Мы используем аналитические cookie только чтобы считать посетителей и оценивать нагрузку — и никогда не продаём ваши данные. Они загружаются только с вашего согласия.
Privacy policy
Штёк и др., 2011. Одновременная менделевская и клональная передача геномов...
Неофициальный рабочий перевод статьи, представляющей безусловный интерес. Stöck M., Ustinova J., Betto-Colliard C., Schartl M., Moritz C., Perrin N. Simultaneous Mendelian and clonal genome transmission in a sexually reproducing, all-triploid vertebrate. Proc. R. Soc. B, 2011. doi:10.1098/rspb.20...
Дмитро Шабанов
Неофіційний робочий переклад статті, що представляє значний інтерес. Stöck M., Ustinova J., Betto‑Colliard C., Schartl M., Moritz C., Perrin N. Simultaneous Mendelian and clonal genome transmission in a sexually reproducing, all‑triploid vertebrate. Proc. R. Soc. B, 2011. doi:10.1098/rspb.2011.1738 Руський переклад назви Bufo baturae — пуштунська жаба — використовується за порадою С. Н. Литвинчука і має попередній характер. Одночасна менделєвська і клональна передача геномів у розмножуваних статевим шляхом повністю триплоїдних хребетних. Мейоз у триплоїдних особин, здавалося б, стикається з непереборними труднощами при діленні непарного числа хромосомних наборів на два. Триплоїдні хребетні зазвичай обходять цю проблему за допомогою або асексуальності, або деяких форм гибридогенезу, у тому числі мейотичного гибридогенезу, який включає репродуктивне співтовариство різної плоїдності і з різними наборами геномів. Пуштунські жаби (Bufo baturae; 3n = 33 хромосом), однак, представляють повністю триплоїдне статеве розмноження. Цей гибридний вид має дві копії геному, що містить ядерцевий організатор (NOR +) на хромосомі 6, а третій геном без нього (NOR—). Самці виробляють лише гаплоїдну NOR + сперму, тоді як яйцеклітини диплоїдні і містять один набор NOR + і один набор NOR—. Тут ми проводимо родинний аналіз за кодомінантними маркерами мікросателітів (1) для підтвердження чисто клональної та материнської передачі набору NOR—, і (2) щоб продемонструвати менделєвську сегрегацію і рекомбінацію наборів NOR + у обох статей. Це новий спосіб розмноження у хребетних («попередньо вирівнювальний гибридний мейоз») пропонує ідеальну можливість для вивчення еволюції нерекомбінантних геномів. Виявлення механізмів, які дозволяють одночасну передачу двох геномів, одного менделєвського, іншого клонального, може пролити світло на загальні процеси, які регулюють мейоз у хребетних. 1. Вступ За другим законом Менделя [1] алелі різних генів рекомбінують незалежно один від одного під час гаметогенезу. У статево розмножуваних видів випадкова передача батьківського і материнського геномів досягається за рахунок незалежної сегрегації хромосом під час мейозу. Деякі геноми тварин, однак, демонструють відхилення передачі, часто залежно від того, від якого батька вони походять, і, як правило, внаслідок стародавньої гибридизації [2]. Гібридні лінії мурах, наприклад, несуть два незалежно розвиваються геноми, які передаються або мейотично [3], або клонально [4,5]. Подібні процеси відбуваються у гибридогенетичних хребетних: при диплоїдному гибридогенезі один геном передається клонально через гибридні лінії, а інший — статевим шляхом від одного з батьківських видів [6]. При мейотичному гибридогенезі обидва гибридизуючі геноми можуть передаватися статевим шляхом, через схрещування диплоїдних і триплоїдних гибридів з різними наборами геномів (рис. 1, [14,16]).
