Кравченко, Шабанов (2008) Возможные пути трансформации популяционных систем
Последняя версия "мандалы" (мандала - высокосимволический графический объект сложной структуры, используемый как тема для медитации) с возможными путями преобразования ГПС зеленых лягушек. Ее pdf-версия лежит тут. Кравченко М. А., Шабанов Д. А. Возможные пути трансформации популяционных...
Кравченко М. А., Шабанов Д. А. Можливі шляхи трансформації популяційних систем Pelophylax esculentus cоmpleх (Ranidae, Anura, Amphibia) // Праці Українського герпетологічного товариства. – № 1, 2008. – С. 15-20.
УДК 57.055:597.851
МОЖЛИВІ ШЛЯХИ ТРАНСФОРМАЦІЇ ПОПУЛЯЦІЙНИХ СИСТЕМ PELOPHYLAX ESCULENTUS COMPLEX (RANIDAE, ANURA, AMPHIBIA)
М. А. Кравченко1, Д. А. Шабанов2
Харківський національний університет ім. В.М. Каразіна,
пл. Свободи, 4, Харків, 61077 Україна;
E-mail: 1m_kravchenko@inbox.ru; 2d.a.shabanov@gmail.com
Можливі шляхи трансформації популяційних систем Pelophylax esculentus complex (Ranidae, Anura, Amphibia). Кравченко М. А., Шабанов Д. А.— Pelophylax esculentus комплекс включає два батьківських види: Pelophylax lessonae (Camerano, 1882) і Pelophylax ridibundus (Pallas, 1771)), а також їх гібрид — Pelophylax esculentus (Linnaeus, 1758). Ця група здатна формувати багатокомпонентні популяційні системи різного складу, які існують завдяки геміклональному успадкуванню, характерному для диплоїдних і триплоїдних гібридних жаб. Такі популяційні системи можуть змінюватися в часі. У роботі представлена схема можливих шляхів їх трансформації, яка може бути підставою для генеалогічної класифікації популяційних систем.
Ключові слова: Pelophylax esculentus (Rana esculenta), геміклональне успадкування, трансформації популяційних систем.
Possible Ways of Transformation of Population Systems of Pelophylax esculentus complex (Ranidae, Anura, Amphibia). Kravchenko M. A., Shabanov D. A.— Pelophylax esculentus complex includes two parental species: (Pelophylax lessonae (Camerano, 1882) and Pelophylax ridibundus (Pallas, 1771)) and their hybrid — Pelophylax esculentus (Linnaeus, 1758). This group has a peculiar ability to form multicomponent population systems of various composition, existing due to the hemiclonal inheritance typical of diploid and triploid hybrid frogs. These population systems can undergo transformation in time. We present a scheme of possible ways of their transformation, which might become a basis of their genealogical classification.
Key words: Pelophylax esculentus (Rana esculenta), hemiclonal inheritance, transformation of population systems.
До складу комплексу середньоєвропейських зелених жаб, Pelophylax esculentus complex, входять ставкова (Pelophylax lessonae (Camerano, 1882), стара назва— Rana lessonae) і озерна (Pelophylax ridibundus (Pallas, 1771) = Rana ridibunda) жаби, а також їх гібрид[1]— їстівна жаба (Pelophylax esculentus (Linnaeus, 1758) = Rana esculenta). Всі три форми здатні населяти одні місцеоселення і спільно розмножуватися, утворюючи змішані популяційні системи (далі — ПС). Однією з причин існування таких ПС є геміклональне (клональне для окремих геномів) успадкування у гібридних жаб. У ході гаметогенезу у гібридів деякі з батьківських геномів можуть елімінуватися, а інші (клональні) — переходити в гамети без рекомбінації.
ПС P. esculentus complex позначають заглавними буквами назв форм, які їх формують (Uzzel, Berger, 1975; Лада, 1995). Наприклад, виділяють L-, R-, E-, L-R-, L-E-, R-E-, і R-E-L-типи популяцій і ПС. Ця класифікація не вичерпує різноманіття відомих ПС зелених жаб. Так, до складу деяких таких систем можуть входити триплоїдні особини; у такому разі ПС із ди- і триплоїдних гібридів можна віднести до Et-типу. У деяких ПС гібридні жаби можуть бути представлені лише однією статтю. Так, R-Em-тип ПС відповідає змішаній системі з P. ridibundus і самців P. esculentus (m— male— самець), а R-Ef-тип — аналогічній системі, в якій P. esculentus представлені лише самками (f— female— самка) (Лада, 1995).
Особливістю P. esculentus complex є високе різноманіття їх ПС навіть у межах відносно обмежених територій. Наприклад, у Харківській обл. зареєстровані (А. В. Коршунов, усн. повідомл.) системи R-, L-, R-E-, R-Et-, R-Em- і R-E-L-типів (і залишається дискусійним існування систем E-типу). Слід зазначити, що переважна частина цього регіону знаходиться за межами ареалу P. lessonae, і для цієї області в цілому характерні системи R-E-типу. Несподіваною знахідкою стало виявлення в цьому регіоні значної частки триплоїдів серед гібридних жаб (Borkin et al., 2004).
