Парадокс ускорения эволюции. Колонка в КомпьютерреOnline #46
Могли ли в изменчивой среде возникнуть организмы, у которых эффективность выработки новых приспособлений снизилась бы, – они ведь неминуемо проиграли бы тем, кто эволюционировал быстрее?
←
Dmytro Shabanov
→
Труднощі профорієнтації
Парадокс прискорення еволюції
Коли відбір стає неефективним?
Колонка в Комп'ютерреOnline #45
Колонка в Комп'ютерреOnline #46
Колонка в Комп'ютерреOnline #47
У позапрошлий раз я обґрунтовував, що в русі до III синтезу еволюційна біологія повинна подолати обмеження II синтезу — СТЕ, синтетичної теорії еволюції, що сформувалася близько сімдесяти років тому. Пора поговорити про недоліки СТЕ.
Для нас поки буде достатньо визначення СТЕ, яке дав Олександр Павлович Расницин. Згідно з цією теорією,
"еволюційний процес представляється як динаміка алельних частот у популяціях, контрольована відбором (через оцінку життєздатності та, відповідно, диференціальне відтворення фенотипів) і стохастичними процесами".
Не зовсім ясно? Я поясню, старанно виклавши класичні для СТЕ погляди (навіть ті, з якими не повністю згоден). Це простіше зробити, коментуючи рисунок.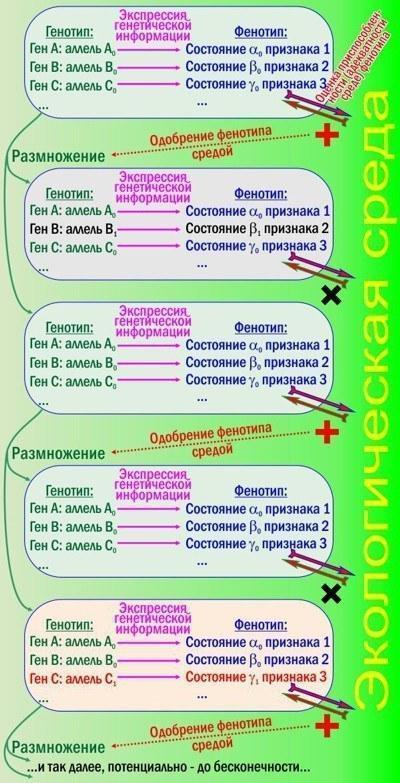
Рис. 1. Дарвінівська еволюція згідно СТЕ. Відбирається фенотип, відтворюється генотип
П'ять прямокутників на рисунку — три покоління дуже простих організмів: бактерій, наприклад. Термінологія зрозуміла? Комплекс успадкованих задатків організму називається генотипом, а сукупність його проявлених ознак — фенотипом. Генотип — сукупність генів; гени можуть перебувати в різних станах — алелях. У фенотипі можна виділити окремі ознаки, які можуть перебувати в різних станах ознаки.
У ході розвитку організму його фенотип змінюється, причому його зміни залежать від генотипу. Організм взаємодіє зі середовищем, будуючи себе за її рахунок. Якщо фенотип адекватний тим можливостям, які надає середовище, організм успішно розвивається і залишає потомство; неадекватні (непристосовані) фенотипи знищуються або відсторонюються від розмноження.
Бачите на рисунку петлі зворотного зв'язку? Генотип визначає фенотип, середовище оцінює фенотип і дозволяє розмножуватись носіям оптимальних генотипів. При розмноженні виникають помилки генотипу. Відбір на рисунку відсів зміни гена В і підтримав зміни гена С. Якщо так, ми спостерігаємо елементарну еволюційну подію: зміну алельних частот у популяції.
Ви пам'ятаєте, що нова інформація — це завжди наслідок запам'ятовування наслідків випадкового вибору? Дарвінівська еволюція — неминучий наслідок розмноження, змінливості, успадкованості та відбору. Ця тетрада на рис. 1 присутня — отже, еволюція буде йти. Звісно, успіх і невдача особин не завжди закономірні; відбір діє через зміну їхніх ймовірностей; тому в визначенні Расницина згадані стохастичні процеси.
Загальна картина ясна, проблема — у деталях. Щоб відбір фенотипів ефективно змінював генотипи, треба, щоб фенотипи і генотипи були однозначно пов'язані один з одним. Так буває не завжди.
На жаль, до сьогодні не розроблена загальноприйнята класифікація ознак (і їх можливих станів) залежно від того, як керується їх розвиток. Показані на рис. 1 ознаки керуються найпростішим способом: кожна з них залежить від певного гена. Так, можливо, що ген В визначає структуру ферменту, що дозволяє бактеріям живитися якимось речовиною. Коли ген нормальний (аллель В0), все добре. У другому поколінні мутація В1 порушила роботу ферменту, порушила засвоєння їжі і призвела організм до загибелі.
Мутація С1 цікавіша: вона призвела не до зниження, а до підвищення життєздатності. Можливо, вона посилила зв'язки між сусідніми ланцюжками амінокислотних залишків у якомусь білку, через що цей білок став стійкішим до підвищених температур.
