
Maro, Shabanov (2005) Механизмы расселения серой жабы и особенности ее популяций в недавно заселенных местообитаниях
Маро А. Н., Шабанов Д. А. Механизмы расселения серой жабы (Bufo bufo (L. 1758); Amphibia, Anura) и особенности ее популяций в недавно заселенных местообитаниях // Матеріали Першої конференції Українського Герпетологічного Товариства. – К.: Зоомузей ННПМ НАНУ, 2005. – С. 107–110.
Маро А. Н., Шабанов Д. А. Механізми розселення сірої жаби (Bufo bufo (L. 1758); Amphibia, Anura) і особливості її популяцій у нещодавно заселених місцевих середовищах // Матеріали Першої конференції Українського Герпетологічного Товариства. – К.: Зоомузей ННПМ НАНУ, 2005. – С. 107–110.
Механізми розселення сірої жаби (Bufo bufo (L., 1758); Amphibia, Anura) і особливості її популяцій у нещодавно заселених місцевих середовищах
А. Н. Маро, Д. А. Шабанов
Харківський національний університет імені В. Н. Каразина, пл. Свободи, 4, Харків 61077
This work is devoted to the studies of common toad Bufo bufo (Linnaeus, 1758) artificial moving to new habitats consequences that was performed in Kharkov region in 1994. The natural dispersion of toads has been found to occur in two ways. The first way includes a transfer of tadpoles with water. The second way includes incoming of toads into the reservoirs located between a former place of spawning and a ground habitat of toads settled by toads upon their metamorphosis. The new-founded groups of breeding (local populations) differ from their progenitor groups with the substantially smaller size of males and bigger size of females. It seems that on settling of new habitats, common toad males tend to accelerate their maturing, while females tend to accelerate their growth.
Для жаб роду Bufo характерно утворення добре ізольованих груп розмноження (локальних популяцій), пов’язаних з нерестовими водоймами і підтримуваних завдяки філопатії. Кожна група розмноження володіє унікальністю, обумовленою її історією, особливостями населених місцевих середовищ і взаємодією з сусідніми групами. Вивчення формування нових груп розмноження при розселенні жаб може бути дуже корисним для розуміння їх особливостей. У 1994 р. в Харківській обл. за участі одного з авторів даної роботи було виконано штучне розселення сірих жаб (Bufo bufo (Linnaeus, 1758)). У ході вивчення його наслідків нами вивчені вибірки сірих жаб з 8 перелічених нижче місцевих середовищ (всього 244 половозрілих особин, спійманих на нересті, з яких 84 самки і 160 самців) і проведені польові спостереження за ходом їх нересту.
1. Стариця: берег р. Північний Донець в окр. біостанції ХНУ імені В. Н. Каразина в с. Гайдари Зміївського р-ка; обмелевше старе русло річки у вологій дубраві з заростями черемші; 9 самок, 9 самців.
2. Коряков яр: окр. біостанції ХНУ; затоплений ставок у дубраві; 22 самки, 27 самців.
3. Іськов ставок: окр. біостанції ХНУ; балковий ставок між дубравою і лугом; 4 самки, 13 самців.
4. Нижній ставок, П’ятихатки: окр. с. Лісне Харківського р-ка; нижній з трьох ставків в Очешетянській балці в дубраві; 15 самок, 19 самців.
5. Верхній ставок, П’ятихатки: там же, верхній з трьох ставків; 8 самок, 16 самців.
6. Верхній ставок, Ольхова балка: окр. с. Русська Лозова Харківського р-ка, верхній з 7 ставків у широкій балці, що проходить через нагірну дубраву; 9 самок, 26 самців.
7. Нижній ставок, Ольхова балка: 6‑й ставок у виході балці в пойму р. Харків; 7 самок, 22 самця.
8. Добра балка; ставок у балці, що виходить у пойму поруч з Ольховою балкою; 10 самок, 28 самців.
Екологічно місцеві середовища 2—8 дуже схожі і типові для сірої жаби в Харківській обл.; місце середовища 1 суттєво відрізняється від них. Географічно вивчені точки утворюють три групи, відстань всередині яких не перевищує 5 км: 1—3; 4 і 5; 6—8. Групи розмноження в точках 1—3 є старими, а 4—8 — молодими; наймолодші локальні популяції в точках 5 і 8. Генеалогічний зв’язок вивчених популяцій: 3->4->5; 3->6->7->8. Розселення 3->4 і 3->6 здійснені штучно в 1994 р. в результаті переміщення не відкладали ікру жаб, спійманих на нересті в Іськовому ставку. У П’ятихатках перенесено близько 200 особин обох статей, в Ольхову балку — близько 100 особин; при цьому була вилучена лише мала частина процвітальної групи розмноження Іськового ставка. Для вселення жаб були обрані водойми, екологічно схожі на Іськовий ставок, але розташовані в добре вивченому харківськими герпетологами лісовому масиві, де відсутні сіри жаби. Подальше поширення жаб проходило природно, причому у випадку 4->5 проти течії води в системі ставків, у випадку 6->7 — за течією води, а у випадку 7->8 — через водоразділ. У 1995 і 1996 рр. під час нересту сірих жаб знайти їх у нижньому ставку в П’ятихатках не вдалося. Ймовірно, вселені жаби віднерестилися в новому для них водоймі, але вдруге в неї не повернулися. Розвиток групи розмноження виявився пов’язаний з поверненням потомства особин‑засновників у водойму, де відбувалося їхнє розвиток.
