
Расницын, 2008. Теоретические основы эволюционной биологии - 01
Здесь представлена первая часть монографии: В.В. Жерихин, А.Г. Пономаренко, А.П. Расницын. Введение в палеоэнтомологию. М.: Товарищество научных изданий КМК. 2008. 371 с. Текст выложен с согласия автора и предназначен, в первую очередь, для использования студентами в учебной работе. 1.1. ПРОЦЕСС ...
A.P.Rasnytsyn. Теоретичні основи еволюційної біології // V.V.Zherykhyn, A.H.Ponomarenko, A.P.Rasnytsyn. Вступ до палеоентомології. М.: КМК. 2008. 371 с
1.1. ПРОЦЕС ЕВОЛЮЦІЇ
1.1.1. СИНТЕТИЧНА ТЕОРІЯ ЕВОЛЮЦІЇ
1.1.2. ЕПІГЕНЕТИЧНА ТЕОРІЯ ЕВОЛЮЦІЇ
1.1.2.1. Основні положення
1.1.2.2. Адаптивна компроміс
1.1.2.3. Проблеми
1.2. МЕТОДОЛОГІЯ ФІЛОГЕНЕТИКИ, ТАКСОНОМІЇ ТА НОМЕНКЛАТУРИ
1.2.1. ФІЛОГЕНЕТИКА
1.2.1.1. Аналіз груп
1.2.1.2. Аналіз ознак
1.2.1.2.1. Аналіз різниць
1.2.1.2.2. Аналіз схожості
1.2.1.3. Комп’ютерний кладизм
1. ТЕОРЕТИЧНІ ОСНОВИ ЕВОЛЮЦІЙНОЇ БІОЛОГІЇ
Еволюція і систематика комах у цій книзі (і в наступних випусках цієї серії) реконструюється, виходячи з припущень, іноді не надто звичних для вітчизняних біологів, вихованих на синтетичній теорії еволюції. Тому здається корисним дати тут коротке обґрунтування моїх поглядів на еволюційний процес і на способи вивчення його результатів. Детальніше ця точка зору викладена в інших роботах (Rasnytsyn, 1987, 2002, 2005; Rasnitsyn, 1996, 2006).
1.1. ПРОЦЕС ЕВОЛЮЦІЇ
Уявлення про повну і остаточну перемогу дарвінізму в сучасній біології не зовсім відповідають дійсності: альтернативні погляди зберігаються і продовжують розвиватися. Серед інших свідчень на це вказує, наприклад, появлення об’ємної книги Ю.В. Чайковського (2003) з багатообіцяючою назвою «Еволюція». Тому, перш ніж розбирати конкуруючі парадигми селектогенезу (того напрямку думки, який частіше називають невдалим терміном дарвінізм, див. нижче), мені доведеться якимось чином обґрунтувати вибір саме цього напрямку.
Як альтернатива селектогенезу пропонуються різні концепції. Чайковський (2003) називає чотири системи поглядів на еволюційний процес: дарвінізм, номогенез, ламаркізм і жоффруїзм (за іменем одного з відомих ранніх еволюціоністів Е. Жоффруа Сент-Ілера). Ця класифікація невдала і термінологічно, і за суттю. Ім’ям засновника логічно називати вчення, суть яких полягає в тлумаченні канонічних текстів (християнство, буддизм, марксизм), а не системи наукових поглядів, оскільки логіка наукового пошуку майже неминуче веде до перегляду більшої чи меншої частини тверджень попередніх авторів. У результаті іноді виникають анекдотичні ситуації. Той же Ю.В. Чайковський при всьому своєму яскравому антидарвінізмі визнає не лише відбір (у сильно скороченій формі) і успадкування набутих властивостей (у сильно розширеній формі), а й пангенезис, і виявляється більшим дарвіністом, ніж ті, хто вважаються і вважають себе послідовниками Дарвіна. Тому поза цим розділом я буду по можливості уникати використання таких назв.
По суті запропонована класифікація невдала в тому сенсі, що її підрозділи перекриваються настільки широко, що їх не розрізнити. Для номогенезу зазначено особливу увагу до закономірного характеру еволюційного процесу, але, як відомо, еволюційні дослідження всіх напрямків зайняті пошуком (не без успіху!) таких закономірностей (см., наприклад, Раутіан, 1988). Навіть якщо звузити номогенез до визнання широкої повторюваності еволюційних шляхів (паралелізми, рефрени Мейєна тощо), то, як буде видно далі, цей підхід прекрасно сумісний з селектогенезом, що визнає важливу роль відбору.
Жоффруїзм визначений як напрямок, що надає особливе значення онтогенетичним перетворенням як механізму еволюції: у рівній мірі це справедливо для епігенетичної версії селектогенезу (Воддінгтон – Шмальгаузен – Шишкін – Rasnytsyn – Tikhomirova, см. нижче). Обмежена ефективність відбору, визнання якої Чайковський вказує як важливу рису номогенезу, жоффруїзму і ламаркізму, також належить до найважливіших положень епігенетичного селектогенезу (см. нижче).
З ламаркізмом ситуація трохи складніша. З цим терміном пов’язують два суттєво незалежних твердження — про прогресивний характер еволюційного процесу (ускладнення і вдосконалення організмів, їх будови і функціонування) і про успадкування набутих властивостей. Проблема прогресу природно входить у проблематику селектогенезу (знову посилаюся на свою роботу: Rasnytsyn, 1971). Успадкування набутих властивостей довгий час вважалося несумісним з дарвінізмом (хоча сам Дарвін вважав інакше), але коли з відкриттям ретровірусів була доведена зворотна зв’язок фенотипу з генотипом, минаючи відбір, виявилося, що нічого серйозного для селектогенезу не сталося. Дійсно, якщо організму вигідно, скажімо, закріпити в ДНК певну модифікацію і якщо можливо розробити механізм такої фіксації (наприклад, за участю ретровірусів), чому відбір не може реалізувати такий механізм? Інша справа, що ДНК існує саме для надійного зберігання інформації і, відповідно, для мінімізації зовнішніх впливів на її структуру, тому будь‑який механізм, цілеспрямовано здійснює таке вплив, небезпечний для виконання основної функції ДНК. Саме з цим, на мій погляд, пов’язаний той факт, що успадкування набутих властивостей не належить до числа рутинних механізмів еволюційного процесу.
