Экология: биология взаимодействия. 3.10. Природа и характеристики сообществ
Естественно, что самой яркой и интересной частью экосистем являются сообщества — совокупности их живых компонентов. Являются ли сообщества четко отличающимися отдельными объектами? Изредка — да, чаще — нет. Спор не закончен до сих пор. Сообщества можно характеризовать по ряду пр...
Українська мова (найновіша версія) / Російська мова (оновлення припинено)
3.09. Тенденції в сукцесіях
Д. Шабанов, М. Кравченко. Екологія: біологія взаємодії
Розділ 3. Біогеоценологія та екологія угруповань
3.11. Трофічні зв'язки та рівні
3.10. Природа та характеристики угруповань
Звісно, що найяскравішою та найцікавішою частиною екосистем є угруповання — сукупності їхніх живих компонентів. Взаємодії між особинами різних видів та популяціями відбуваються саме всередині угруповань. Деякі з цих зв'язків утворюють досить хитромудрі ланцюжки. Наведемо тут лише один приклад, який давно став класичним.
«Дарвін виявив, що джмелі з їхнім довгим хоботком — єдині комахи, які здатні запилювати глибокі трубчасті квітки червоної конюшини. З цього він зробив висновок, що поширення червоної конюшини в Англії пояснюється великою кількістю джмелів. При цьому, посилаючись на одну з ентомологічних робіт, він вказує, що найчастіше гнізда джмелів зустрічаються поблизу міст і сіл, де їх менше нищать полівки, що поїдають личинок і лялечок. Чому ж в околицях міст і сіл мало полівки? Та тому, що там багато кішок, які сильно знижують чисельність популяцій полівки. Один німецький вчений продовжив це міркування наступним чином: якщо доведено, говорив він, що кішки відповідальні за поширеність в Англії конюшини — основного корму великої рогатої худоби, а конюшина екологічно пов'язана з британським морським флотом, оскільки яловичина — основна їжа моряків, то, отже, кішкам належить головна заслуга в тому, що Британія є великою морською державою. Наступний крок зробив Томас Гекслі: він стверджував — частково жартома, — що оскільки кішок в Англії тримають переважно старі діви, то британську могутність можна логічно — і екологічно — вивести з «кошаколюбства» численних англійських стареньких» (П. Фарб, 1971).
У результаті спільної еволюції різні частини угруповання пристосовуються одне до одного. У результаті цього багато типів угруповань мають характерний видовий склад і цілком певне співвідношення організмів, що належать до різних екологічних груп.
Чи є угруповання чітко відмінними окремими об’єктами? Зрідка — так, частіше — ні. На початку XX століття розгорілася дискусія про природу угруповань між американськими екологами. Ф. Клементс та інші прихильники організмічної концепції екосистем розглядали угруповання як надорганізми. Навпаки, Х. Глізон та інші прихильники індивідуалістичної концепції розглядали угруповання як конгломерати видів зі схожими вимогами до середовища.
Суперечка не закінчена досі. З одного боку, градієнтний аналіз (тобто вивчення поширення видів уздовж градієнтів зміни якихось значущих, насамперед кліматичних, факторів) показав, що межі поширення окремих видів не обов'язково збігаються з межами угруповань (рис. 3.10.1 і 3.10.2). Чим крутіші градієнти умов, тим чіткіші межі поширення видів. Межі поширення видів-домінантів характерніші за межі розподілу випадкових видів. З іншого боку — оскільки угруповання мають здатність до саморегуляції, їхні межі розташовані досить складно і не цілком надійно можуть бути визначені за допомогою градієнтного аналізу.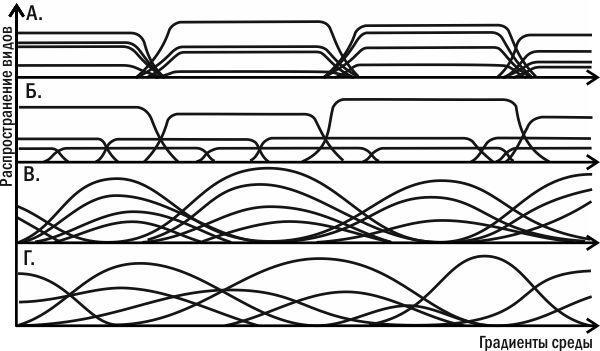
Рис. 3.10.1. Теоретичні криві, що ілюструють можливі варіанти зміни поширення видів уздовж градієнта (плавного зміни) певного істотного чинника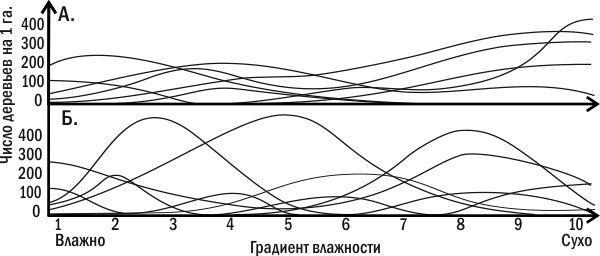
Рис. 3.10.2. Фактичні криві поширення різних видів дерев залежно від градієнта вологості. Зареєстровані криві не відповідають жодному з теоретичних варіантів, показаних на рис. 3.10.1