Рисунок 1. Способи розмноження триплоїдних хребетних. Показані геноми батьків, гамет і потомства (рядки) при різних способах репродукції (стовпці). A, B: геноми різних батьківських видів. Напівжирні кольорові символи вказують на клонально передані копії, тоді як звичайні чорні символи з індексами позначають різні (рекомбінантні) копії. Істинний партеногенез: клональний (самці відсутні), виключно у рептилій [7,8]; Спермозалежний партеногенез (т. з. гіногенез): клональний, ембріогенез вимагає запуску аллоспецифічними сперматозоїдами, які не включаються (при рідкісному «батьківському просочуванні» може передаватися субгеномна кількість батьківської ДНК), відбувається у кісткових риб і хвостатих амфібій [9]; Клептогенез: самки отримують, повністю або частково, геноми від своїх партнерів за не до кінця зрозумілими механізмами, що дозволяє їм очищати геном від шкідливих алелей (тут BB), описаний у хвостатих амфібій [10]; Неназвана форма гибридогенезу: клональні диплоїдні яйцеклітини запліднюються спермою рекомбінуючих видів зі статевим розмноженням, які можуть бути диплоїдними або триплоїдними (як при мейотичному гибридогенезі), зустрічається у безхвостих амфібій і кісткових риб [11‑13]; Мейотичний гибридогенез: може відбуватися у триплоїдних самців і/або самок; виявлений у кісткових риб і безхвостих амфібій [14,15]; підвищення плоїдності диплоїдного потомства, яке може виробляти гибридні диплоїдні гамети, може відбуватися в наступному поколінні (стають, наприклад, ABB′) для відновлення триплоїдності (за аналогією з попередньою формою гибридогенезу); Попередньо вирівнювальний гибридний мейоз: відбувається у пуштунській жабі: обидві статі триплоїдні і демонструють менделєвську сегрегацію і рекомбінацію в геномах B (еквівалент NOR + даної статті), тоді як геном A (т. з. NOR—) клонально передається від матері. Двостатеве розмноження чистих триплоїдів ускладнює проблема рівного розподілу трьох наборів хромосом у мейозі [17], см. огляд [18]. Гібриди Poeciliopis, наприклад, є гибридогенетичними у своїх диплоїдних формах, але стають гіногенетичними як триплоїди [19,20]. В альтернативі гіногенезу або партеногенезу (рис. 1, [7‑9,21]) деякі триплоїдні хребетні поєднують клональні і статеві елементи у своєму способі розмноження, наприклад, у клептогенезі або різних формах гибридогенезу [10,22‑24], у тому числі мейотичному гибридогенезі, який вимагає репродуктивного співтовариства різного рівня плоїдності і складу геномів. У цьому контексті пуштунські жаби (Bufo baturae) особливі, бо є триплоїдами, що розмножуються статевим шляхом [25]. Цей гибридний вид, що походить з високих гір на півночі Пакистану (понад 1500 м над рівнем моря), має геном (3n = 33; [25,26]) з двох наборів хромосом, що несуть ядерцевий організатор (NOR +) на хромосомі 6, і інший набор без такого регіону (NOR—; рисунок 2). Самці виробляють лише гаплоїдну NOR + сперму, ймовірно, усуваючи набір NOR— (11 хромосом) до початку мейозу. На відміну від цього, яйцеклітини диплоїдні (2n = 22) з одним набором NOR + і одним набором NOR—. У незрілих ооцитах виявлено 22 бівалентів хромосом типу лампових щіток [25]. Таким чином, було припущено, що набір NOR— передається чисто по материнській лінії і клонально. Проте залишилося невідомим, чи рекомбінаує набір NOR + і як це відбувається. Для самок можна висунути дві гіпотези (рис. 2): (а) Один набір NOR + елімінується (випадково або залежно від походження батьків), після чого в процесі ендоміозу автодуплікуються два залишкові набори. Мейоз у такому випадку відбувається між псевдобівалентами [27] і продукує один або максимум два класи у випадку клональних диплоїдних яйцеклітин. Альтернатива (б) — набір NOR— автодуплікується до мейозу, під час якого два набори NOR + нормально рекомбінаються.
