
Экология: биология взаимодействия. 5.21. Клинальная изменчивость и некоторые экологические правила
Сравнивая друг с другом организмы, населяющие область с градиентным изменением какого-либо фактора, мы можем увидеть закономерное изменение тех или иных признаков этих организмов. Когда речь идет о внутривидовой изменчивости, такая постепенная географическая изменчивость называется клинальной. Пр...
Українська мова (найновіша версія) / російська версія (оновлення припинено)
5.20. Концепція ефективних температур
Д. Шабанов, М. Кравченко. Екологія: біологія взаємодії
Глава 5. Аутекологія та основи середовідознавства
5.22. Основні середовища проживання та їхні особливості
5.21. Клінальна змінність та деякі екологічні правила
Порівнюючи умови в різних частинах земної поверхні, ми можемо переконатися, що багато важливих екологічних факторів змінюються поступово, утворюючи градієнт — плавну послідовність змін). Наприклад, подорожуючи в якусь сторону, ми можемо спостерігати, як поступово зменшується висота над рівнем моря, зростає вологість і температура. На якусь межу відбудеться дискретна (різка) зміна умов середовища — наприклад, від суши ми перейдемо до моря. Але при продовженні нашого руху, уже по морю, ми знову побачимо, що багато факторів (глибина, освітленість) змінюються градієнтно.
Порівнюючи один з одним організми, що населяють область з градієнтною зміною якого-небудь фактора, ми можемо спостерігати закономірну зміну тих чи інших признаків цих організмів. Якщо йдеться про внутрішновидову змінність, така поступова географічна змінність називається клінальною. Говорячи про признаки, можна сказати, що вони утворюють кліни. Наприклад, і в широколиствених лісах, і в лісостепу, і навіть у степній та полупустельній зонах ми можемо знайти дубрави — дубові ліси. Перехід між названими зонами визначається в першу чергу кількістю доступної для рослин води. Якщо в зоні широколиствених лісів дуби ростуть на рівних поверхнях поза зв’язком з руслами річок, то в степу та полупустелі вони прив’язані до понижень (балок) і річкових долин — місцях із підвищеною вологістю. Порівнюючи дуби з цих дубрав один з одним, ми переконаємося, що в них змінюється багато признаків: зріст, діаметр стовбура, розмір листя та особливості будови листової пластинки. Ці відмінності пов’язані з реакцією рослин на поступово змінювані кліматичні признаки.
Дослідження клінальної змінності представляє особливий інтерес для вияснення особливостей зв’язку організмів із середовищем (рис. 5.21.1). Клінальна змінність є результатом відбору, що пристосовує кожну популяцію до локальних умов середовища, та взаємодії між сусідніми популяціями (спарювання, міграції), що згладжують відмінності між ними. Фактично клінальна змінність «проявляє», робить наочними та доступними для дослідження ці зазвичай приховані від дослідника процеси.
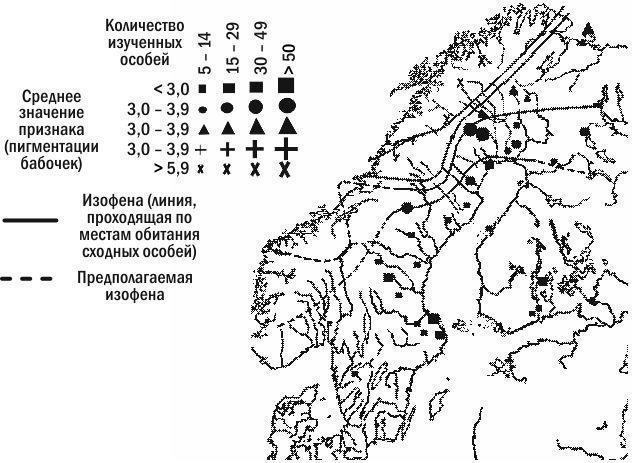
Рис. 5.21.1. На цій схемі показано клінальну змінність за пігментацією крил одного з видів метеликів, брюквенниць (Pieris napi), за результатами дослідження, виконаного на початку XX століття. Ізофени — лінії, що з’єднують області поширення організмів із однаковою вираженістю досліджуваного признака
Як правило, клінальна змінність організмів не вважається підставою для виділення підвидів. Географічні підвиди (форми всередині виду з самостійним статусом у системі організмів) виділяють за умови дискретної (перервистої, стрибкоподібної) змінності. Проте іноді між підвидами або навіть видами, здатними до гібридизації, існують перехідні зони — зони інтерградації.
У більшості випадків поняття кліна застосовують для опису внутрішновидової змінності, хоча подібні зміни можна зареєструвати, розглядаючи різноманіття різних видів одного роду або навіть сімейства, якщо вони ведуть подібний спосіб життя та населяють середовище з градієнтною зміною якого-небудь важливого для цих організмів фактора.
Деякі правила, що описують клінальну, а також міжвидову географічну змінність, сформульовано досить давно. Наприклад, ще в XIX столітті сформульовано правила Бергмана та Аллена, які стосуються як внутрішновидової змінності, так і відмінностей між близькими видами, що ведуть подібний спосіб життя.
Правило Бергмана (1847): серед споріднених форм гомеотермічних тварин, що ведуть подібний спосіб життя, ті, які живуть у холоднішому кліматі, мають більші розміри тіла.
