
БиоСтатистика — 15. Тема 11. Некоторые методы, характерные для зоологии и экологии
В зоологии и экологии используется ряд специфических способов обработки данных. Некоторые из них обсуждаются в этом разделе

←
D.A. Shabanov, M.A. Kravchenko. Статистичний аналіз даних у зоології та екології
→
Тема 10. Дискримінантний аналіз
Тема 11. Деякі методи, характерні для зоології та екології
Додаток. Програма розділу великої практики
Біостатистика-14
Біостатистика-15
Біостатистика-16
Тема 11. Деякі методи, характерні для зоології та екології
11.1. Аналіз флуктуючої асиметрії
Тут ми розглянемо один із чудових методів, який може використовуватись у зоологічних та екологічних дослідженнях, і, звичайно, не лише в них. У цьому методі поєднуються простота збору даних, потенційна потужність… і дуже нетривіальні обставини, які треба враховувати при інтерпретації отриманих результатів.
Всім відомо, що у визначенні успадкованості багатьох ознак дуже ефективним виявляється близнюковий метод. Якщо ми хочемо визначити, який внесок вроджених факторів у розвитку якоїсь ознаки у людини, можна порівняти мінливість цієї ознаки у однояйцевих близнюків (клонів) і просто у братів і сестер (сибс). З іншого боку, порівнюючи мінливість за цікавлячою нас ознакою у клонів, що розвивалися в різному середовищі, ми можемо оцінити, наскільки середові впливи сприяли або заважали реалізації генетичної програми, що керує розвитком цієї ознаки.
У випадках, коли можна отримати генетично ідентичне потомство, порівняння результатів його розвитку виявляється дуже цікавим методом як для оцінки стійкості системи управління онтогенезом (при порівнянні розвитку різних ознак в одних умовах), так і для оцінки якості середовища, у якому відбувається розвиток (при порівнянні розвитку одних і тих самих ознак в різних умовах). Ту форму мінливості, яка вивчається в таких дослідженнях, Б. Л. Астауров назвав реалізаційною мінливістю — відмінностями між різними результатами реалізації однієї й тієї ж успадкованої програми.
Ідея, яка тут описується, полягає в тому, що тіло білатерально симетричних організмів можна розглядати як два «клони» — праву і ліву половини. Якщо програма розвитку для правої і лівої половини тіла ідентична, генетичний контроль розвитку розглянутих ознак стійкий, а середовище — сприятливе, результат розвитку буде повністю симетричним. Однак треба враховувати, що асиметрія може бути наслідком не лише реалізаційної мінливості, вона може бути адаптацією до тих чи інших факторів. Наприклад, у омарів одна клешня є більш сильною і має тупий край, оскільки переважно використовується для дроблення раковин і панцирів, а інша — менш сильна, але гостра, використовується для відрізання шматків відносно м’якої їжі.
Американський біолог Лей Ван-Вален запропонував класифікацію форм білатеральної асиметрії, показану на рисунку. Щоб визначити, з яким типом асиметрії ми маємо справу, треба побудувати розподіл відхилень від симетрії вправо (з більшим розвитком ознаки на правій стороні) або вліво.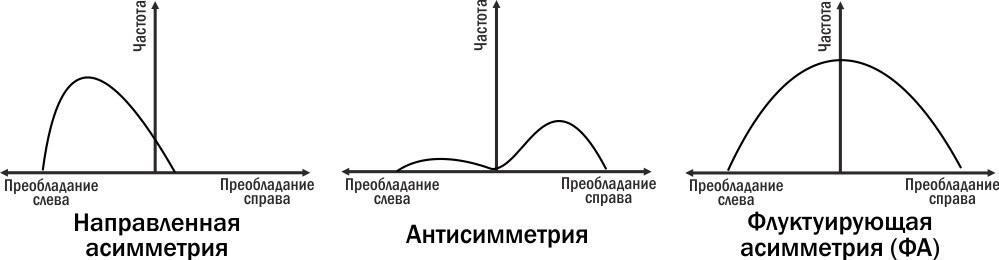
Рис. 11.1.1. Три типи асиметрії за Леєм Ван-Валеном (1962). Для людини прикладом направленої асиметрії може бути розташування серця, антисиметрії — розміри правої і лівої кисті, ФА — розміри правої і лівої вушної раковини
У випадку направленої асиметрії реєструється систематичне переважа́ння ознаки на одній стороні. Наприклад, серце людини закладається симетрично, але одна його половина виявляється більшою за іншу. Ліве серце викачує кров у великий круг кровообігу, праве — у малий. Серце практично завжди зміщене вліво. У випадку антисиметрії можуть спостерігатися як відхилення в одну, так і в іншу сторону, і ці відхилення зустрічаються частіше, ніж симетричний стан ознак. Наприклад, у правшів розмір кисті правої руки більший, у лівшів — лівої. Найрідкіснішою виявляється однаковість розвитку обох рук. Клешні омарів також є прикладом антисиметрії.