Чи рівноймовірні несприятливі та корисні мутації? Чим складніший і досконаліший організм, тим менш ймовірно, що випадкова зміна принесе йому користь. Деякі автори навіть вважають ймовірність корисної зміни астрономічно малою. Мабуть, це не так, і переконливі експерименти (присвячені саме пристосуванню бактерій до підвищеної температури) показують, що корисні мутації — явище нечасте, але спостережуване.
Але чи будь-яка ознака залежить від одного гена? Ні. Наприклад, у щойно згаданому експерименті з еволюцією термостійкості бактерій ефект багатьох мутацій неможливо було оцінити окремо, без урахування алельного стану інших генів. Одна і та сама мутація в комплекті з одними алелями інших генів виявляється сприятливою, у поєднанні з іншими — шкідливою. Це означає, що визначальний стан ознаки залежить від кількох генів.
Чи знижують взаємодії між генами ефективність відбору? Так, і особливо для видів з статевим розмноженням. Я наведу приклад саме для таких організмів, кожен з яких має по два однакових або різних алеля кожного гена і передає нащадкам лише один з них.
Уявімо, що певний щасливий організм набуває поєднання двох рідкісних (можливо, нещодавно виниклих) алелей, кожен з яких окремо несприятливий: А1 і В1. У популяції поширені алелі А0 і В0. Поєднання А1 і В1 сприятливі, а ось А1 і В0, як і А0 і В1 — шкідливі (і те, і інше — у порівнянні з нормою, А0 і В0).
Не буду пояснювати логіку розв'язання генетичних задач, а просто розпишу співвідношення чисельностей груп нащадків від схрещування щасливого власника вдалого поєднання двох мутацій з найймовірнішим партнером. Розглянемо випадки, коли обидва ці алелі домінантні (впливають на фенотип навіть у особин, у яких присутні в одній копії) і коли вони рецесивні (проявляються у фенотипі лише у організму, що має дві їх копії).
Якщо алелі А1 і В1 домінантні, лише чверть нащадків матиме покращений фенотип (показані напівжирним шрифтом), а дві чверті — погіршений (виділені підкресленням):
А1А0В1В0 × А0А0В0В0 → А1А0В1В0 : А1А0В0В0 : А0А0В1В0 : А0А0В0В0.
Якщо розглянуті алелі рецесивні, у всіх нащадків буде нормальний фенотип:
А1А1В1В1 × А0А0В0В0 → А1А0В1В0.
А що проявиться у наступному поколінні?
А1А0В1В0 × А1А0В1В0 → А1А1В1В1 : 2А0А1В1В1 : 2А1А1В1В0 : 4А1А0В1В0 : 2А1А0В0В0 : 2А0А0В1В0 : А0А0В1В1 : А1А1В0В0 : А0А0В0В0.
Сприятлива ознака проявиться лише у одного нащадка з шістнадцяти, а шість з шістнадцяти нащадків матимуть погіршений фенотип! У цих (і інших, не розглянутих нами) випадках подібної взаємодії алелей вони частіше будуть зустрічатися в популяції окремо, коли відбір працюватиме проти них, ніж разом, у стані, схваленому відбором. Ситуації, коли відбір підтримуватиме взаємодоповнюючу пару алелей, виявляться рідкісними порівняно з випадками, коли він знищуватиме ці алелі поодинці.
Взаємодія генів знижує ефективність відбору! І чим складніші організми, чим складніше визначаються їхні ознаки, тим в меншій мірі спрямований відбір фенотипів буде приводити до спрямованого відбору алелей.
У по-справжньому складних організмів між генотипом і фенотипом вкладаються ще кілька механізмів регуляції, що ускладнюють сортування алелей відбором. Один із них у термінах СТЕ називається розширенням норми реакції. Організми з однаковим генотипом можуть відрізнятися через розвиток у різних умовах (або просто внаслідок випадковості). Уявіть собі трьох людських близнюків‑третят з однаковими генотипами. Одного виховало плем'я первісних мисливців, інший потрапив у інтернат для спортсменів‑силовиків, третій — у спеціальну школу. Незважаючи на ідентичну успадкованість, вони виростуть дуже різними — і фізично, і за поведінкою. На відміну від ненаправлених мутацій, їхні відмінності підвищать пристосованість їх носіїв. При відборі (наприклад, статевому — завойовуванні серця потенційно плодовитої красуні) ці три близнюки матимуть різні шанси, адже відбір (і статевий, зокрема) оцінює саме фенотипи!
Другий механізм пов'язаний з епігенетичною регуляцією генної активності, особливо характерною для ссавців. Залежно від стану організму, він передає нащадкам геноми з різними хімічними "мітками" на генах (метилюванням або іншими змінами "букв" генетичного тексту). Ці мітки впливають на активність генів і, отже, на ознаки. Зазвичай епігенетичні мітки забезпечують той варіант розвитку (в межах норми реакції), який більш відповідає даним умовам середовища. При змінених умовах ці мітки можуть бути стерті або переставлені.