Рисунок 2. Схема репродуктивної системи у триплоїдних пуштунських жаб з гіпотетичними механізмами (а) і (б) оогенезу та (c) і (d) сперматогенезу. Синій NOR— символ: нерекомбінантний (клональний) хромосомний набор без ядерцевих організаторів. Червоний або пурпуровий NOR + символи: різні NOR‑несучі набори хромосом. Змішані червоний і пурпуровий NOR + символи: рекомбінаційні NOR‑несучі набори. Механізми, підтверджені в даному дослідженні, вкладаються в рамку. Для самців, аналогічно, перша гіпотеза (рис. 2, c) полягає в тому, що весь материнський внесок (NOR +, NOR—) виключається, а потім батьківський NOR + дуплікується внаслідок премейотичного ендоміозу. Мейотичні пари (NOR +/NOR +; [25]) таким чином представляють псевдобіваленти, передбачаючи клональну передачу одного набору NOR +. Сперматоцити будуть складатися з одного полілокусного генотипу (або максимум двох, якщо елімінація NOR + відбулася випадково). Альтернатива (рис. 2, d) — усунення лише набору NOR— і проходження нормального мейозу та рекомбінації двома наборами NOR +. За даними мульти‑локусного фінгерпринту Шток та співавтори [25] визначено кілька генотипів серед потомства однієї пари. Однак домінантні мульти‑локусні маркери не завжди чітко інтерпретовані, і таким чином не дозволяють прояснити основний механізм. У даній роботі ми провели аналіз груп потомків за 15 кодомінантних локусів мікросателітів, щоб встановити характерні для геномів моделі передачі та сегрегації. Наші результати ясно підтверджують чисто клональну материнську передачу одного набору хромосом (NOR—) і показують незалежну сегрегацію та рекомбінацію двох інших наборів (NOR +) у самців і самок. Ці перестановки генетичного матеріалу мають забезпечити ефективне очищення двох наборів хромосом NOR +, як при нормальному статевому розмноженні. Це перший приклад паралельної клональної і мейотичної передачі хромосомних наборів в межах однієї лінії хребетних. 2. Матеріали та методи Тварини, використані в контрольованих експериментальних схрещуваннях, були відловлені в трьох населених пунктах на півночі Пакистану (електронний додатковий матеріал, таблиці S1) протягом трьох періодів польових робіт (червень‑липень 1996, 1997 і 2000). Ми провели п’ять експериментів з розведенням триплоїдних пуштунських жаб. Крім того, ми схрестили одну самку пуштунської жаби як з диплоїдним самцем Bufo variabilis з Сирії (2n = 22, з двома наборами NOR +), так і з тетраплоїдним самцем Bufo oblongus з Ірану (4n = 44, що містить два набори NOR + і два набори NOR—; [28]). Двадцять з 100 потомків були вирощені в резервуарах до личинки довжиною 2‑3 см (етапи Госнера від 30 до 38, [29]). Всього у 85 головастиків з семи схрещувань були обрані зразки для генетичного аналізу. Головастики були або кариотиповані, або їх плоїдність визначали за допомогою проточної цитометрії; ДНК виділяли, як описано у Штока та співавторів [30]. Ми протестували серії мікросателітних маркерів з генетичної бібліотеки повторюваних елементів пуштунських жаб, деякі з яких раніше використовувалися для інших видів (електронні додаткові матеріали, таблиці S2). Алелі були ампліфіковані, марковані GeneMapper v. 3.7 (Applied Bio‑systems) і названі залежно від їх довжини в парах основ, як описано [30]. Алелі від наборів NOR + і NOR—, а також нульові алелі (0) були визначені за характером успадкування (див. § 3). Аналіз зв’язку проводився в GENEPOP (https://genepop.curtin.edu.au/, [31,32]) з параметрами за замовчуванням, і потенційні групи зв’язків перевірялися візуальним оглядом. Враховуючи розмір набору даних, генотипи потомства були візуально оглянуті на випадки рекомбінації. Кількість рекомбінацій була нормована щодо числа інформативних подій у парі, і відхилення від випадкової сегрегації тестувалися на значущість (за допомогою X‑тесту). 3. Результати (а) Характер успадкування у пуштунській жабі В цілому 15 пар праймерів мікросателітів ампліфіковано в одній або кількох сім’ях. П’ять з них (D103, D105, D5, C224 і C203, електронні додаткові матеріали, набір даних S1) відображають три алелі на особину, які передбачають ампліфікацію продуктів і з NOR—, і з двох наборів NOR +. Алелі NOR— було легко визначити за постійною гомоморфністю серед потомства від пари, ідентичною материнській копії і відмінною від батьківської в тих випадках, коли батьківські копії різнилися (таблиця 1 та додаткові електронні матеріали, таблиця S3 і набір даних S1). Обидва набори NOR +, навпаки, відображають успадкування від обох батьків і менделєвську сегрегацію (додаткові електронні матеріали, таблиця S3 і набір даних S1). Кожен гетерозиготний батько передавав дві свої алелі з рівною ймовірністю (біноміальні тести). 10 інших маркерів представлено максимум двома алелями на особину, з успадкуванням від обох батьків і менделєвською сегрегацією, відповідно до очікувань для мейотичних наборів NOR + (додаткові електронні матеріали, набір даних S1). Таблиця 1. Патерн успадкування алелей NOR— у п’яти локусах (рядки) у п’яти 3n парах (стовпці). Вказано кількість потомства з материнським алелем NOR—/кількість інформативних подій. ‘ni’ означає неінформативність, (—) означає, що аналіз не проводився. По всіх локусах і парах ми підрахували 139 випадків материнського успадкування з 139 інформативних подій. Повні дані представлені в електронному додатковому матеріалі, набір даних S1.