У випадку флуктуючої асиметрії (ФА) найчастішим станом білатеральної ознаки є її симетричність; сильні відхилення від симетричності спостерігаються рідше, ніж слабкі. Проаналізувавши розміри правої і лівої вушної раковини у людей, можна побачити, що відхилення у бік більшого розміру правого або лівого вуха мають приблизно такий характер. Саме такі ознаки з таким розподілом можна намагатися використовувати для оцінки реалізаційної мінливості. Отже, ФА — це випадкові, ненаправлені відхилення від білатерально симетричного стану. ФА можна використовувати для оцінки стійкості розвитку і впливу на неї різноманітних внутрішніх і зовнішніх факторів.
Прийнято вважати, що ФА зменшують (і, відповідно, підвищують стійкість розвитку):
– оптимальні умови;
– відносно чисте середовище;
– висока пристосованість особини;
– оптимальна генетична відстань між батьками;
– еволюційна консервативність, хороша регульованість розвитку вивчених ознак.
Підвищують ФА і знижують стійкість розвитку:
– екстремальні умови;
– забруднене середовище;
– низька пристосованість особини;
– інбридинг або віддалена гібридизація;
– несуттєвість, еволюційна новизна, нестійка регуляція розвитку розглянутих ознак.
Вивчення ФА належить до числа методів, де за значенням одного інтегрального ознаки дослідник намагається судити про весь комплекс факторів. Такі роботи вимагають високої ретельності в інтерпретації, уваги до дизайну дослідження. Хороша програма дослідження ФА міститься в цитованій далі статті.
«У статистичному аналізі ФА білатеральних ознак можна виділити два аспекти. Перший з них пов’язаний з аналізом індивідуальних ознак і включає:
— вивчення направленості (ненаправленості) асиметрії ознаки;
— вивчення залежності величини асиметрії ознаки (L‑R) від величини (розміру) ознаки на обох сторонах тіла (L+R) або (L+R)/2;
— вивчення ступеня корельованості величини асиметрії різних ознак, що використовуються в інтегральній оцінці ФА організму;
— вивчення наявності (відсутності) статевих (гендерних) різниць асиметрії ознак;
— вивчення внеску помилки вимірювання ознаки у кінцеву оцінку ФА (важливо для пластичних ознак і неактуально для меристичних).
Другий аспект пов’язаний з вибором і коректним застосуванням інтегральних показників, що оцінюють ФА вибірки організмів за комплексом індивідуальних ознак у задачах біомоніторингу». Гелашвілі Д. Б., Якимов В. Н., Логінов В. В., Єпланова Г. В. Статистичний аналіз флуктуючої асиметрії білатеральних ознак різнокольорової ящурки Eremias arguta // Актуальні проблеми герпетології та токсикології: Збірник наукових праць. Вип. 7. — Тольятті, 2004. — С. 45–59.
Отже, для дослідження ФА слід, перш за все, вибрати білатеральні ознаки, мінливість яких можна виміряти кількісно. Слід виміряти значення цих ознак на правій і на лівій стороні тіла у достатньо чисельної вибірки вивчених організмів. Щоб зробити позначення зрозумілішими, зазначимо, що вони стосуються ознаки, яку позначимо C (character). У такому випадку, значення цієї ознаки на правій і на лівій стороні можна позначити як C_r і C_l відповідно.
Важливим кроком вивчення ознаки, яку планується використовувати як міру ФА, є побудова її розподілу з урахуванням знаку. Слід обчислити величину D_C = C_r – C_l і визначити, чи відповідає її розподіл характеру, очікуваному для ФА. Гелашвілі та співавтори у процитованій вище статті радять оцінити нормальність/ненормальність ознаки, позначеної нами як D_C, за допомогою критеріїв Колмогорова‑Смирнова, Шапіро‑Уїлка, Лілієфорса. Ці критерії слугують для перевірки нульової гіпотези про відсутність відмінностей спостережуваного розподілу від нормального. Значущі відмінності свідчать про ненормальність розподілу. Однак слідування цій рекомендації призведе до того, що чим більшою буде вивчена вибірка, тим імовірніше буде значне відхилення її розподілу від нормального. На нашу думку оптимальний спосіб дій інший. Слід побудувати розподіл ознаки D_C і хоча б візуально оцінити, чи має він колоколообразну форму, чи ні. Якщо найчастішим станом ознаки є симетричність і чим сильніше відхилення від симетрії, тим рідше воно зустрічається, цю ознаку можна використовувати для оцінки ФА. У випадку малої вибірки емпіричний розподіл буде «нерівним»; у такому випадку можна застосовувати статистичні критерії, щоб оцінити значимість його відхилення від нормального. Якщо вибірок достатньо великі, розподіл відносно рівномірний і колоколообразний, гострої необхідності у використанні статистичних критеріїв, ймовірно, немає.