Чудовою властивістю ПС P. esculentus complex є їх здатність до трансформації, яка відзначена в ряді робіт і випливає з теоретичних уявлень про природу таких систем (Holenweg Peter, Reyer, 2002; Vorburger, Reyer, 2003; Межжерін та ін., 2005). Одним із прикладів таких перетворень може бути доля ПС Іськового ставу в окол. біостанції Харківського університету, докладно розглянута в іншій публікації (Шабанов та ін., 2006). Це означає, що ПС зелених жаб слід розглядати не лише у статиці, але й у динаміці. Ми запропонували можливу схему перетворення таких ПС, що враховує специфіку передачі клональних геномів (Шабанов та ін., 2006; А. В. Коршунов, усн. повідомл.). У цій роботі ми представляємо розширений і вдосконалений варіант цієї схеми (рис. 1).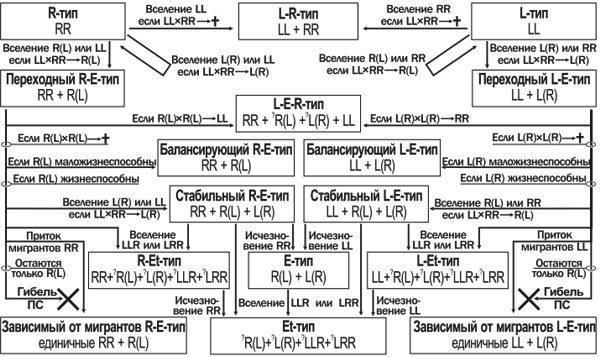
Рис. 1. Можливі шляхи трансформацій популяційних систем (ПС) Pelophylax esculentus complex.
Позначення: L— геном Pelophylax lessonae; R— геном Pelophylax ridibundus; L, R— неклональні (такі, що рекомбінуються) геноми; (L), (R) — клональні (такі, що не рекомбінуються) геноми
Основним методом цієї роботи є теоретичний аналіз літературних і оригінальних даних про різноманіття популяційних систем зелених жаб з урахуванням уявлень про закономірності геміклонального успадкування, характерного для гібридних особин.
Відправною точкою в перетворенні ПС можна вважати чисту популяцію батьківського виду. Розглядаючи можливість вселення до неї жаб з іншими генотипами, можна виділити три вірогідних шляхи перетворення цієї ПС. Якщо до вихідної популяції батьківського виду потраплять особини іншого батьківського виду, і утворення гібридів між ними з якихось причин виявиться неможливим, ці два види можуть залишитися в одному місцеоселенні в тому співвідношенні, яке визначатиметься їх конкурентоспроможністю в даних умовах. Повернення до стану одновидової популяції можливе при конкурентному витісненні одного виду іншим. Якщо ж до вихідної ПС потраплять гібридні особини або особини іншого батьківського виду, схрещування з якими призведе до утворення гібридів, то доля такої ПС визначиться тим, які геноми передаватимуться клонально. Якщо клональним виявиться консспецифічний батьківськийі геном, вже в наступному поколінні система повернеться до початкового стану. Якщо клональний геном виявиться гетероспецифічним, то ПС почне трансформуватися. Співвідношення особин батьківського виду і гібридів від покоління до покоління змінюватиметься. З трьох типів схрещування, можливих у такій системі, два (батьківський вид — батьківський вид і гібрид — гібрид) не змінюють співвідношення основних форм у ПС, а третє (батьківський вид — гібрид) призводить до зростання частки гібридів. При схрещуванні особин батьківського виду і гібридів з гетероспецифічними гаметами (RR× R(L) —>R(L), а також LL× L(R) —> L(R)) у потомстві присутні лише гібриди. Це призведе до зростання частки гібридів у складі розглядуваної ПС.
Те, чим завершаться такі зміни, залежить від кількох факторів. Першим із них є можливість виживання особин іншого батьківського виду, які виділяються при схрещуванні гібридів, т. зв. «гібридолізі» (Plotner, 2005): R(L) × R(L) —>- LL і L(R) × L(R) —>- RR. Якщо ці особини життєздатні, то система переходить до L-E-R-типу ПС. Однак нерідко особини, що несуть два однакових клональних геноми, виявляються нежиттєздатними. Цей ефект розглядають як наслідок так званого «тріщотки Мюллера» — накопичення несприятливих мутацій у геномах, які передаються без рекомбінації (Хедрік, 2003).
Якщо життєздатність гібридів і особин батьківського виду порівнянна, трансформація ПС R-E- або L-E-типу може призвести до повн