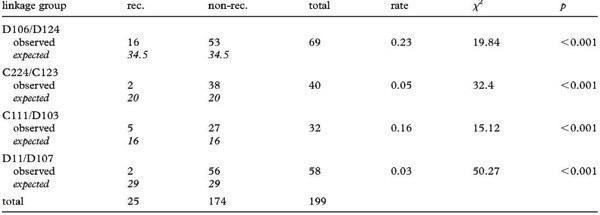
Чотири групи зв’язку можуть бути визначені за участю двох маркерів кожна (таблиця 2). З 199 інформативних подій ми виявили в цілому 25 випадків рекомбінації (таблиця 2), що відбувалися у обох статей (електронні додаткові матеріали, набір даних S1 і таблиця S3). Усі інші пари маркерів передавалися незалежно одна від одної, породжуючи велике різноманіття полілокусних генотипів у сім’ї. Цікаво, що п’ять маркерів, що ампліфікували продукти NOR—, належали до різних груп зв’язку в геномі NOR + (табл. 2 і електронні додаткові матеріали, набір даних S1), підтримуючи геномне поширення маркерів NOR—. (b) Міжплоїдні схрещування Локус D105 також може бути ампліфікований у потомстві самки B. baturae і (i) диплоїдного самця B. variabilis (2n = 22, що має два набори NOR +) і (ii) тетраплоїдного самця B. oblongus (4n = 44, включаючи два набори NOR + і два набори NOR—). Все потомство від осіменення самцем B. variabilis було триплоїдним і успадкувало материнську алель NOR— у локусі D105, а два набори NOR + продемонстрували успадкування від обох батьків з менделєвською сегрегацією у обох. Потомство від запліднення B. oblongus було тетраплоїдним і представляло чотири алельні копії у локусі D105, що відповідали двом набором NOR— і двом набором NOR +. Одна алель NOR— була ідентична материнській копії, інша випадково успадкована від двох батьківських копій NOR—. Два набори NOR + також показали успадкування від обох батьків, з менделєвською сегрегацією у обох. Отже, в обох схрещуваннях пуштунська жаба‑мати виробляла 2n ооцити з клонально переданим NOR— і рекомбінантним NOR +. Самець B. variabilis виробляв сперму з гаплоїдним рекомбінованим NOR +, тоді як самець B. oblongus виробляв диплоїдні сперматозоїди з рекомбінантними наборами NOR + і NOR— (див. також [25,33]). Таблиця 2. Модель рекомбінації в чотирьох групах зв’язку. Rec., рекомбінація спостерігалася; non‑rec., рекомбінація не спостерігалася. Значення, очікувані за умовою незалежності успадкування, курсивом. Total вказує загальну кількість інформативних подій за парами і статями.
4. Обговорення Наші результати показують чисто материнську і клональну передачу всіх маркерів NOR—. Потомство успадковує лише материнські копії п’яти маркерів (D5, D103, D105, C224 і C203), які ампліфікуються в алелі NOR—. Як буде показано, ці п’ять маркерів локалізовано в різних групах зв’язку на геномах NOR +, що підкріплює припущення, що весь набір хромосом NOR— підлягає клональній материнській передачі. Крім того, наші результати свідчать про рекомбінацію NOR + у обох статей. 15 маркерів, які ампліфікували алелі NOR +, були розподілені по 11 різних групах зв’язку (відповідних гаплоїдній кількості хромосом у пуштунській жабі), що відображає випадкову і незалежну сегрегацію у обох статей (електронний додатковий матеріал, набір даних S1). Гетерозиготні дорослі завжди передавали дві свої алелі з рівною ймовірністю, що вказує на випадкову сегрегацію батьківського і материнського набору хромосом NOR + у обох статей (рис. 2: шляхи (б), (г)). Крім того, рекомбінація також відбувалася у обох статей між локусами з однієї і тієї ж групи зв’язку (таблиця 2). У підсумку наші результати виключають гіпотезу (а) для оогенезу (передбачаючу клональне виробництво ооцитів NOR +/NOR—) і (c) для сперматогенезу (передбачаючу клональне виробництво сперми NOR +). Ці результати піднімають важливі питання щодо подібних механізмів і еволюційних наслідків, пов’язаних з цим незвичним способом розмноження. (a) Схожі механізми NOR— геном пуштунських жаб елімується у самців, але дуплікується у самок до мейозу. Премейотична елімінація і/або дуплікація геномів вже описані у гибридогенетичних хребетних, таких як зелені жаби Rana (Pelophylax) esculenta, гибрид між Rana ridibunda (RR) і Rana lessonae (LL) [33,34]. Коли самки R. esculenta (RL), що живуть з R. ridibunda, втрачають батьківський