Розподіл асиметричності білатеральних структур може мати нетривіальний характер. Нижче наведено розподіл річкових окунів за симетричністю/асиметричністю кількості лусок у боковій лінії справа і зліва (за роботою: Виноградова К. П., Сакун Ю. В., Белоусова К. М., Гончаров Г. Л., Шабанов Д. А. Вивчення флуктуючої асиметрії річкового окуня (Perca fluviatilis L., 1758) // Біологія та валеологія, 2012. — Вип. 14. — С. 9‑17). Увагу привертає характерна форма розподілу у вигляді тризубця: відмінності між правою і лівою стороною тіла на одну луску зустрічаються рідше, ніж на дві (з 256 окунів відхилення на одну луску мали 72 особи, а на дві — 91 особа; при порівнянні відмінностей за Фішером з використанням одностороннього критерію переважає відхилення на дві луски, p=0,04). 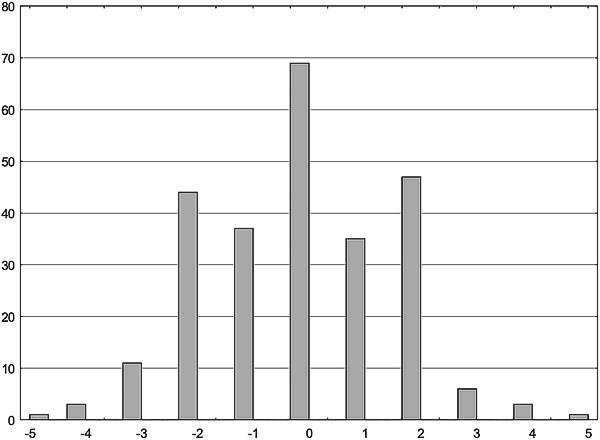
Рис. 11.1.2. Розподіл окунів за асиметрією кількості лусок вздовж бокової лінії справа і зліва. По осі абсцис — різниця в кількості лусок справа і зліва, по осі ординат — кількість спостережень (Виноградова К. П., Сакун Ю. В., Белоусова К. М., Гончаров Г. Л., Шабанов Д. А. Вивчення флуктуючої асиметрії річкового окуня (Perca fluviatilis L., 1758) // Біологія та валеологія, 2012. — Вип. 14. — С. 9‑17)
Як пояснити відображену на рис. 11.1.2 особливість розподілу? За версією, викладеною у процитованій статті, це може бути пов’язано з тим, що ті особи, відхилення яких від симетричності є відносно незначними, можуть регулювати свій розвиток, повертаючи його до норми. Якщо рівень відхилень, які є причиною ФА (тобто «онтогенетичного шуму», спонтанних відхилень, а також середових збурень) є низьким, епігенетичні механізми, що керують розвитком, можуть забезпечити симетричний розвиток. Після певного порогу ймовірність відхилень від симетричного стану зростає.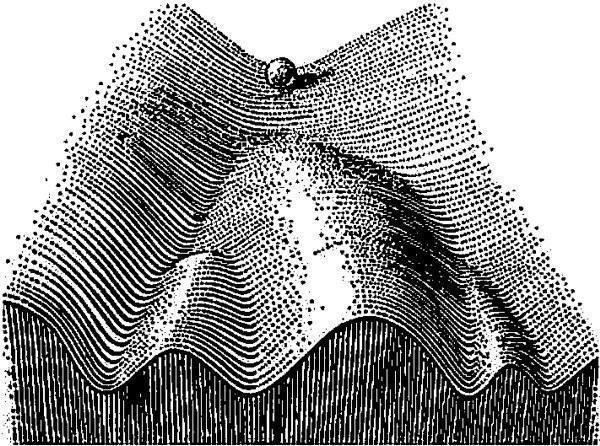
Рис. 11.1.3. Модель епігенетичного ландшафту, запропонована К. Х. Воддінгтоном. Онтогенез порівнюється в цій моделі зі скочуванням кульки по поверхні складної форми. Більш імовірним станам розвиваючоїся системи відповідають западини на поверхні. Невеликі відхилення від нормальної траєкторії розвитку внаслідок здатності системи до саморегуляції будуть виправлені; більш суттєві відхилення призведуть до зміни кінцевого стану розвиваючоїся системи
Епігенетичні механізми (дія яких у метафорі епігенетичного ландшафту виражається у западинах траєкторій нормального розвитку) стабілізують розвиток, а середові збурення і онтогенетичний шум — дестабілізують. З цієї точки зору такий розподіл, як показано на рис. 11.1.2, не суперечить припущенню, що він породжений ФА. Отже, асиметрія за кількістю лусок у окунів може бути використана для оцінки стійкості їхнього розвитку.