Отже, ламаркізм, жоффруїзм і номогенез у наведеній вище трактовці не становлять принципової альтернативи селектогенезу. Водночас усім зрозуміло, що принаймні ламаркізм і номогенез — це щось зовсім інше, ніж сучасний дарвінізм (тобто селектогенез), просто їх фундаментальна (суттєва) відмінність ще не сформульована. Що стосується жоффруїзму, його описано недостатньо, щоб розташувати його на якусь сторону водоразділу, про який йтиме мова.
На мою думку, суттєва різниця між селектогенезом з одного боку, ламаркізмом і номогенезом з іншого, полягає у ставленні до джерела доцільності. Доцільною (адаптивною) називають структуру (включно з системою дій), яка виглядає, принаймні зовні, як створена спеціально для виконання певної, у певному контексті корисної функції. Тому пояснення походження доцільності передбачає урахування майбутньої функції виникаючої структури (системи): в певному сенсі майбутня функція формує структуру.
Походження суттєво нової доцільної структури (системи) можливо лише шляхом прямого цілепокладання, тобто за участю розуму (грубо кажучи, людини або бога), або шляхом запам’ятовування і відтворення вдалого вибору, тобто через відбір. Ефективність відбору як методу пошуку доцільного рішення продемонстровано У. Ешбі (1964) на моделі гомеостата. Для застосування до еволюційного процесу ця модель була модифікована як паралельний гомеостат, що відрізняється тим, що ступеневі змінні використовуються не послідовно, як у Ешбі, а одночасно: у їх ролі виступають фенетично і генетично різні особини (Rasnytsyn, 1971). Іншим яскравим прикладом паралельного гомеостата є, за сучасними уявленнями, система імунітету теплокровних, як її описав Чайковський (2003) на с. 189‑190.
Безсумнівним механізмом прийняття доцільних рішень є розум. Також безсумнівно, що розум людського типу виникає пізно і не може бути джерелом еволюційної доцільності. Оскільки суттєва доцільність еволюції, здається, ніким не заперечується, мова може йти про розум, або про іманентне життю, або про передіснуюче йому, тобто про надприродну (грубо кажучи, божественну) природу еволюційної доцільності. Саме тут, на мій погляд, лежить водоразділ між селекціонізмом, що відмовляється розглядати еволюцію як продукт розумової діяльності, і номогенезом і ламаркізмом, що передбачають надприродне (альтернативне відбору) джерело прогресу (ламаркізм) або доцільності (номогенез). Оскільки залучення надприродних факторів еволюції, на мій погляд, законно лише в межах віри, а з наукою несумісне, вибір селектогенезу для мене неминучий.
У сучасній теорії селектогенезу конкурують два головних підходи — синтетичний і епігенетичний. Синтетичну теорію еволюції, більш знайому нашим біологам, можна охарактеризувати як редукціоністську, що зводить ефекти на рівні цілого до явищ на рівні його елементів — передусім елементарних генетичних факторів і їх частот у популяціях. Протиставну їй епігенетичну теорію можна лише дуже умовно назвати антиредукціоністською: її прихильники намагаються зрозуміти властивості еволюційного процесу, виходячи з деяких властивостей цілого організму, передусім — його онтогенезу. По суті це подальший розвиток холістичного підходу до еволюції (Smuts, 1926, cit. per Smuts, 1987). Взаємовідносини синтетичної та епігенетичної гіпотез спеціально розглянуті М.А. Шишкиним (1987, 1988а, 2006) і Д.Л. Гродницьким (2000, 2002). Спираючись на їхні результати, спробую викласти своє бачення цієї проблеми.
1.1.1. СИНТЕТИЧНА ТЕОРІЯ ЕВОЛЮЦІЇ
Синтетична, а точніше — популяційно‑генетична теорія еволюції (бо синтез там скоріше проголошений, ніж досягнутий) розглядає успадкуваність, тобто здатність до стабільного відтворення фенотипу і його ознак у послідовних поколіннях, як властивість особливих елементарних носіїв‑генів, нині ототожнюваних з ділянками (локусами) хромосом і далі з ділянками ДНК (цистронами тощо). Відповідно еволюційний процес представлений як динаміка частот алелей у популяціях, контрольована відбором (диференціальним відтворенням фенотипів). Ознаки фенотипу виникають і змінюються внаслідок мутацій і рекомбінацій генів. Оскільки всі ці зміни не мають властивості первісної доцільності, саме відбір формує склад популяцій і властивості організмів, і робить це з точністю, обмеженою лише стохастичними (ймовірнісними) факторами. Окрім фактора випадковості, єдиною перешкодою повному відповідності організації живих істот вимогам відбору є обмін генами, який уніфікує структуру популяцій на рівні, що відповідає не локальним, а усередненим характеристикам відбору.
Відповідно до цих поглядів організація живих істот представлена м’якою глиною в руках відбору, набором ознак, вільно тасованих відбором. Тому еволюційний процес з цієї точки зору виявляється рівномірним — принаймні настільки, наскільки рівномірні варіації середовища і відбору. Рівномірність еволюції стабільно порушується лише дивергенціями і вимираннями, які в розглянутій системі поглядів і становлять єдину природну основу для побудови системи організмів.
Синтетична теорія, звичайно, визнає і плейотропність генів (вплив одного гена на кілька ознак), і полигенність ознак (множинну генетичну обумовленість окремої ознаки), так що кожна ознака фактично залежить від усього геному. Однак ці визнання в реальності декларативні, оскільки тотальна взаємозалежність елементів структури і функцій організму і, відповідно, відсутність будь‑якого постійного внеску гена у пристосованість роблять неможливим зведення еволюційного процесу до елементарних генетичних, а тим більше молекулярно‑генетичних подій.