«На гніздовій території пари великих строкатих дятлів (Dendrocopos major L.) у приміському дубовому лісі ростуть сотні стовбурів дерев кількох видів, тисячі, якщо не десятки тисяч стебел трав'янистих рослин десятків видів (строго кажучи, на цій території можна виділити одну або кілька рослинних асоціацій); кожен стовбур є місцем проживання для десятків і сотень особин кількох видів комах-ксилофагів та тисяч особин комах-фітофагів, що харчуються зеленою масою, квітками та плодами; трав'янисті рослини формують місця проживання для десятків тисяч комах-фітофагів та тисяч особин їхніх хижаків; у лісовій підстилці та верхніх горизонтах ґрунту мешкають тисячі й десятки тисяч особин десятків видів комах (хижаків, сапрофагів, копрофагів), багатоніжок, павукоподібних, сухопутних ракоподібних, черв’яків, а чисельність сотень видів мікроорганізмів (бактерії, гриби, найпростіші), що мешкають у ґрунті, обчислюється десятками, якщо не сотнями мільйонів. Крім того, окрім пари дятлів, на цій території можуть гніздитися одна-дві пари повзиків (Sitta europaea L.), по кілька пар великих синиць (Parus major L.), мухоловок-білошийок (Ficedula albicolllis (Temm.)), зябликів (Erithacus rubecula L.), зябликів (Fringilla coelebs L.),піснярів-тріскотників (Phylloscopus sibilatrix Viell.), а для такого хижака, як яструб-тетерев'ятник (Accipiter gentilis L.), ця територія може становити лише невелику частину (від 1/25 до 1/10) його гніздової та мисливської території. Тут же розміщені території десятка особин землерий (Soricidae) і мишоподібних гризунів (Muridae), може прогодуватися одна білка (Sciuris vulgaris L.), а для горностая (Mustella erminea L.) або ласки (M. nivalis L.) так само, як і для яструба, ця територія становить приблизно 1/10–1/5 їхньої території пошуку їжі, не кажучи вже про великих копитних ссавців (лось (Alces alces L.), козуля (Capreolus capreolus L.), кабан (Sus scrofa L.)), для кожної особини яких необхідна територія в сотні разів більша і які можуть також бути виявлені на даній території, якщо площа лісового масиву достатньо велика або він пов'язаний із сусідніми масивами» (Ю.І. Вергелес, 2000).
Ймовірно, угруповання все-таки є реальною екологічною одиницею. Функціонування угруповання складається з діяльності особин, що його утворюють, але являє собою щось більше, ніж їхню суму. Ефективність угруповання та його стабільність зростають пропорційно тому, наскільки злагоджені, еволюційно «притерті» популяції, що його утворюють. Приклад значення «притертості» видів у спільноті один до одного — реакції спільнот на інтродуцентів (чужинців). Найчастіше інтродуценти не можуть увійти до складу цілісних спільнот і вимирають, але іноді дають вибухи чисельності, змінюючи функціонування спільнот.
Спільноти можуть бути повночленними (що включають продуцентів, консументів і редуцентів) і неповночленними.
Спільноти можна характеризувати за рядом ознак, розглядаючи їх емергентні властивості, відсутні на рівні частин цілісної системи. Деякі з них такі.
Склад угруповання (видова структура) — види, з представників яких складається угруповання. Найчастіше вигляд угруповання визначається якимось основним (або кількома основними) видами. Такі види називаються едіфікаторами. Зазвичай навколо окремих особин видів-едіфікаторів розвивається комплекс із тісно пов’язаних з ним видів — консорціум. Консорції є структурними одиницями угруповань.
Чисельність — кількість особин на одиницю площі або об’єму.
Частота видів — частка особин певного виду до загальної чисельності особин.
Різноманіття — видове багатство. Воно тим вищим, чим більша кількість видів, і тим нижчим, чим більше особин кожного виду трапляється в окремій вибірці.
Просторова структура — особливості розташування особин одна відносно одної. Розрізняють вертикальну структуру, або ярусність, і горизонтальну — мозаїчність. Ярусність характерна для фітоценозів, що складаються з рослин, які відрізняються за висотою. Приклад ярусності в лісі: I — дерева першої величини (ялина, сосна, дуб, береза, осика); II — дерева другої величини (горобина, черемха); III — підлісок із чагарників (ліщина, бересклет, черемха); IV — підлісок із високих чагарничків і великих трав (багульник, лохина, верес, аконіт, іван-чай); V — низькі чагарнички та дрібні трави (водяник, журавлина, кислиця); VI — мохи, надґрунтові лишайники, печіночники.
Екологічна структура — співвідношення основних екологічних груп організмів, а також різних життєвих форм.
Періодичність — добова, сезонна, багаторічна, столітня.
3.09. Тенденції в сукцесіях
Д. Шабанов, М. Кравченко. Екологія: біологія взаємодії
Розділ 3. Біогеоценологія та екологія угруповань
3.11. Трофічні зв’язки та рівні