Наступний пункт програми обробки результатів дослідження ФА, запропонованої у статті Гелашвілі та співавторів, — перевірка того, чи направлена асиметрія. Для відповіді достатньо порівняти значення стовпців C_r і C_l за допомогою критерію Вілкоксона (непараметричного методу парних порівнянь, Statistics / Nonparametrics / Comparing two dependent samples (variables)). Можна припустити, що у випадку дослідження по‑справжньому великої вибірки і в даному випадку ймовірна реєстрація значущої асиметрії розподілу. На нашу думку, у випадку незначних відхилень від симетричності розподілу відхилень, аналізована ознака все ж можна використовувати для оцінки ФА, але при інтерпретації результатів не забувати про цю обставину. У деяких випадках можна обчислити відхилення не від симетричного стану, а від медіанного значення розподілу ознаки D_C.
Ще один етап аналізу даних за ФА — оцінка розмірної залежності. Для цього слід обчислити коефіцієнт кореляції між двома величинами: абсолютним значенням асиметрії, A_C = Abs(C_r – C_l), і середнім значенням ознаки C_mean = (C_r + C_l)/2. Якщо величина асиметрії не пов’язана з розміром вивчених структур, у якості міри ФА можна використовувати абсолютне значення асиметрії, A_C. Якщо з збільшенням розміру організму і вивчених структур зростає і абсолютне значення асиметрії, для оцінки ФА слід використовувати нормовану асиметрію: N_C = 2×Abs(C_r – C_l)/(C_r – C_l).
Наступний етап «програми» Гелашвілі та співавторів пов’язаний з визначенням того, наскільки корелюють між собою різні міри асиметрії, які можна використовувати при вивченні одного і того ж організму. Автори обговорюваної статті радять використовувати міри, які слабо пов’язані між собою. «Очевидно, що чим менш корельовані величини асиметрії ознак, тим більш незалежну і об’єктивну оцінку можна отримати за допомогою даного набору ознак» (Гелашвілі та ін., 2004). З іншого боку, міри ФА цікаві не самі по собі, а як способи оцінки стійкості/нестійкості розвитку організму. Стійкість розвитку організму (наприклад, «глибину» креодів, онтогенетичних траєкторій, показаних на рис. 11.1.3, безпосередньо оцінити неможливо). Ті ознаки, які ми використовуємо в ході дослідження, мають бути пов’язані з тією величиною, яку ми оцінюємо їх допомогою. З цієї точки зору, стійкість розвитку оптимально оцінювати за кількома ознаками, пов’язаними з нею, а отже — і пов’язаними між собою.
Якщо вибірка складається з підвибірок (наприклад, особин різної статі), слід перевірити, чи однорідні підвибірки за асиметричністю розглянутих ознак. Якщо частини вибірки однорідні, їх можна розглядати спільно (наприклад, об’єднати дані про самок і самців).
При використанні кількох білатеральних ознак для оцінки ФА можна застосовувати різні інтегральні міри. Найпростіша з них — просте підсумовування значень (абсолютних або нормованих) асиметрії всіх використаних ознак.
11.2. Приклад обробки даних про флуктуючу асиметрію
Розглянемо аналіз ФА на прикладі. Наведені тут дані отримані студентами II курсу під час практики з зоології хребетних у Гайдарах і опубліковані у цитованій вище статті (Виноградова К. П., Сакун Ю. В., Белоусова К. М., Гончаров Г. Л., Шабанов Д. А. Вивчення флуктуючої асиметрії річкового окуня (Perca fluviatilis L., 1758) // Біологія та валеологія, 2012. — Вип. 14. — С. 9‑17). Тут наведено не всі дані, а лише їх частину, відредаговану так, щоб при її обробці отримувався той самий результат, що і при обробці оригінальних даних. Роботи з аналогічним дизайном виконувалися і пізніше, і на окунях, і на інших видах риб; обробка зібраних у них даних привела до аналогічних висновків.
Вивчена вибірка річкових окунів. У кожної риби вимірювали довжину тіла (L_c) і визначали вік (за лініями на лусках). Наведено дані про першорічки, позначені I (0+), і другорічки, II (1+). Позначення пов’язані з тим, що у першорічок на лусках ще немає ліній, які утворюються під час зимувань, а у риб другого року життя є одна така лінія. Риби обох вікових груп розподілені на три групи, що відрізняються швидкістю їх росту. Це можливо, бо нерест окунів проходить у досить стислі терміни, і різниця в розмірі риб пов’язана перш за все не з різницею у віці в кілька днів, а з різною швидкістю росту. Повільно зростаючі особини позначені Slow, риби зі середньою швидкістю росту — Moderate, а швидко зростаючі — Fast.
У вивчених окунів визначали три білатеральні ознаки: кількість лусок вздовж бокової лінії (S), кількість променів у грудних (P) і у черевних плавниках (V). Позначення «_r» відповідає значенню ознаки на правій стороні тіла, а «_l» — на лівій.