геном (R) з зародкової лінії і одночасно подвоюють свої L‑геноми премейотичним ендоміозом, наступний мейоз включає повністю гомозиготні псевдобіваленти (LL) і продукує нерекомбінантні гаплоїдні (L) ооцити [35]. Схрещування з самцем R. ridibunda відновлює геном RL. Таким чином, один набір хромосом (R) рекомбінає у батьківських видах, тоді як інший набір (L) передається клонально у гибридів. Обидва набори хромосом рекомбінають при мейотичному гибридогенезі [15,36], який включає репродуктивні спільноти гибридів різної плоїдності і геномного складу (RRL, LLR і RL). Триплоїди RRL видаляють L‑геном з зародкової лінії і продукують рекомбінатні гаплоїдні R‑гамети під час нормального мейозу. Триплоїди LLR аналогічно видаляють R‑геном до мейозу, продукуючи рекомбінатні гаплоїдні гамети L. Нарешті, RL‑диплоїди формують клональні диплоїдні гамети RL після ендоміозу [11]. Поєднання цих гамет відновлює вихідні диплоїдні і триплоїдні геноми [36‑38].Подібні механізми були зафіксовані у кількох гібридогенетичних кісткових риб [39-44]. Таким чином, премейотичне усунення NOR— у самців пуштунських жаб, з подальшим нормальним диплоїдним мейозом двох наборів NOR+, має схожість з деякими процесами, що відбуваються під час мейотичного гібридогенезу та інших форм гібридогенетичного або клептогенетичного способу розмноження, відомого у триплоїдних хребетних (рис. 1). Крім того, дуплікація NOR— у самок пуштунської жаби відбувається через гаметогенетичний механізм (премейотичний ендомітоз), який добре відомий у патреногенетичних і гібридогенетичних хребетних [23,45,46]. Проте система пуштунської жаби здається унікальною серед хребетних тим, що (i) процес мейозу різниться у різних статей, і (ii) самки одночасно передають один геном, який клонально дуплікується, і інший, який проходить нормальний мейоз. Найближча система, схоже, була виявлена у рослин, таких як вереск і шипшина, у яких пилок передає лише геном, відтворений статевим шляхом, тоді як яйцеклітини додатково передають клонально відтворений геном [47-49]. Однак у цих випадках не відбувається премейотична дуплікація клональних геномів. За нашими даними, автодуплікація одного повного набору хромосом (NOR—) у присутності інших диплоїдних геномів (NOR+/NOR+', які залишаються прейотично незмінними і пізніше передаються менделєвським шляхом) досі не продемонстрована (рис. 1). (b) Еволюційні аспекти Пуштунські жаби демонструють дивовижну однорідність (в межах виду) мітохондріальних послідовностей [50], що дозволяє припустити унікальне та недавнє походження (хоча жоден із його потенційних батьківських видів не зустрічається в межах ареалу виду, який включає водозбірні басейни трьох річок північного Пакистану). Хоча триплоїди можуть виникати безпосередньо при схрещуванні диплоїдних і тетраплоїдних батьківських видів, комплекс мейотичних процесів, описаних тут (включаючи дуплікацію цілого геному у самок і його елімінацію у самців), не міг виникнути одразу при первинній гібридизації. Проміжні етапи могли включати періодичні гібридні взаємодії між лініями різної плоїдності та геномного складу, подібні ситуації спостерігаються зараз у північній Киргизії, де деякі триплоїдні самці, що виникають у результаті природних схрещувань 2n Bufo turanensis самок з 4n Bufo pewzowi самцями, здійснюють зворотні схрещування з самками будь‑якого з батьківських видів [30]. Однак такі механізми мають бути рідкісними через низьку ймовірність зустрічі комплексу генетичних умов, необхідних для досягнення стабільних гібридних комбінацій клональних і менделєвських геномів, що останнім часом також передбачається для старих ліній гіногенетичних риб [51]. Цей спосіб розмноження також піднімає питання щодо селективних процесів і еволюційної долі. Поліплоїдні (3n, 4n) лінії зелених жаб, які розвивалися незалежно один від одного кілька разів, явно пов’язані з суворими середовищами існування [50]. Пуштунські