Ці спрощення не безневинні: вони ведуть до невідповідностей між передбаченнями теорії і результатами спостережень. Наведемо деякі з таких невідповідностей. Перш за все звернемося до темпів еволюційного процесу, оцінених за швидкістю виникнення і вимирання таксонів того чи іншого рангу. Таксон — єдине поняття, сенсом існування якого є спроба інтегрованої оцінки схожості і різниці організмів. Однак на практиці порівнянність таксонів одного рангу в різних групах не забезпечена нічим, окрім інтуїції систематиків (про що йдеться нижче), тому такий метод оцінки швидкості еволюції недосконалий. Але інших способів порівняння темпів еволюції організмів у нас просто немає. Яка б не була недосконалість (суб’єктивність) оцінки різниць у таксономічних рангах, вона більш інформативна, ніж більш об’єктивна оцінка різниць за допомогою ознак. Добре відомо, що зовні однакові різниці можуть досягатися настільки різними шляхами і виявляються проявами настільки різномасштабних перетворень організації, що оцінка темпів еволюції за їх допомогою лише вводитиме в оману. Особливо яскраво цей висновок підтверджують матеріали щодо еволюції в островних умовах і на ранніх етапах еволюції таксонів (феномен архаічного різноманіття). Тут швидко і легко виникають гротескні форми з незбалансованою організацією. Вони часто відрізняються за такими ознаками, які в нормі (у стабілізованих групах) характеризують таксони значно вищого рангу (детальніше см. нижче). Водночас палеонтологічна оцінка темпів таксономічної еволюції дає результати і виявляє тенденції, які навряд чи можна списати на несумісність таксонів одного рангу в різних групах (Rasnytsyn, 1987).
Згідно з синтетичною теорією еволюційний процес керується в основному тими ж факторами і механізмами, якими оперує генетика популяцій. Тоді і швидкість макроеволюції має підкорятися тим же закономірностям, що і швидкість зміни частот генів у генетиці популяцій. Ця швидкість, у свою чергу, визначається потужністю потоку адаптивно компетентних генетичних варіацій (змін, значимих для відбору), які протікають через еволюціонує суму. Отже, швидкість має бути вищою, чим вища частота мутацій і швидкість зміни поколінь, чим більший розмір популяцій і мобілізаційний резерв змінності.
Палеонтологічний матеріал не дозволяє прямо оцінити більшість перелічених параметрів, що використовуються в генетиці популяцій. Деякі з них все ж можуть бути косвенно оцінені у викопних — у тій мірі, в якій вони корелюють з іншими ознаками, такими як розміри, таксономічна приналежність тощо. Зокрема, якщо генетичний підхід до еволюції справедливий, то її швидкість має бути мінімальною у ссавців, що мають порівняно повільну зміну поколінь і невеликі популяції (через відносно великі розміри особини), максимальною у одноклітинних і проміжною у безхребетних. На практиці ж, як показують численні палеонтологічні дані, ці співвідношення мають зворотний характер (Rasnytsyn, 1987). При цьому різниці дуже великі навіть на рівні виду, а ця таксономічна категорія вважається порівнянною в різних групах. Так, вік півжиттєвої фауни (фауни, у якій половина видів сучасна, половина — вимерлі) у великих ссавців (слонових і копитних) 0,2 млн років, у малих 0,5, у птахів і риб 0,7, у комах 3‑7, у молюсків 3,5‑5, у діатомових водоростей — 15 млн років. Час піввимирання (за аналогією з піврозпадом, час вимирання половини початкової кількості видів) найменший у слонів (0,18 млн років), в середньому у ссавців 0,54, у кісткових риб 3,5, у граптолитів 1,3, у голчатих 4,2, у двостулкових молюсків 7, у планктонних формініфер 5, у Benthos 18‑24, у діатомових водоростей 5,5, у дінофлагеллат 9 млн років. На вищих таксономічних рівнях різниці подібні, але часто різкіші. Наприклад, вік півжиттєвої фауни для родів ссавців 4 млн років, птахів 10, рептилій 20, риб 30‑50, комах 40, молюсків 60, формініфер 230 млн років.
Наведені цифри не ставлять під сумнів генетичні передбачення. Відбір абсолютно автоматичний: якщо в популяції з’являються особини, що різняться за ступенем пристосованості, вони будуть послідовно відбиратися у повній відповідності з положеннями генетики популяцій. У тій мірі, в якій ці різниці успадковані, різна участь особин у відтворенні популяції так само автоматично призведе до зміни її успадкованої структури. Отже, передбачуваний генетичними теоріями розподіл швидкостей еволюції обов’язково має мати місце в природі — у тій мірі, в якій потужність загального потоку генетичних варіацій у довгостроковій перспективі корелює з потужністю потоку відбираємих (підвищено адаптивних) успадкованих змін. Підтвердженням слугують добре відомі різниці за типічними швидкостями селекції між породами домашніх тварин, сортами рослин і штамами мікроорганізмів. Не менш показові і іноді спостережувані максимальні темпи еволюції комах, коли на острові Гаваї віком 0,5 млн років (Rotondo et al., 1981) в результаті 17 колонізацій виникло 46 видів жука роду Plagithmysus (Gressit, 1978), тобто не менше одного акту видоутворення відбувалося в середньому за 150‑200 тис. років. Г.Х. Шапошников майже вивів новий вид тлі за один сезон (см. нижче).
Водночас очевидно, що в цілому розподіл швидкостей еволюції на рівні виду і вище в першому наближенні протилежний передбаченому популяційною генетикою. Це означає, що популяційно‑генетичні обмеження не є такими для еволюційного процесу. Грубо кажучи, потік загальної генетичної змінності еволюційно надмірний навіть у популяціях великих ссавців, не кажучи вже про інші, більш чисельні та швидше розмножувані організми. Швидкість змін на надпопуляційному рівні (швидкість еволюції) має регулюватися якимось іншим фактором. Питання, які це можуть бути фактори, розглянемо пізніше. Тут можна лише підвести підсумок, що той рівень, до якого синтетична теорія еволюції намагається звести еволюційний процес і тим самим його пояснити, не відповідає покладеним на нього надіям і не дає очікуваних пояснень — принаймні щодо парадоксу еволюційних швидкостей.