Таблиця 11.1. Результати опису 100 річкових окунів (за Виноградова та ін., 2012, скорочений і відредагований варіант)
L_c
Age
Growth
S_r
S_l
P_r
P_l
V_r
V_l
L_c
Age
Growth
S_r
S_l
P_r
P_l
V_r
V_l
31
I (0+)
Slow
69
68
10
10
6
5
74
II (1+)
Slow
70
69
10
11
6
6
31
I (0+)
Slow
66
64
10
10
6
6
74
II (1+)
Slow
69
68
9
10
6
6
31
I (0+)
Slow
62
64
10
10
6
6
74
II (1+)
Slow
63
61
11
11
6
6
31
I (0+)
Slow
66
68
10
10
6
6
74
II (1+)
Slow
67
65
12
11
6
6
32
I (0+)
Slow
66
65
11
10
6
6
75
II (1+)
Slow
71
70
11
11
6
6
32
I (0+)
Slow
63
65
10
10
6
6
76
II (1+)
Slow
69
67
11
11
6
6
32
I (0+)
Slow
64
62
10
9
6
5
77
II (1+)
Slow
73
73
11
11
6
6
33
I (0+)
Slow
69
67
10
10
6
6
77
II (1+)
Slow
68
67
11
10
6
6
35
I (0+)
Slow
70
69
10
10
6
6
77
II (1+)
Slow
68
66
10
11
6
6
35
I (0+)
Slow
70
67
9
10
5
7
78
II (1+)
Slow
72
72
11
10
6
6
36
I (0+)
Slow
61
63
9
10
5
6
78
II (1+)
Slow
70
68
11
11
6
6
36
I (0+)
Slow
66
68
10
11
5
6
79
II (1+)
Slow
72
72
10
10
6
6
36
I (0+)
Slow
60
63
11
10
6
6
80
II (1+)
Slow
73
73
11
10
6
6
37
I (0+)
Slow
65
66
10
10
6
6
80
II (1+)
Slow
67
65
10
11
6
5
37
I (0+)
Slow
66
66
11
9
6
6{"translated":"81\n\nII (1+)\n\nSlow\n\n70\n\n70\n\n11\n\n10\n\n6\n\n6\n\n\n\n37\n\nI (0+)\n\nSlow\n\n56\n\n57\n\n12\n\n11\n\n6\n\n6\n\n83\n\nII (1+)\n\nModerate\n\n69\n\n68\n\n11\n\n11\n\n6\n\n6\n\n\n\n38\n\nI (0+)\n\nModerate\n\n65\n\n66\n\n9\n\n9\n\n6\n\n6\n\n83\n\nII (1+)\n\nModerate\n\n62\n\n65\n\n11\n\n11\n\n6\n\n6\n\n\n\n38\n\nI (0+)\n\nModerate\n\n71\n\n70\n\n10\n\n10\n\n6\n\n6\n\n83\n\nII (1+)\n\nModerate\n\n62\n\n60\n\n10\n\n10\n\n6\n\n6\n\n\n\n38\n\nI (0+)\n\nModerate\n\n67\n\n69\n\n10\n\n9\n\n6\n\n6\n\n84\n\nII (1+)\n\nModerate\n\n72\n\n72\n\n11\n\n11\n\n6\n\n7\n\n\n\n38\n\nI (0+)\n\nModerate\n\n64\n\n62\n\n11\n\n10\n\n5\n\n6\n\n84\n\nII (1+)\n\nModerate\n\n73\n\n73\n\n12\n\n12\n\n6\n\n6\n\n\n\n39\n\nI (0+)\n\nModerate\n\n58\n\n58\n\n9\n\n9\n\n5\n\n5\n\n84\n\nII (1+)\n\nModerate\n\n69\n\n68\n\n11\n\n10\n\n6\n\n6\n\n\n\n39\n\nI (0+)\n\nModerate\n\n68\n\n68\n\n10\n\n9\n\n6\n\n6\n\n84\n\nII (1+)\n\nModerate\n\n67\n\n65\n\n11\n\n12\n\n6\n\n6\n\n\n\n39\n\nI (0+)\n\nModerate\n\n68\n\n68\n\n9\n\n9\n\n5\n\n6\n\n85\n\nII (1+)\n\nModerate\n\n71\n\n70\n\n11\n\n11\n\n6\n\n6\n\n\n\n39\n\nI (0+)\n\nModerate\n\n66\n\n64\n\n10\n\n10\n\n6\n\n6\n\n85\n\nII (1+)\n\nModerate\n\n74\n\n73\n\n11\n\n11\n\n6\n\n6\n\n\n\n39\n\nI (0+)\n\nModerate\n\n63\n\n65\n\n10\n\n10\n\n6\n\n6\n\n85\n\nII (1+)\n\nModerate\n\n64\n\n65\n\n10\n\n11\n\n6\n\n6\n\n\n\n39\n\nI (0+)\n\nModerate\n\n66\n\n68\n\n9\n\n9\n\n5\n\n5\n\n86\n\nII (1+)\n\nModerate\n\n76\n\n76\n\n11\n\n11\n\n7\n\n6\n\n\n\n40\n\nI (0+)\n\nModerate\n\n58\n\n57\n\n10\n\n10\n\n6\n\n6\n\n86\n\nII (1+)\n\nModerate\n\n71\n\n72\n\n11\n\n11\n\n6\n\n6\n\n\n\n40\n\nI (0+)\n\nModerate\n\n63\n\n63\n\n10\n\n11\n\n5\n\n6\n\n87\n\nII (1+)\n\nModerate\n\n70\n\n70\n\n12\n\n11\n\n6\n\n6\n\n\n\n40\n\nI (0+)\n\nModerate\n\n66\n\n68\n\n10\n\n10\n\n6\n\n6\n\n89\n\nII (1+)\n\nModerate\n\n63\n\n63\n\n10\n\n10\n\n5\n\n5\n\n\n\n40\n\nI (0+)\n\nModerate\n\n65\n\n67\n\n10\n\n11\n\n6\n\n6\n\n91\n\nII (1+)\n\nModerate\n\n68\n\n69\n\n11\n\n11\n\n6\n\n6\n\n\n\n41\n\nI (0+)\n\nModerate\n\n67\n\n66\n\n11\n\n11\n\n6\n\n6\n\n91\n\nII (1+)\n\nModerate\n\n58\n\n58\n\n11\n\n11\n\n5\n\n5\n\n\n\n41\n\nI (0+)\n\nModerate\n\n61\n\n60\n\n10\n\n10\n\n6\n\n6\n\n91\n\nII (1+)\n\nModerate\n\n69\n\n70\n\n10\n\n11\n\n6\n\n6\n\n\n\n41\n\nI (0+)\n\nModerate\n\n67\n\n66\n\n10\n\n11\n\n6\n\n6\n\n92\n\nII (1+)\n\nModerate\n\n72\n\n72\n\n11\n\n11\n\n6\n\n6\n\n\n\n43\n\nI (0+)\n\nFast\n\n67\n\n67\n\n10\n\n11\n\n6\n\n6\n\n95\n\nII (1+)\n\nFast\n\n70\n\n70\n\n10\n\n11\n\n6\n\n6\n\n\n\n43\n\nI (0+)\n\nFast\n\n68\n\n66\n\n11\n\n11\n\n6\n\n6\n\n95\n\nII (1+)\n\nFast\n\n65\n\n63\n\n11\n\n11\n\n6\n\n6\n\n\n\n43\n\nI (0+)\n\nFast\n\n64\n\n66\n\n11\n\n11\n\n6\n\n7\n\n98\n\nII (1+)\n\nFast\n\n63\n\n64\n\n12\n\n12\n\n5\n\n6\n\n\n\n43\n\nI (0+)\n\nFast\n\n68\n\n72\n\n10\n\n10\n\n6\n\n5\n\n99\n\nII (1+)\n\nFast\n\n72\n\n70\n\n11\n\n11\n\n5\n\n5\n\n\n\n44\n\nI (0+)\n\nFast\n\n65\n\n65\n\n10\n\n10\n\n6\n\n6\n\n99\n\nII (1+)\n\nFast\n\n69\n\n67\n\n11\n\n11\n\n6\n\n6\n\n\n\n44\n\nI (0+)\n\nFast\n\n68\n\n66\n\n10\n\n10\n\n6\n\n6\n\n100\n\nII (1+)\n\nFast\n\n74\n\n74\n\n12\n\n12\n\n6\n\n6\n\n\n\n44\n\nI (0+)\n\nFast\n\n66\n\n68\n\n10\n\n11\n\n6\n\n6\n\n101\n\nII (1+)\n\nFast\n\n62\n\n61\n\n12\n\n11\n\n6\n\n6\n\n\n\n45\n\nI (0+)\n\nFast\n\n66\n\n66\n\n11\n\n11\n\n6\n\n6\n\n102\n\nII (1+)\n\nFast\n\n58\n\n59\n\n12\n\n11\n\n5\n\n6\n\n\n\n45\n\nI (0+)\n\nFast\n\n69\n\n70\n\n10\n\n11\n\n5\n\n6\n\n104\n\nII (1+)\n\nFast\n\n67\n\n67\n\n12\n\n12\n\n5\n\n5\n\n\n\n45\n\nI (0+)\n\nFast\n\n70\n\n68\n\n11\n\n10\n\n6\n\n6\n\n105\n\nII (1+)\n\nFast\n\n72\n\n72\n\n10\n\n11\n\n6\n\n6\n\n\n\n45\n\nI (0+)\n\nFast\n\n68\n\n71\n\n11\n\n10\n\n6\n\n6\n\n105\n\nII (1+)\n\nFast\n\n68\n\n67\n\n11\n\n11\n\n6\n\n6\n\n\n\n46\n\nI (0+)\n\nFast\n\n69\n\n69\n\n11\n\n10\n\n6\n\n6\n\n105\n\nII (1+)\n\nFast\n\n72\n\n74\n\n11\n\n11\n\n6\n\n6\n\n\n\n46\n\nI (0+)\n\nFast\n\n68\n\n66\n\n11\n\n10\n\n6\n\n5\n\n107\n\nII (1+)\n\nFast\n\n67\n\n67\n\n11\n\n12\n\n6\n\n6\n\n\n\n47\n\nI (0+)\n\nFast\n\n65\n\n65\n\n9\n\n9\n\n5\n\n5\n\n110\n\nII (1+)\n\nFast\n\n69\n\n69\n\n11\n\n10\n\n6\n\n6\n\n\n\n49\n\nI (0+)\n\nFast\n\n66\n\n67\n\n10\n\n11\n\n6\n\n6\n\n113\n\nII (1+)\n\nFast\n\n71\n\n71\n\n11\n\n10\n\n7\n\n6\n\n\n\n50\n\nI (0+)\n\nFast\n\n68\n\n66\n\n11\n\n10\n\n6\n\n6\n\n115\n\nII (1+)\n\nFast\n\n69\n\n70\n\n11\n\n11\n\n6\n\n6\n\n\n\n52\n\nI (0+)\n\nFast\n\n70\n\n71\n\n11\n\n10\n\n6\n\n7\n\n116\n\nII (1+)\n\nFast\n\n72\n\n72\n\n10\n\n11\n\n6\n\n6\n\n\nПриводити результати аналізу розподілів розглянутих тут ознак не будемо, оскільки в даному випадку маємо справу лише з частиною даних. Розподіл ознаки D_S на всій вивченій вибірці показано на рис. 11.1.2. Значних відхилень вправо або вліво (при порівнянні «правих» і «лівих» значень з використанням критерію Вілкоксона) також не виявлено.\nТе, які саме ознаки обчислювали при обробці даних, зрозуміло на підставі рис. 11.2.1. Позначення ознак на цьому рисунку відповідають тим, що використовувалися у статті Гелашвілі та співавторів.\n \nРис. 11.2.1. Заходи спрямованої та неспрямованої асиметрії за окремими ознаками, а також загальна міра флуктуючої асиметрії, використані при вивченні окунів\nТретій крок викладеної в попередньому пункті програми пов’язаний з вивченням розмірної залежності, тобто визначенням кореляції між абсолютним значенням асиметрії та розміром структур, асиметрія яких розглядається. У статті Гелашвілі та співавторів обговорюється необхідність нормування асиметрії, якщо зі зростанням особини зростає і абсолютне значення її асиметрії. У такому випадку збільшення абсолютної асиметрії може бути наслідком збільшення розмірів тіла.\nВипадок, при якому зі збільшенням розмірів тіла абсолютна асиметричність зменшується, у статті Гелашвілі та співавторів не розглядається. Як ні дивно, наш випадок саме такий (рис. 11.2.2).\n