Правило Аллена (1877): серед споріднених форм гомеотермічних тварин, що ведуть подібний спосіб життя, ті, які живуть у холоднішому кліматі, мають відносно менші виступні частини тіла: вуха, ноги, хвіст тощо.
Наприклад, географічна змінність звичайної лисиці відповідає обом названим правилам. Лінійні розміри тіла південних лисиць приблизно на 10–15 % менші, ніж у північних. Крім того, південні лисиці мають відносно довші вуха і хвіст. Якщо ми вийдемо за межі цього виду та розглянемо більш північних та більш південних родичів лисиці, ми побачимо, що вони підпорядковуються тій же закономірності. Полярні песці мають досить велике тіло, короткі лапи, морду, хвіст та вуха. Пустельні фенеки — це досить малі лисички з довгими лапами, мордою та хвіст і прямо-таки величезними вухами.
Найбільші тигри — амурські, що живуть у найсуворішому кліматі, а найменші — суматранські. У тундрі живуть вовки вагою до 50 кг, а в пустелях їхні розміри становлять лише 35 кг. Правило Бергмана застосовуємо й до міжвидової географічної змінності. Воно чудово підпорядковується ведмедям — від гігантських білих ведмедя та гризлі до малого гімалайського ведмедя. Найбільші пінгвіни (імператорський та королівський) живуть у Антарктиді, а найменші — на Галапагоських островах.
Правила Бергмана та Аллена мають подібну фізіологічну природу: вони відображають той факт, що в холодному кліматі простіше підтримувати постійну температуру тіла тим тваринам, які мають меншу відносну площу поверхні тіла. Тепловіддача через поверхню тіла пропорційна її площі, а теплоємність та теплопродукція — обсягу тіла. Зменшити відносну площу поверхні можна, збільшуючи лінійні розміри тіла, або «заокруглюючи» його, зменшуючи виступні його частини.
Як усі правила, правила Бергмана та Аллена мають багато винятків. Наприклад, їм не підпорядковуються копаючі ссавці. На їхні розміри та пропорції суттєво впливають особливості пересування в ґрунтовому середовищі, яке, зокрема, захищає їх від холодного повітря.
Наприклад, слони підпорядковуються правилу Аллена, але не підпорядковуються правилу Бергмана. Африканські слони більші за індійських, хоча живуть у більш спекотному кліматі. Це пов’язано з тим, що африканські слони переважно живуть на відкритих просторах (в савані), а індійські — пов’язані з лісами. До речі, лісові африканські слони менші, ніж індійські! Проте, як і слід було очікувати, африканські слони мають значні труднощі зі зниженням температури при перегріві, тому у них набагато більші вуха, ніж у індійських.
Чи можна порівнювати один з одним тварин, що ведуть несхожий спосіб життя? Чи можемо ми стверджувати, що правило Бергмана спростовують, наприклад, тропічні крилани (летючі лисиці, летючі собаки тощо), які набагато більші за летючих мишей помірних широт? Звісно, такий висновок був би помилковим. Ці тварини ведуть принципово різний спосіб життя. Крилани живляться переважно плодами, а наші летючі миші спеціалізовані на полюванні літаючих нічних комах. Різний характер харчування призводить до значних відмінностей у тепловому балансі цих тварин, що й обумовлює різницю в їхніх розмірах.
Чи застосовні правила Бергмана та Аллена до внутрішновидової змінності людини? Хоча всі люди й належать до одного виду, наша екологічна пластичність настільки велика, що в різних частинах ареалу люди ведуть різний спосіб життя. Ці відмінності перешкоджають прояву правила Бергмана. Разом із тим порівняння багатьох народів дозволяє побачити прояви правила Аллена. Так, ескімоси та інші корінні народи Крайнього Півночі мають крепких тіла з короткими руками, ногами та шиєю. Мешканці відкритих просторів екваторіальної Африки (наприклад, масаї) худі та мають відносно довгі й тонкі ноги й руки. Природно, прояви правила Аллена можна спостерігати лише на корінних народах, що існують у тісному взаємодії зі своїм середовищем проживання. Представники сучасного глобального людства, багато з яких переселялися по всьому світу та живуть у штучно зміненому середовищі, зазвичай не демонструють проявів названих екологічних правил.
Ще одним із відомих екологічних правил, що описує клінальну змінність, є правило Глогера, запропоноване в 1833 році орнітологом К. Глогером. Воно полягає в тому, що серед споріднених форм (підвидів або видів) гомеотермічних тварин ті, що живуть у умовах теплого й вологого клімату, мають яскравіший окрас, ніж ті, що живуть у умовах холодного й сухого клімату.
Ймовірними причинами, що призводять до такого характеру змінності, можуть бути міркування, пов’язані із захисною окраскою — ґрунти (або поверхня снігу) у холодному й сухому кліматі зазвичай світліші, ніж у теплому й вологому. Проте для пояснення правила Глогера цього обставини недостатньо, адже воно поширюється навіть на нічних тварин. Не виключено, що вологий і теплий клімат більш сприятливий для синтезу пігментів тварин. У певній мірі правило Глогера застосовуємо й до людини.
Додаткові матеріали:
Навчальна модель: Правило Бергмана
Навчальна модель: Правило Аллена
5.20. Концепція ефективних температур
Д. Шабанов, М. Кравченко. Екологія: біологія взаємодії
Глава 5. Аутекологія та основи середовідознавства
5.22. Основні середовища проживання та їхні особливості