Як ці зміни впливають на ефективність відбору? Нову схему з багатьма поколіннями малювати не буду, покажу лише логіку формування ознак однієї особини.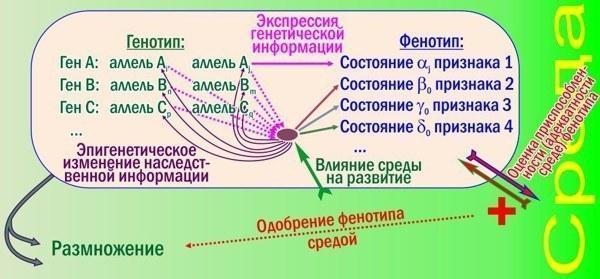
Рис. 2. Тут показані лише деякі ускладнювальні роботу відбору при сортуванні алелей механізми, характерні для високорозвинених видів
Окрім основної петлі зворотного зв'язку (генотип впливає на фенотип; відбираючи кращі фенотипи, середовище відбирає кращі генотипи) з'явилися нові рівні регуляції. Дивіться: розвиток ознаки 1 і надалі керується досить просто, він залежить від одного алеля. А ось ознаки 2, 3 і 4 залежать і від усіх показаних генів, і від середовища, причому в різній мірі. Взаємодія генних продуктів і середовищних впливів формує складний вузол взаємопереплетених причин і наслідків. Від результатів такої взаємодії залежить і сама генна активність: у залежності від неї гени піддаються епігенетичному редагуванню — оборотному успадкованому зміненню.
Ці зміни знижують ефективність сортування алелей відбором. Зв'язок фенотипів з генотипами стає дуже неоднозначним (крім "простих" ознак, як ознака 1 на рис. 2). Механізм еволюції, ефективно працювавший на рис. 1, буде гальмуватись на рис. 2.
Еволюція — це змагання у швидкості вироблення нових пристосувань. Поверніться до першого рисунка: у другому показаному там поколінні виграв нормальний генотип, а в третьому носій недавньої норми вже програв, оскільки з'явилися більш пристосовані організми. Як же могли виникнути організми, у яких ефективність відбору (і ефективність вироблення нових пристосувань) знизилася, — вони ж неминуче мали програвати тим, хто еволюціонував швидше?
Справа в тому, що у складних організмів ефективність відбору не знижується, а зростає. Знову процитую Расницина:
...вік (у мільйонах років) півчасової фауни (у якій половина видів — сучасні, половина — вимерлі) у великих ссавців (хоботних і копитних) — 0,2, у малих — 0,5, у птахів і риб — 0,7, у комах — 3‑7, у молюсків — 3,5‑5, у діатомових водоростей — 15. Час піввимирання (за аналогією з радіоактивним розпадом, час вимирання половини початкової кількості видів) у тих же одиницях найменший у слонів — 0,18, а в середньому у ссавців — 0,54, у кісткових риб — 3,5, у граптолитів — 1,3, у голкових — 4,2, у двостворок — 7, у планктонних формініфер — 5, у Benthos — 18‑24, у діатомових водоростей — 5,5, у дінофлагеллат — 9. На вищих таксономічних рівнях різниці подібні, але часто різкіші. Наприклад, вік півчасової фауни для родів ссавців — 4 млн років, для птахів — 10, рептилій — 20, риб — 30‑50, комах — 40, молюсків — 60, формініфер — 230"
Окрім викладених причин, існує ще цілий ряд наочних факторів, які мали б зробити еволюцію великих тварин дуже повільною (про них докладно пише Расницин). Наприклад, у кожної особини хоботних є лише небагато потомків, а тривалість поколінь катастрофічно велика. За той час, поки росте один слоненя, зміняться десятки (якщо не сотні) поколінь комах, діатомей або формініфер з безліччю особин. У хоботних просто не буде достатньо матеріалу для відбору, вони мають еволюціонувати надзвичайно повільно...
Рис. 3. За кілька десятків мільйонів років історії хоботних виникло понад сорок їх родів і понад триста видів!
За швидкістю їх еволюції (виміряної і у швидкості утворення нових видів, і у швидкості зміни ознак) хоботні — одні з рекордсменів у біосфері. З точки зору СТЕ цей феномен необґрунтований. Потрібна ідея еволюції еволюції — удосконалення у міру розвитку живих самих механізмів вироблення пристосувань. Наведу лише один приклад. Сімейство Люди різко підвищило швидкість вироблення пристосувань, коли удосконалило механізми культурної успадкованості і додало її до загальної, генетичної успадкованості. Ймовірно, це удосконалення еволюційного механізму не було першим у нашій історії.
Слони, як і інші ссавці, еволюціонують і з використанням механізмів, описаних СТЕ, і якимось іншим шляхом. Додаткові петлі зворотного зв'язку у реалізації їх генетичної інформації не знижують, а підвищують ефективність їх еволюції. Як — ми ще не знаємо до кінця. Що ж, нашому знанню є куди еволюціонувати!
←
Dmytro Shabanov
→
Труднощі профорієнтації
Парадокс прискорення еволюції
Коли відбір стає неефективним?
Колонка в Комп'ютерреOnline #45
Колонка в Комп'ютерреOnline #46
Колонка в Комп'ютерреOnline #47