\nРис. 11.2.1. Заходи спрямованої та неспрямованої асиметрії за окремими ознаками, а також загальна міра флуктуючої асиметрії, використані при вивченні окунів\nТретій крок викладеної в попередньому пункті програми пов’язаний з вивченням розмірної залежності, тобто визначенням кореляції між абсолютним значенням асиметрії та розміром структур, асиметрія яких розглядається. У статті Гелашвілі та співавторів обговорюється необхідність нормування асиметрії, якщо зі зростанням особини зростає і абсолютне значення її асиметрії. У такому випадку збільшення абсолютної асиметрії може бути наслідком збільшення розмірів тіла.\nВипадок, при якому зі збільшенням розмірів тіла абсолютна асиметричність зменшується, у статті Гелашвілі та співавторів не розглядається. Як ні дивно, наш випадок саме такий (рис. 11.2.2).\n \nРис. 11.2.2. Флуктуюча асиметрія демонструє значну від’ємну (!) кореляцію з довжиною тіла\nЯкби рівень ФА зростав із розміром тіла, його треба було б використовувати у нормованій формі (щоб усунути ефект збільшення показника асиметрії, просто пов’язаного зі збільшенням розмірів). Оскільки він знижується, ми, ймовірно, маємо справу з проявом якоїсь закономірності, яка потребує розуміння.\nПроведемо двофакторний дисперсійний аналіз, у якому як незалежні фактори розглянемо вік і швидкість росту, а в якості залежної величини – загальний показник флуктуючої асиметрії (суму трьох заходів асиметрії за окремими ознаками). Результат показано на рис. 11.2.3.\n
\nРис. 11.2.2. Флуктуюча асиметрія демонструє значну від’ємну (!) кореляцію з довжиною тіла\nЯкби рівень ФА зростав із розміром тіла, його треба було б використовувати у нормованій формі (щоб усунути ефект збільшення показника асиметрії, просто пов’язаного зі збільшенням розмірів). Оскільки він знижується, ми, ймовірно, маємо справу з проявом якоїсь закономірності, яка потребує розуміння.\nПроведемо двофакторний дисперсійний аналіз, у якому як незалежні фактори розглянемо вік і швидкість росту, а в якості залежної величини – загальний показник флуктуючої асиметрії (суму трьох заходів асиметрії за окремими ознаками). Результат показано на рис. 11.2.3.\n \nРис. 11.2.3. І вік, і група, виділена на підставі швидкості росту, значно пов’язані з флуктуючою асиметрією окунів\nХоча взаємодія факторів у двофакторному дисперсійному аналізі виявилася незначущою, ілюструючий його графік зручний, щоб показати характерні для кожної з груп значення ФА (рис. 11.2.4).\n
\nРис. 11.2.3. І вік, і група, виділена на підставі швидкості росту, значно пов’язані з флуктуючою асиметрією окунів\nХоча взаємодія факторів у двофакторному дисперсійному аналізі виявилася незначущою, ілюструючий його графік зручний, щоб показати характерні для кожної з груп значення ФА (рис. 11.2.4).\n \nРис. 11.2.4. Молоді особини асиметричніші за більш дорослих, а повільно‑ і швидко‑ростучі особини – асиметричніші за тих, хто демонструє середню швидкість росту. Чому?\nЙмовірно, пояснити показаний на рис. 11.2.4 результат можна за допомогою кількох гіпотез. Перерахуємо їх, а потім встановимо, яка з них найкраще підходить як пояснення. Розглянемо лише один аспект: значно (p=0,00132) меншу асиметрію окунів старшої вікової групи.