Існують і інші групи даних, що не вкладаються в рамки синтетичної теорії еволюції. Якщо організація живих істот — м’яка глина в руках відбору або набір ознак, вільно тасованих відбором, то її результати мають бути передбачуваними в тій мірі, в якій відомі умови, у яких протікала еволюція. Антиселекціоністи продемонстрували на численних прикладах, що це не так (см., наприклад, Любищев, 1982). Навіть близькородинні форми в схожих, здавалося б, умовах можуть вести себе зовсім по‑різному.{"translated_text":"Найпарадоксальнішим прикладом цього є надзвичайно екстравагантна у репродуктивній сфері плямиста гієна та цілком звичайна у цьому відношенні близька до неї смугаста гієна.\nЩе менш пояснювана з синтетичної точки зору широко поширена, якщо не універсальна дискретність біологічного різноманіття. У цій системі поглядів природна лише одна форма дискретності — дискретність амфіміктичного виду за умов, коли обмін генами достатньо інтенсивний для уніфікації складу популяцій всупереч локальним варіаціям відбору. Але тоді між ступенем (повнотою та давністю) ізоляції і ступенем дивергенції має існувати хороша кореляція. Це в реальності не спостерігається: гальмуючий (вирівнювальний різниці) вплив обміну генами на швидкість дивергенції підтвердити не вдається. До такого висновку прийшов, зокрема, S.S. Shvarts (1980), який порівнював види ссавців з різною схильністю утворювати ізоляти. Очевидний контрприклад островного видоутворення насправді належить до іншого контексту (див. нижче).\nПовністю необґрунтована з точки зору синтетичної теорії еволюції дискретність видів у партеногенетичних і безстатевих організмів, позбавлених обміну генами. Порівняння близьких двостатевих і партеногенетичних видів у коловраток (Mair, 1974) і жук-стрічкових (Ivanova, 1978; V.V. Zherikhin, особисте повідомлення), двостатевих і безстатевих видів у протистів (Yu. Polyansky, 1957; Romashko, 1977), нижчих водоростей (V. Polyansky, 1956) і папоротей (Fagge, 1990) показало, що одностатеві і безстатеві види настільки ж дискретні, як і двостатеві.\nНе менш показовий тривалий (мільйони, десятки і, можливо, навіть сотні мільйонів років) еволюційний стазис на видовому і родовому рівнях, коли про обмін спадковою інформацією не може бути і мови. Так, популяції чотирьох або п’яти з тридцяти австралійських видів безслідних комах (Rgotuga) на видовому рівні не відрізняються від популяцій з далеких континентів і островів — Калімантан, Японія, Південна Африка, Європа (Tihep, 1967). Безслідні не стійкі до висихання і не залишають ґрунт, тому трансконтинентальні міграції для них практично виключені. Пояснюючи їх поширення, важко уникнути посилань на дрейф континентів, тим більше що для безслідних відомий і класичний «дрейфовий» ареал (Delamarentulus tristani Silv. розповсюджений на обох берегах Атлантичного океану, у Коста-Ріці та у Західній Африці; Tihep, 1963). Але з цього автоматично випливає вік виду, оцінюваний десятками мільйонів років.\nВідомі й більш прямі вказівки на велику давність деяких видів. Так, у фауні еоценового балтійського янтаря (вік не менше 40 млн. років) зараз відомо кілька видів комах і близько десятка видів кліщів, на видовому рівні не відрізняних від сучасних; ще частіше спостерігається персистенція протягом 10‑20 млн. років (Zherikhin, 1999). Знахідки різноманітних пліоценових галлів, не відрізняних від галлів, викликаних сучасними видами комах на тих самих рослинах, значно молодші (3‑5 млн. років), але вони не менш важливі (Zherikhin, 2002a). Морфологічна специфіка галла обумовлена біохімічним впливом комах‑галлоутворювача на тканини рослини‑хазяїна. Тому морфологічна стабільність галлів доводить біохімічний стазис комах протягом мільйонів років. Багато з цих знахідок походять із Північної Європи, отже мова йде про види, що пережили льодовиковий період (правда, переважно за рахунок міграцій), але не змінилися навіть біохімічно. Таким чином, стазис тривалістю в мільйони років здійснюється не лише за відсутності обміну генами, а й всупереч беззаперечним глибоким, можливо навіть катастрофічним, змінам умов.\nРекордсменом, однак, є щитень Triops cancriformis (Schaffer), який практично не змінився з раннього триасу (близько 230 млн. років), і навіть питання про видову самостійність пермських популяцій залишається невирішеним (Tasch, 1969). Це підтверджують дані молекулярних годинників для двох морфологічно майже нерозрізнених японських популяцій іншого виду, T. longicaudatus (LeConte): вони свідчать про розходження видів близько 15 млн. років тому. Для інших видів цього роду, включаючи T. cancriformis, дивергенції датуються 25‑45 млн. років (Suno‑Uchi et al., 1997).\nНе менш зрозумілий у цій системі поглядів звичайний феномен, коли старші таксони виявляються більш дискретними і демонструють більш ясні родинні зв’язки, ніж види. Дійсно, види розходилися відносно нещодавно, мали менше часу на дивергенцію, ніж старші таксони і, відповідно, мали зберегти більше слідів того, як саме протікала дивергенція (детальніше см. нижче).\nВсе сказане, на мій погляд, достатньо переконливо показує серйозну невідповідність між популяційно‑генетичним підходом з його похідними (синтетичною теорією еволюції та біологічною концепцією виду) і результатами спостережень. Тому настав час перейти до іншої системи поглядів на еволюційний процес.