жаби, зокрема, живуть в екстремальних умовах висоти та посушливості [52]. Одне з можливих припущень полягає в тому, що клональне розмноження NOR— геному дозволяє зберегти епістатичні компоненти життєздатності, що може мати значення при відборі, що випливає з абіотичних і передбачувано суворих природних факторів [53]. Насправді, безстатеві лінії часто зустрічаються в маргінальних місцевих середовищах з більш екстремальними умовами (холодними, сухими, великих висот і підвищеного УФ‑випромінювання), ніж їх статеві родичі [54-57]. Важливе питання в цьому контексті полягає в тому, чи експресуються обидва геноми, і якщо так, чи є експресія диференціальною (тканеспецифічною), як це спостерігається у аллополіплоїдних риб [58] і рослин [59]. Цей виключний режим NOR— успадкування також має мати несприятливі еволюційні наслідки. По‑перше, чисто материнська передача відкриває можливості для геномних конфліктів. Ми очікуємо, зокрема, що фактори фемінізації розвиватимуться на материнському NOR—, щоб потім балансуватись факторами маскулінізації, що еволюціонують у NOR+, які передаються від обох батьків. Це може призвести до статево‑специфічного зсуву, як це спостерігається у гібридогенетичних R. esculenta, де материнськи переданий клональний геном R несе лише фактори фемінізації [60]. Подібним чином, мутації, які шкідливі лише для самців, не будуть видалятись відбором і можуть накопичуватись, наприклад, як це спостерігається і в інших материнсько‑лінійних мітохондріальних [61] або ядерних [62] геномах. По‑друге, не рекомбінуючий NOR— набір має поступово накопичувати шкідливі мутації внаслідок поєднання сил підвищеного дрейфу, селективної переваги, фонового відбору та «годинника» Мьоллера [63-65], як це відбувається з статевими хромосомами (Y або W), а також не рекомбінуючими геномами геміклональних хребетних [66]. Пуштунська жаба, однак, могла виникнути надто недавно, щоб такі мутаційні кризи або геномні конфлікти через визначення статі могли бути виявлені [25,50]. Навпаки, менделєвська сегрегація і рекомбінація, виявлені в геномі NOR+, мають перешкоджати його еволюційному виведенню, забезпечуючи довгостроковий еволюційний потенціал пуштунських жаб, характерний для розмножуваних статевим шляхом хребетних з нормальним мейозом. Порівняння послідовностей генів з NOR— геному B. baturae з такими його батьківськими видами, а також тетраплоїдними лініями зелених жаб, як B. oblongus і B. pewzowi, де NOR— геноми рекомбінають відповідно до цитологічних [28] і мікросателітних (дане дослідження) доказів, може допомогти отримати інформацію не лише про філогенетичну історію наборів NOR—, а й про характер відбору, що діє на цей не рекомбінуючий геном. Це може також дозволити дослідження потенційних конфліктів у шляхах визначення статі, а також можливих інтергеномних рекомбінацій, зокрема, спостережуваних у клептогенетично розмножуваних амбистах [67]. 5. Висновок Спосіб розмноження пуштунської жаби відрізняється від відомих до сьогодні у інших хребетних (рис. 1) не лише тим, що мейотичні процеси різняться у різних статей, а й тим, що самки демонструють клональне і статеве розмноження одночасно (явище премейотичної автодуплікації зачіпає один хромосомний набір, тоді як інший проходить нормальний мейоз). Тут ми назвали цей процес «попередньо вирівняний гібридний мейоз». Виявлення механізмів, що лежать в основі цих особливостей, могло б пролити світло на загальні процеси регуляції мейозу у хребетних. Пуштунські жаби також представляють інтригуючу можливість порівняти еволюційні сили в рекомбінуючих і не рекомбінуючих геномах у межах одного організму. Література 1 Mendel, G. 1866 Versuche uber Pflanzenhybriden [Experiments on plant hybridization]. Verh. Naturforsch. Ver. Briinn 4, 3-47. [In German.] 