В Ольховій балці, куди жаби були перенесені у верхній ставок, вони швидко розповсюдилися по всій системі, пов’язаній спільним водотоком. Очевидно, їх розселення було пов’язане з перенесенням головастиків водою. У П’ятихатках середній (розташований між нижнім і верхнім) ставок заселявався суттєво пізніше нижнього, а пізніше за всіх виявився заселений верхній ставок (5‑те місце середовища). У 2004 р. на верхньому ставку в П’ятихатках зареєстровано лише кілька половозрілих жаб, що прийшли на нерест, а в 2005 р. їх кількість збільшилася більш ніж на порядок. Очевидно, що в даному випадку був використаний інший механізм розселення, спрямований проти водотоку. Для його розуміння суттєві спостереження, зроблені в 2005 р. на середньому ставку.
Ми спостерігали, що більшість пар жаб у амплексусі на середньому ставку рухалися в напрямку нижнього ставка. Самки підходили до ставка, несучи на собі самців, або зустрічали їх у воді. Потім пари рухалися в напрямку утримуваної ставком плотини, виходили на берег і скочували в нижній ставок. Лише деякі пари на середньому ставку залишалися в ньому і далі не переміщувалися. Ймовірно, при розселенні личинок з нижнього ставка деякі з них мігрували за середній ставок. По досягненню половозрілості, рухаючись до місця свого розвитку, вони спускалися в балку і потрапляли в розташований на їхньому шляху середній ставок. Більшість жаб подолало його і дійшло до кінцевої точки свого маршруту, але деякі не встигали дістатися туди і нерестилися в середньому ставку. Їх потомство поверталося вже в середній ставок і з часом колонізувало верхній.
Отже, можна припустити, що освоєння нових місць нересту жабами відбувається двома способами: при перенесенні головастиків течією води і при попаданні у водойми, розташовані між раніше освоєним місцем нересту і наземним місцем існування розселившихся особин.
Порівняння вибірок проведено за 24 морфометричними, 21 якісним і 8 дискретним ознаками жаб, а також за 29 пропорціями (відношеннями морфометричних ознак). Компонентний аналіз проводився за нормалізованими середніми значеннями у вибірках і виконувался за допомогою пакету Statistica for Windows (рис. 1). Видно, що самки виявилися менш змінливими, ніж самці, і взаємне розташування їх вибірок на площині головних компонент більш відповідає генеалогічним відносинам між групами розмноження. Відмінності вибірок з окр. біостанції ХНУ (1-3) перевищують відмінності вибірок з молодих груп розмноження, які в більшості випадків виявляються найближчими до вибірки з батьківського місцевого середовища (3). Самці з наймолодших груп розмноження (5 і 8) найсильніше відрізняються від інших за своїми абсолютними розмірами, і дуже близькі до представників батьківської групи розмноження за якісними, дискретними ознаками і пропорціями тіла.
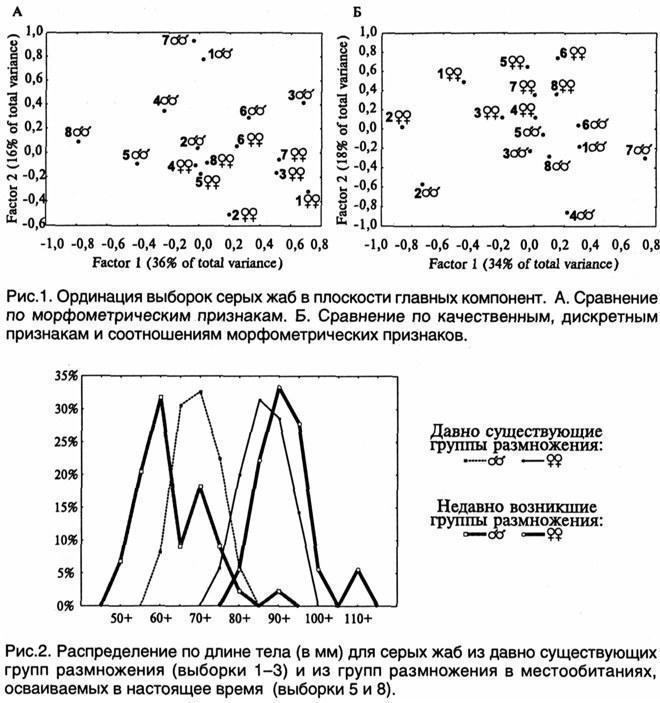
Молоді групи розмноження високодостовірно відрізняються за розмірами тіла складових їх особин (рис. 2). У заселяних місцевих середовищах у нересті беруть участь (і входять до складу утворюваних пар) дуже малі самці (розмір тіла 45—60 мм). Можливо, це молоді особини, що не беруть участі у нересті стабільних популяцій. Другий піковий вияв самців у «молодих» групах розмноження (65—70 мм) відповідає їх характерному розміру у давно заселених місцевих середовищах. Парадоксальним чином участь у нересті самців‑«недорослих» поєднується з більшими розмірами самок. Оскільки у заселяючому верхньому ставку в П’ятихатках (вибірка 5) на нерест одночасно прийшли і малі самці, і великі самки, вони схожі за своїм віком. Ймовірно, ці самки характеризувалися особливо швидким ростом. Таким чином, можна зробити висновок, що при заселенні нових місцевих середовищ у самців сірих жаб відбувається прискорення дозрівання, а у самок — росту. Виявлений феномен потребує подальшого вивчення (зокрема, визначення віку і швидкості росту жаб).
Автори висловлюють подяку А. В. Коршунову, М. А. Кравченку і Т. С. Фоменко за допомогу у зборі матеріалу і польових спостереженнях.