\n1.1.2. ЕПІГЕНЕТИЧНА ТЕОРІЯ ЕВОЛЮЦІЇ\n1.1.2.1. Основні положення\nНа відміну від синтетичної, епігенетична теорія бачить еволюційний процес передусім як процес еволюційного перетворення онтогенезу (Shishkin, 1987, 1988a, b, 2006; Rautian, 1993). При цьому особливу увагу приділяють цілісності онтогенезу в тому сенсі, що і сам процес, і його результат (будова організму на послідовних етапах його розвитку) значно стійкіші, ніж будь‑які окремі фактори та процеси розвитку. Нормальний розвиток еквіфінально і здатний релаксувати (пригнічувати, поглинати, вирівнювати) дуже широкий спектр впливів і збурень, як зовнішніх, так і внутрішніх, включаючи результати різних помилок і порушень нормального розвитку.\nТаким чином, успадковується (стійко відтворюється у наступних поколіннях) нормальний онтогенез (норма реакції) в цілому, а не окремі ознаки. Окрім норми існує маса різноманітних відхилень (аберацій) розвитку з нестійким відтворенням, які в нормальних умовах реалізуються рідко. У несприятливих умовах, коли механізми захисту норми вдається порушити або подолати — внаслідок сильного зовнішнього впливу чи внутрішніх причин (помилки розвитку), — розвиток іде по аберативному (відхильному і нестійкому) шляху. Оскільки ендогенні аберації розвитку легко виникають не лише при статевому, а й при партеногенетичному та безстатевому розмноженні, їх виникнення є реалізацією прихованої гетерогенності популяції, а не виникненням de novo якихось генетичних змін.\nЕпігенетична теорія розглядає аберації як матеріал відбору, який може створити на його основі нову адаптивну норму. Такий процес легше описати в рамках метафори епігенетичного ландшафту. Нормальний онтогенез являє собою стійку (стабілізовану) послідовність тісно пов’язаних окремих епігенетичних процесів. Більш‑менш окремі відрізки цієї послідовності називаються креодами. Якщо уявити креод у вигляді глибокої долини в певному ландшафті, по схилу якої тече онтогенетичний процес (рис. 1), то абераціями будуть пологі бокові долинки, підняті в бортах головної. Щоб аберація реалізувалася, тобто щоб онтогенез міг вийти в бокову долину, потрібен або сильний зовнішній вплив на розвиваючийся організм, що виштовхує його в бокову долину (рис. 2a), або зміна самого ландшафту, що виполює дно долини (додаткове аберативне зміна, рис. 2b), або те й інше разом (рис. 2c).\n
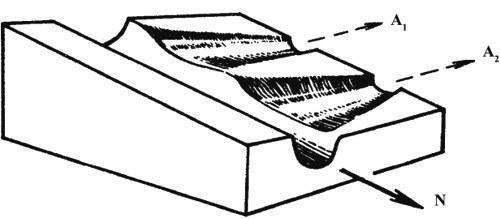
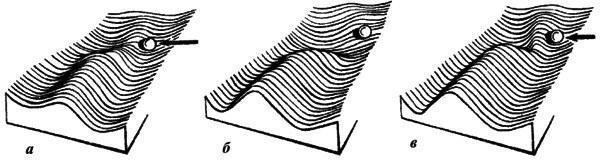
Оскільки аналіз окремих факторів не дає відповіді на питання про конкретний механізм, що забезпечує еволюційний процес, спробуємо підійти з іншого боку. Проаналізуємо ті умови, у яких еволюція реально йде, причому йде відносно швидко.
Хоча репродуктивна ізоляція, як уже говорилося, у нормальних умовах не є важливим еволюційним фактором, добре відомо, що у умовах ізольованих островів, водойм тощо еволюція йде особливо швидко і часто призводить до утворення сильно змінених, навіть гротескних форм. С. С. Шварц (1980), спеціально аналізувавший цей парадокс, дійшов висновку, що причина полягає у зниженій напруженості конкурентних відносин у бідних, незаповнених (у еволюційному масштабі часу) островних біоценозах. Конкретно еволюційна роль незаповненості цено́зів, за Шварцем, полягає в тому, що ослаблена конкуренція робить можливою швидку односторонню спеціалізацію. Це приваблива інтерпретація для концепції адаптивного компромісу: предковий вид, потрапляючи з материка в обіднений островний біоценоз, опиняється в умовах, пом’якшених за багатьма параметрами і допускаючих додаткову оптимізацію функцій, що залишилися під жорстким контролем середовища, за рахунок інших, неконтрольованих відбором так само суворо. Насправді, як ми побачимо, справа складніша і не зводиться до ослаблення відбору, хоча поки скористаємося цим нестрогим терміном.
Подана модель еволюції на островах цікава ще й тим, що вона відповідає концепції інадаптивної еволюції за В. О. Ковальовським (Расніцин, 1986). Більш важливо, однак, що вона демонструє досить реальний механізм, здатний подолати, зламати стійкість добре збалансованого адаптивного компромісу, що гальмує еволюцію. Очевидно виникає питання, чи може подібний механізм діяти не лише при заселенні островів, а й в якихось інших ситуаціях.
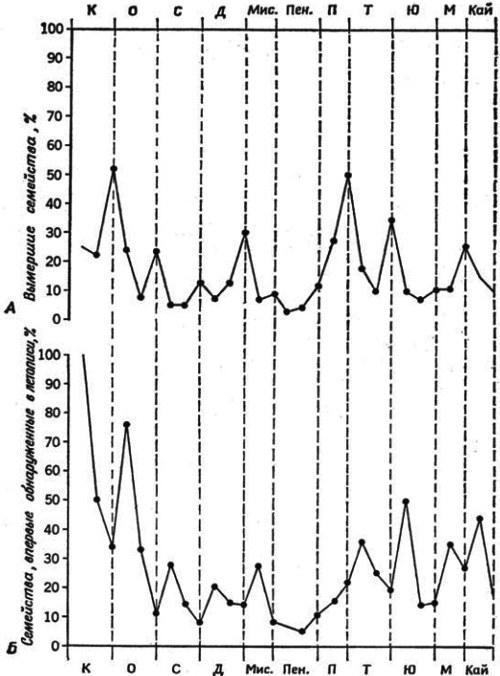
Щоб відповісти на це питання, розглянемо деякі інші випадки переходу групи в подібні умови — чи не спостерігатиметься і там прискорення еволюції та її схожість з інадаптацією, тобто збільшення масштабу змін у поєднанні з їх односторонністю, незбалансованістю. Перехід у відносно вільний, незаповнений екологічний простір характерний, наприклад, для ранніх етапів еволюції багатьох таксонів, і палеонтологічна летопис дійсно підтверджує, що на ранніх етапах еволюції таксона зростає як частота, так і масштаб еволюційних змін, так і інтенсивність відмирання новоутворених інадаптивних груп у міру заповнення ніші. Відомо, що масові вимирання і диверсифікації часто відбуваються майже одночасно. Більше того, періоди диверсифікації слідують за періодами вимирання (рис. 3), що підтверджує той факт, що звільнення екологічного простору вимерлими групами стимулює еволюцію збережених. Прискорення еволюції на ранніх етапах історії таксона більш конкретно демонструє факт приблизного збігу віку роду і багатьох його видів (Шварц, 1980); це показує, що значна частина дивергенції відбувається на найранніших етапах еволюції виниклого таксона.