\nГіпотеза 1. Умови змінилися. Попередній спостереженням рік був сприятливим, розвиток риб був стабільним. Рік спостережень виявився значно більш травматичним, і розвиток молоді риб виявився розрегульованим.\nГіпотеза 2. У міру росту і дорослішання риб їх асиметричність зменшується. Відносно симетричні особини старшої групи могли бути менш симетричними в молодшому віці.\nГіпотеза 3. Смерть менш симетричних особин імовірніше; в результаті селективної смертності частка симетричних особин у поколінні зростає.\nЩоб спростувати гіпотезу № 1, потрібно залучити дані інших років. Це зроблено (поза межами описуваного дослідження). І в випадку окунів, і в випадку деяких інших видів, за результатами досліджень різних років риби старших вікових виявляються більш симетричними.\nГіпотезу № 2 слід визнати вкрай малоймовірною. Відхилення від симетричного стану призводять до асиметричних навантажень на парні структури, які призводять до посилення асиметрії. Спостереження за асиметричними особинами в умовах зоокультури також не дозволяють вважати такий варіант ймовірним.\nГіпотеза № 3 відповідає ситуації стабілізуючого відбору. Те, що особини зі середньою (ймовірно, відповідною популяційній нормі) швидкістю росту виявляються більш симетричними, також добре узгоджується з цією гіпотезою.\nОтже, можна обґрунтовано припускати, що збільшення з віком частки симетричних особин у поколінні є результатом стабілізуючого відбору: більш імовірного виживання більш симетричних особин."}
\nРис. 11.2.4. Молоді особини асиметричніші за більш дорослих, а повільно‑ і швидко‑ростучі особини – асиметричніші за тих, хто демонструє середню швидкість росту. Чому?\nЙмовірно, пояснити показаний на рис. 11.2.4 результат можна за допомогою кількох гіпотез. Перерахуємо їх, а потім встановимо, яка з них найкраще підходить як пояснення. Розглянемо лише один аспект: значно (p=0,00132) меншу асиметрію окунів старшої вікової групи.\nГіпотеза 1. Умови змінилися. Попередній спостереженням рік був сприятливим, розвиток риб був стабільним. Рік спостережень виявився значно більш травматичним, і розвиток молоді риб виявився розрегульованим.\nГіпотеза 2. У міру росту і дорослішання риб їх асиметричність зменшується. Відносно симетричні особини старшої групи могли бути менш симетричними в молодшому віці.\nГіпотеза 3. Смерть менш симетричних особин імовірніше; в результаті селективної смертності частка симетричних особин у поколінні зростає.\nЩоб спростувати гіпотезу № 1, потрібно залучити дані інших років. Це зроблено (поза межами описуваного дослідження). І в випадку окунів, і в випадку деяких інших видів, за результатами досліджень різних років риби старших вікових виявляються більш симетричними.\nГіпотезу № 2 слід визнати вкрай малоймовірною. Відхилення від симетричного стану призводять до асиметричних навантажень на парні структури, які призводять до посилення асиметрії. Спостереження за асиметричними особинами в умовах зоокультури також не дозволяють вважати такий варіант ймовірним.\nГіпотеза № 3 відповідає ситуації стабілізуючого відбору. Те, що особини зі середньою (ймовірно, відповідною популяційній нормі) швидкістю росту виявляються більш симетричними, також добре узгоджується з цією гіпотезою.\nОтже, можна обґрунтовано припускати, що збільшення з віком частки симетричних особин у поколінні є результатом стабілізуючого відбору: більш імовірного виживання більш симетричних особин."}