50 # Початкова чисельність самиць
al_0_N_ma <- 50 # Початкова чисельність самців
al_0_N_pg <- 0 # Початкова чисельність партеногенетиків
# I.3. Parameters — параметри
K <- 100 # Обмеження ємності середовища
fe_o_fFe <- 1.99 # кількість самиць в потомстві фертильної (при наявності самця) самиці
ma_o_fFe <- 2 # кількість самців в потомстві фертильної (при наявності самця) самиці
pg_o_fFe <- 0.01 # кількість партеногенетиків в потомстві фертильної (при наявності самця) самиці
pg_o_Pg <- 4 # кількість партеногенетиків в потомстві партеногенетика
# I.4. Experimental conditions — умови експерименту з моделлю
cycles <- 50 # Кількість робочих циклів моделіРезультатом виконання першого завдання може бути наступний графік.
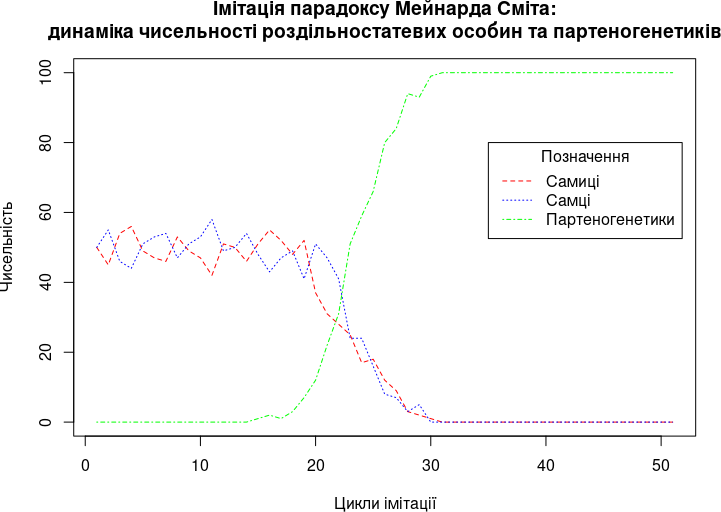
Крок 2
Після того, як описана в завданні 1 модель створена, можна побудувати на її основі моделі II та III типів.
Завдання 2: побудова моделі парадокса Мейнарда Сміта II типу. Побудуйте у R модель, яка проводить низку ітерацій (повторів з однаковими початковими параметрами та умовами).
Результатом виконання другого завдання може бути наступний графік.
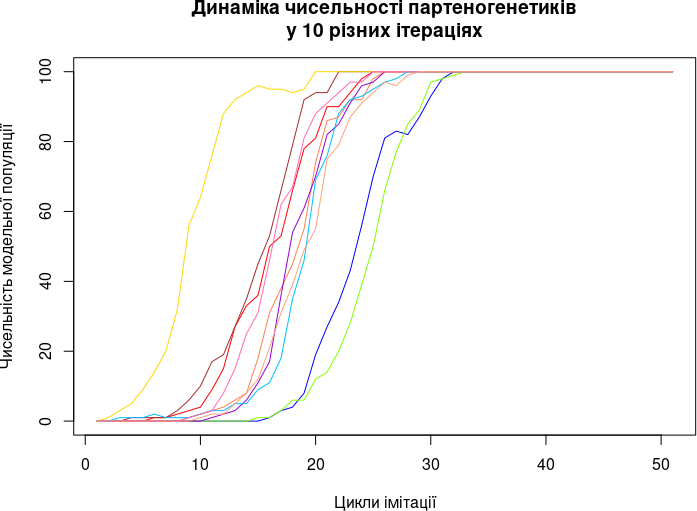
Завдання 3: побудова моделі парадокса Мейнарда Сміта III типу. Побудуйте у R модель, яка перевіряє, за яких значень плодючості самиць та партеногенетиків відбувається витіснення роздільностатевих особин клональними, а за яких — не відбувається.
Крок 3
За перевірених умов партеногенетики витісняють роздільностатевих особин. Це відбувається у разі, якщо:
— партеногенетики з'являються у роздільностатевій популяції;
— кількість потомків у партеногенетика не менша, ніж у самиці (або хоча б перевищує половину кількості потомків самиці);
— смертність партеногенетиків та роздільностатевих особин є однаковою.
Скоріше за все, у багатьох випадках перелічені умови виконуються. Чому ж роздільностатеві особини не виникли? Ймовірно, є якась причина, яка не була врахована в нашій моделі. Ми можемо її умовно назвати «прихованим фактором Мейнарда Сміта». Спробуємо його знайти!
Завдання 4: побудова концептуальної моделі механізму, що запобігає витісненню роздільностатевих особин клональними. Запропонуйте гіпотезу, що саме утримує популяції роздільностатевих організмів, захищає роздільностатевих особин від витіснення з боку клональних. Те ж саме можна сформулювати так: припустить, що саме є «прихованим фактором Мейнарда Сміта», як він працює!
Може, запропоновану вами гіпотезу можна буде перевірити завдяки побудові R-моделі (а може й ні...). Щоб спробувати це зробити, слід спочатку розробити концептуальну модель. Запропонуйте її!