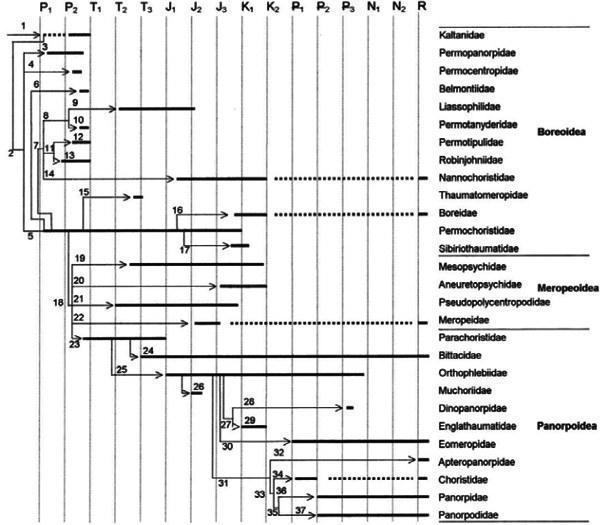
Закон архаїчного різноманіття (Мамкаєв, 1968) описує різко підвищену змінливість організації на ранніх етапах еволюції таксона, через що види і роди в цей час можуть різнитися за ознаками, пізніше характерними для сімей і родів. Не менш характерна для етапу архаїчного різноманіття і мала еволюційна стійкість ранніх членів таксона, що відображається у великій кількості коротких базальних гілок майже на будь‑якій філогенетичній схемі, побудованій з використанням багатого палеонтологічного матеріалу (рис. 4).
Все перелічене дозволяє зробити висновок, що перехід групи в умови обідненого біоценозу і незаповненої екологічної ніші дійсно провокує еволюцію, причому еволюцію суттєво інадаптивну. У такому випадку становлення евадаптивних груп, тобто гармонійних, пристосованих до заповнених біоценозів і напружених конкурентних відносин, можна пов’язати з подальшим жорсткішим умовами і інтенсивним вимиранням більшості новоутворених груп (тих, що виявилися нездатними достатньо швидко перетворитися з інадаптивних у евадаптивні).
Проаналізуємо знамениті експерименти Г. Х. Шапошникова (1961, 1965, 1966, 1978). У цих експериментах тлі Dysaphis anthrisci maicopica Shap. пересаджували з придатної кормової рослини Anthriscus nemorosa MB спочатку на малопридатну (Chaerophyllum bulbosum L.), а потім на раніше зовсім непридатну (Ch. maculatum Wild.). У результаті тлі за кілька поколінь не лише змінилися морфологічно, а й набули репродуктивної несумісності зі своїм видом і неповної сумісності з D. chaerophyllina Shap., істинним споживачем Ch. maculatum. У експериментах Шапошникова відбір явно був одностороннім і незбалансованим (інадаптивним). Дійсно, у досліді здатність харчуватися на новій рослині‑господарі на якийсь час виявилася єдиним життєво важливим комплексом адаптації, а всі інші відступили на задній план. Звісно, експеримент не був завершений, але якби його продовжили і перенесли в природу, ми, ймовірно, могли б спостерігати і наступний етап еволюції. А саме випробування зароджуваної групи всебічним, евадаптивним відбором, для якого важливі всі аспекти адаптації — живлення, розмноження, індивідуальна стійкість різних стадій онтогенезу в умовах різної густини популяції, конкурентоспроможність тощо. Трохи шансів, що конкретна популяція витримає це випробування, але вони є.
Повернемося до проблеми м’яких умов і ослабленого відбору. Чи можна вважати умови експериментів Шапошникова жорсткими? На різних етапах експерименту смертність личинок коливалася від 15‑22 % до 53‑75 %. Здавалося б, висока смертність, але за вказаним показником плодючості (36,6 личинок від однієї партеногенетичної самки; дані лише для кінцевого етапу досліду) вона означає виживання щонайменше близько 10 личинок у потомстві кожної самки і, відповідно, майже десятикратний приріст популяції в кожному поколінні. Іншими словами, смертність у цих дослідах була досить помірною — очевидно, через те, що турботою експериментатора вороги тлей і інші несприятливі впливи були по можливості усунуті. Не дивно, що лінії в експерименті виявилися дуже стійкими і змогли підтримуватись протягом 50 поколінь — термін, який здається малоймовірним для окремих ліній у природних умовах. Що ж до самого факту пересадки на малопридатну і зовсім непридатну рослину, то він саме не є чимось незвичним. У природі тлі часто опиняються в подібному становищі: то крила самка занесена вітром у невідповідний біотоп, то дощ чи вітер скидають безкрилу тлю на іншу рослину. Просто в природі це важко помітити, бо там, на відміну від лабораторії, комаха не на своєму місці зазвичай гине.