2 Keller, L. 2010 Genetics: biased transmission of genomes according to parents of origin. Curr. Biol. 20, R601- R602. (doi:10.1016/j.cub.2010.05.048) 3 Helms Cahan, S. & Keller, L. 2003 Complex hybrid origin of genetic caste determination in harvester ants. Nature 424, 306-309. (doi:10.1038/nature01744) 4 Fournier, D., Estoup, A., Orivel, J., Foucaud, J., Jourdan, H., Le Breton, J. & Keller, L. 2005 Clonal reproduction by males and females in the little fire ant. Nature 435, 1230-1234. (doi:10.1038/nature03705) 5 Kulmuni, J., Seifert, B. & Pamilo, P. 2010 Segregation distortion causes large-scale differences between male and female genomes in hybrid ants. Proc. Natl Acad. Sci. USA 107, 7371-7376. (doi:10.1073/pnas. 0912409107) 6 Schultz, R. J. 1969 Hybridization, unisexuality and poly- ploidy in the teleost Poeciliopsis (Poeciliidae) and other vertebrates. Am. Nat. 103, 605-619. (doi:10.1086/ 282629) 7 Darevsky, I. S. 1958 Natural parthenogenesis in certain subspecies of rock lizards, Lacerta saxicola Eversmann. Dokl. Akad. Nauk SSSR, Biol. Sci. 122, 730. [In Russian.] 8 Kearney, M., Fujita, M. K. & Ridenour, J. 2009 Lost sex in the reptiles: constraints and correlations. In Lost sex: the evolutionary biology of parthenogenesis (eds I. Schoen, K. Martens & P. van Dijk), pp. 447-474. Heidelberg, Germany: Springer. 9 Beukeboom, L. W. & Vrijenhoek, R. C. 1998 Evolutionary genetics and ecology of sperm-dependent parthenogenesis. J. Evol. Biol. 11, 755-782. (doi:10. 1007/s000360050117) 10 Bogart, J. P. & Klemens, M. W. 1997 Hybrids and genetic interactions of mole salamanders (Ambystoma jeffersonianum and A. laterale) (Amphibia: Caudata) in New York and New England. Am. Mus. Novit. 3, 1-78. 11 Polls-Pelaz, M. 1994 Modes of gametogenesis among kleptons of the hybridogenetic water frog complex: an evolutionary synthesis. Zool. Polon. 39, 123-138. 12 Morishima, K., Horie, S. Y. E. & Arai, K. 2002 A cryptic clonal line of the loach Misgurnus anguillicaudatus (Tele- ostei: Cobitidae) evidenced by induced gynogenesis, interspecific hybridization, microsatellite genotyping and multilocus DNA fingerprinting. Zool. Sci. 19, 565-575. (doi:10.2108/zsj.19.565) 13 Oshima, K., Morishima, K., Yamaha, E. & Arai, K. 2005 Reproductive capacity of triploid loaches obtained from Hokkaido Island, Japan. Ichthyol. Res. 52, 1-8. (doi:10.1007/s10228-004-0245-3) 14 Alves, M. J., Coelho, M. M. & Collares-Pereira, M. J. 1998 Diversity in the reproductive modes of females of the Rutilus alburnoides complex (Teleostei, Cyprinidae): a way to avoid the genetic constraints of uniparentalism. Mol. Biol. Evol. 15, 1233-1242. 15 Gunther, R., Uzzell, T. & Berger, L. 1979 Inheritance patterns in triploid Rana 'esculenta' (Amphibia, Salientia). Mitt. Zool. Mus. Berlin 55, 35-37. 16 Vinogradov, A. E., Borkin, L. J., Gunther, R. & Rosanov, J. M. 1990 Genome elimination in diploid and triploid Rana esculenta males: cytological evidence from DNA flow cytometry. Genome 33, 619-627. (doi:10.1139/g90-092) 17 Charles, J. S., Hamilton, M. L. & Petes, T. D. 2010 Meiotic chromosome segregation in triploid strains of Saccharomyces cerevisiae. Genetics 18, 537-550. (doi:10. 1534/genetics.110.121533) 18 Comai, L. 2005 The advantages and disadvantages of being polyploid. Nat. Rev. Gen. 6, 836-846. (doi:10. 1038/nrg1711) 19 Schultz, R. J. 1967 Gynogenesis and triploidy in the viviparous fish Poeciliopsis. Science 157, 1564-1567. (doi:10. 1126/science.157.3796.1564) 20 Mateos, M. & Vrijenhoek, R. C. 2005 Independent origins of allotriploidy in the fish genus Poeciliopsis. J. Hered. 96, 32-39. (doi:10.1093/jhered/esi010) 21 Monaco, P., Rasch, E. & Balsano, J. 1984 Apomictic reproduction in the Amazon molly, Poecilia formosa, and its triploid hybrids. In Evolutionary genetics of fishes (ed. B. Turner), pp. 311-318. New York, NY: Plenum Press. 22 Bogart, J., Bi, K., Fu, J., Noble, D. & Niedzwiecki, J. 2007 Unisexual salamanders (genus Ambystoma) present a new reproductive mode for eukaryotes. Genome 50, 119-136. (doi:10.1139/G06-152) 23 Avise, J. 2008 Clonality. Oxford, UK: Oxford University Press. 