Ми рідко замислюємось, говорячи про жорсткі, катастрофічні та подібні умови. Рівноважна густина популяції відповідає тим її значенням, при яких смертність зрівноважує репродуктивний потенціал популяції. Це означає, що з усього потомства кожної самки в середньому лише одній її дочці вдається залишити своє потомство («закон Бекетова»: А. Н. Бекетов, 1860; рік раніше про те ж писав, наприклад, Ч. Дарвін, 1991: 69). Тому елімінація в популяціях у нормі (за винятком коротких і більш‑менш зрівноважуваних один одного епізодів коливань чисельності) практично дорівнює плодючості, тобто близькій до максимуму.
Але якщо практично повна елімінація потомства є нормою існування популяції, якщо шанси залишити потомство для кожної особини завжди мінімальні, якщо вимирання цілих популяцій — звичне явище навіть у екологічному (доеволюційному) масштабі часу, що ми маємо на увазі, говорячи про погіршення умов існування?
Схоже, що жорсткість умов на популяційному рівні в природі взагалі не буває. Навіть у застосуванні до видового і надвидового рівня про це важко говорити. Тут погіршення умов може означати лише тривіальне (у еволюційному масштабі часу) вимирання видів, вплив якого на еволюцію буде відчутний лише як звільнення екологічного простору і, відповідно, пом’якшення умов. Але природа не терпить порожнечі, а екологічний простір особливо, і виживші види миттєво заповнять порожнечу. Вони інтенсифікують процес розмноження збережених популяцій і тим самим зведуть до нуля короткочасне пом’якшення умов їх існування.
Отже, еволюційно значуще пом’якшення умов існування настільки ж сумнівне, як і їх жорсткість. Пом’якшення умов саме по собі означає лише підвищення виживаності і, відповідно, зростання популяції. При цьому мальтусова геометрична прогресія розмноження за кілька поколінь призведе до насичення екологічного простору і відновлення нормальних для особини і популяції умов, включаючи нормальну (рівноважну з плодючістю, тобто майже стопроцентну) смертність. Зміна характеристик популяції внаслідок коливань її густини («хвилі життя») обмежується лише змінами в межах нормально‑внутрішньовидової змінливості. Той факт, що коливання густини популяції є найпростішим явищем, і при цьому випадків породженого ним незворотного еволюційного зміни досі не описано, доводить, що для еволюції це явище не суттєве.
Отже, ні пом’якшення, ні жорсткість умов існування не представляють реальних факторів еволюції. Що ж у такому випадку може подолати стійкість збалансованого епігенотипу і успішно вивести організацію живого організму за межі історично зумовленої норми?
Причина стійкості збалансованого епігенотипу полягає в колись досягнутому компромісі між протилежними потребами оптимізації різних адаптивних функцій організму. Ці потреби протилежні в тому сенсі, що через складне переплетення морфогенетичних і функціональних зв’язків між усіма частинами і ознаками організму адаптивне посилення однієї функції має відбиватися втратою ефективності інших. Ослаблення цілісності онтогенезу, що полегшує незалежні перетворення окремих систем, звичайно, не лише можливе, а й має місце в природі. Однак цей напрямок (спрощення організації і загальна дегенерація) досить специфічний, вимагає особливих умов і не є ні характерним, ні особливо поширеним. Зрештою, щоб втратити висоту організації, спочатку її треба набути. Ослаблення цілісності як механізм еволюції існує, але для пояснення еволюційного процесу його явно недостатньо.
Тому нам необхідно реконструювати умови, у яких зміна організації відбувається за збереженої або зростаючої цілісності, або, швидше, коли тимчасове зниження цілісності швидко відновлюється. Для цього спробуємо уточнити метафору адаптивного компромісу. Головну проблему для еволюції представляє, як ми бачили, протиріччя між адаптивними функціями в процесі їх оптимізації. Протиріччя створюється, з одного боку, тісними онтогенетичними і функціональними зв’язками між системами забезпечення цих функцій, а з іншого — жорстким контролем за їх належним виконанням. Ми бачили, що ослаблення зв’язків можливе, але недостатньо для пояснення еволюції. Загальне ослаблення селективного контролю над належним виконанням адаптивних функцій, як ми бачили, теж неможливе: у еволюційному масштабі часу елімінація завжди наближається до максимально можливих значень. Де ж вихід?
Підказкою тут служать обговорювані ситуації, що провокують швидку еволюцію. Спільне для них — потрапляння популяції в нові, незвичні умови. Найбільш очевидні для нас експерименти з тлями показують, у чому саме полягає незвичність нових умов. Там експериментатор пред’явив тлям дуже жорсткі вимоги за одним або кількома параметрами (у даному випадку — здатність харчуватися на невідповідній рослині), але при цьому звільнив їх від жорсткого середовищного контролю за іншими параметрами: захистив від негоди, хижаків, паразитів, хвороб тощо.
Мені не відомі дослідження, що аналізують подібний розподіл інтенсивності селективного контролю у первісних поселенців на островах або в нових екологічних ніша́х. Однак загальні міркування щодо низької напруженості конкурентних відносин в островних умовах порівняно з континентальними цілком правдоподібні. Вони підтверджуються відносно високим імміграційним потенціалом континентальних видів на островах і надзвичайно низьким у зворотному випадку. У таких умовах цено́тичний контроль виконання деяких функцій (наприклад, функції захисту від хижаків і спеціалізованих паразитів, які в островних спільнотах часто є дефіцитними) може бути знижений на користь посилення контролю в інших напрямках. Зрештою, умови саме нові, щоб розподіл селективних вимог був іншим, ніж раніше, і, зокрема, щоб деякі функції контролювалися значно жорсткіше, ніж у попередніх умовах. Якщо популяція при цьому вижила, значить, сумарна елімінація стабільно не перевищувала її репродуктивний потенціал і, відповідно, селективний контроль інших функцій був знижений.
Отже, еволюцію, ймовірно, провокує лише одностороннє пом’якшення середовищного контролю організації, при якому лише деякі з адаптивних функцій залишаються під жорстким спостереженням відбору. Очевидним наслідком такого одностороннього (незбалансованого) контролю є розбалансування, дестабілізація склалої організації, і вона дійсно проявляється у подібних випадках (Жерихин у: Діяльність..., 1967; Беляєв, 1974; Шишкін, 1984, 1987; МсСипе, 1990 тощо).
Оскільки при дестабілізації адаптивність системи неминуче знижується, перебудова навряд чи може відбуватись у нормальних умовах. З цього, між іншим, випливає, що новоутворена група навряд чи зможе вкоренитися в уже зайняту нішу і витіснити її власника. Більш правдоподібно, що вона або займе нішу, звільнену внаслідок вимирання її колишнього мешканця, або зможе створити (відкрити) нову нішу. Можливо, саме тому нам відомо досить багато правдоподібних прикладів недавнього виникнення нових таксонів в островних умовах, а також при антропогенній інтродукції. Особливо вражають гельмінтологічні приклади (цитовані в дискусії за доповіддю Rausch, 1982: 184) про два послідовних акти видоутворення за 300 років (у паразита домашньої миші, завезеної в Північну Америку) і про дивергенцію двох видів людських ленців не більше 16 тис. років тому. А навколо нас таких прикладів дуже мало, хоча в звичайних умовах і біорізноманіття, і його вивчення значно вищі, і, відповідно, більша ймовірність виявлення випадків природного видоутворення.
Сказане дозволяє припустити, що еволюція на видовому і, особливо, на вищих таксономічних рівнях можлива лише в результаті такої зміни умов, при якій діючий на популяцію відбір стає суттєво одностороннім. Тільки такий відбір здається здатним подолати стійкість попереднього добре збалансованого адаптивного компромісу. Однак новоутворена група, пройшовши дестабілізацію і втративши колишню збалансованість, повинна пройти жорсткий всебічний контроль (відбір за всіма адаптивними функціями) і під його дією відновити свою стійкість. Іншими словами, еволюційна зміна повинна пройти через два етапи — дестабілізацію і потім стабілізацію новоутвореного епігенотипу.
Як уже згадувалося, результати цих подій можна ототожнити відповідно з інадаптивною і евадаптивною зміною організації. Іншими словами, у процесі еволюції організація живих істот проходить три послідовних етапи: інадаптацію, евадаптацію і стазис. Якщо, звичайно, не станеться вимирання, що можливо на будь‑якому етапі, хоча найбільш ризикованим у цьому плані, природно, є процес евадаптації, а найспокійнішим — стазис. Перші два етапи, як найуразливіші, мають бути короткочасними, а стазис може тривати неоприділену довжину. Вихід зі стазису навряд чи може бути спонтанним: причиною має стати зміна умов. Якщо така зміна викликає зростання загальної елімінації понад репродуктивний потенціал, неминуче вимирання популяції (і виду, якщо вимерли всі його популяції, і вищого таксона, якщо те ж сталося зі всіма його видами). Якщо ж умови існування змінюються так, що контроль організації стає одностороннім, не виключено, що популяція зможе увійти в процес інадаптивного перетворення її збалансованого епігенотипу. А чи зможе вона успішно пройти його, і тим більше — пройти надзвичайно ризикований процес евадаптації, це вже справа випадку, причому випадку дуже рідкісного.
Тут треба зазначити ще одну важливу рису нашої моделі — її авторегуляторність.{
"translated_text": "Як би високо не піднялася стійкість (забуференність) епігенотипу навіть помітної частини жителів нашої планети, катаклізми навряд чи серйозно загрожуватимуть різноманіттю життя на Землі, якщо, звичайно, мати на увазі еволюційно значущі відрізки часу. Навіть якщо помітна частина біорізноманіття «закоснеє» у досконалості і, отже, жорсткості своїх епігенотипів і виявиться суттєво «прорідженою» черговою катастрофою (субглобальним оледенінням, астероїдною чи ядерною зимою або подібним апокаліпсисом), нічого страшного для біосфери не станеться. Виживші види потраплять у ситуацію неповних спільнот із суттєво ослабленими конкурентними відносинами. Це буде провокувати швидку інадаптивну еволюцію, і тим швидшу, чим ширша була «прополка». У міру заповнення спільнот багато з тих, хто витягнув виграшний квиток, його втратять, але чим більша частка невдачників, тим більше шансів у тих, хто зможе продовжити трохи довше. Священне місце порожнім не залишиться, воно заповниться. Правда, нові спільноти можуть виявитися мало схожими на попередні.\nЯк відомо, навіть у околицях межі перм і триасу вимирання було хоча й масштабним, але не подавляючим: зазвичай говорять, що з середини верхньої перми було втрачено порядку половини сімей і трьох чвертей родів у морях і суттєво менше на суші (рис. 5). Проте далі світ, відновившись приблизно до середини триасу, виявився дуже суттєво іншим, причому не лише за складом, а й за тенденціями. Сумарне різноманіття, досягнуте в морях у другій половині ордовика, далі до середини верхньої перми (двісті мільйонів років, тобто лише трохи менше, ніж мезозой і кайнозой разом!) не зростало і лише коливалося навколо пізньоордовикського рівня. Після пермотріасового кризису ріст відновився і триває до сьогодні, вже багаторазово перекривши пізньопалеозойський рівень стабілізації (рис. 5). Очевидно, сама структура морських спільнот радикально змінилася, давши місце значно більшому зростанню біорізноманіття.\n
![Рис. 5. Зміна різноманіття організмів моря [і суші] у фанерозої: порядки (1), сімейства (2) і роди (3) морських тварин (N/NR - відношення числа відповідних таксонів на нижній межі епохи до числа сучасних таксонів, відомих у реціентному стані) (за](https://s3.prozo.re/batrachos/images/rasn-5_tyMUMRJ.original.jpg)
}
1.2. МЕТОДОЛОГІЯ ФІЛОГЕНЕТИКИ, ТАКСОНОМІЇ ТА НОМЕНКЛАТУРИ
1.2.1. ФІЛОГЕНЕТИКА
1.2.1.1. Аналіз груп
1.2.1.2. Аналіз ознак
1.2.1.2.1. Аналіз різниць
1.2.1.2.2. Аналіз схожості
1.2.1.3. Комп’ютерна кладистика