24 Lamatsch, D. & Stock, M. 2009 Sperm-dependent parthenogenesis and hybridogenesis in teleost fishes. In Lost sex: the evolutionary biology of parthenogenesis (eds I. Schoen, K. Martens & P. van Dijk), pp. 399-432. Heidelberg, Germany: Springer. 25 Stock, M. et al. 2002 A bisexually reproducing all- triploid vertebrate. Nat. Genet. 30, 325-328. (doi:10. 1038/ng839) 26 Stock, M., Schmid, M., Steinlein, C. & Grosse, R. 1999 Mosaicism in somatic triploid specimens of the Bufo viridis complex in the Karakoram with examination of calls, morphology and taxonomic conclusions. Ital. J. Zool. 66, 215-232. (doi:10.1080/11250009909 356259) 27 Uzzell, T. 1970 Meiotic mechanisms of naturally occurring unisexual vertebrates. Am. Nat. 104, 433-445. (doi:10.1086/282678) 28 Stock, M., Steinlein, C., Lamatsch, D. K., Schartl, M. & Schmid, M. 2005 Multiple origins of tetraploid taxa in the Eurasian Bufo viridis subgroup. Genetica 124, 255-272. 29 Gosner, L. K. 1960 A simplified table for staging anuran embryos and larvae with notes on identification. Herpetologica 16, 513-543. 30 Stock, M., Ustinova, J., Lamatsch, D. K., Schartl, M., Perrin, N. & Moritz, C. 2010 A vertebrate reproductive system involving three ploidy levels: hybrid origin of triploids in a contact zone of diploid and tetraploid Palearctic green toads (Bufo viridis subgroup). Evolution 64, 944-959. (doi:10.1111/j.1558-5646.2009.00876.x) 31 Raymond, M. & Rousset, F. 1995 GENEPOP (version 1.2): population genetics software for exact tests and ecumenicism. J. Hered. 86, 248 -249. 32 Rousset, F. 2008 GENEPOP'007: a complete reimplementation of the GENEPOP software for Windows and Linux. Mol. Ecol. Res. 8, 103-106. (doi:10.1111/j.1471-8286. 2007.01931.x) 33 Kauri, H. 1954 Uber die systematische Stellung der Europaischen Griinen Frosche Rana esculenta L. und Rana ridibunda Pall. [On the systematic position of the European green frogs Rana esculenta L. and Rana ridi- bunda Pall]. Acta Univ. Lund. N.F. 5, 1 -30. [In German.] 34 Berger, L. 1968 Morphology of the F1 generation of various crosses within Rana esculenta complex. Acta Zool. Cracow 13, 301-324. 35 Graf, J.-D. & Muller, W. P. 1979 Experimental gynogen- esis provides evidence of hybridogenetic reproduction in the Rana esculenta complex. Experientia 35, 1574-1576. (doi:10.1007/BF01953200) 36 Christiansen, D. G. & Reyer, H. U. 2009 From clonal to sexual hybrids: genetic recombination via triploids in all-hybrid populations of water frogs. Evolution 63, 1754-1768. (doi:10.1111/j.1558-5646.2009.00673.x) 37 Gunther, R. 1990 Die Wasserfrosche Europas [The water frogs of Europe]. Die Neue Brehm-Bucherei, vol. 600. [In German.] Wittenberg, Germany: Ziemsen 38 Christiansen, D. G. 2009 Gamete types, sex determination and stable equilibria of all-hybrid populations of diploid and triploid edible frogs (Pelophylax esculentus). BMC Evol. Biol. 9, 135. (doi:10.1186/1471-2148-9-135) 39 Goddard, K. A. & Schultz, R. J. 1993 Aclonal reproduction by polyploid members of the clonal hybrid species Phoxinus eos-neogaeus (Cyprinidae). Copeia 3, 650-660. (doi:10.2307/1447226) 40 Goddard, K. A., Megwinoff, O., Wessner, L. L. & Giaimo, F. 1998 Confirmation of gynogenesis in Phoxinus eos-neogaeus (Pisces: Cyprinidae). J. Hered. 89, 151-157. (doi:10.1093/jhered/89.2.151) 41 Kim, I. S. & Lee, E. H. 2000 Hybridization experiment of diploid-triploid cobitid fishes, Cobitis sinensis-longicor- pus complex (Pisces: Cobitidae). Folia Zool. 49, 17-22. 42 Alves, M. J., Coelho, M. M. & Collares-Pereira, M. J. 2001 Evolution in action through hybridisation and polyploidy in an Iberian freshwater fish: a genetic review. Genetica 111, 375-385. (doi:10.1023/A:1013 783029921) 43 Itono, M., Morishima, K., Fujimoto, T., Bando, E., Yamaha, E. & Arai, K. 2006 Premeiotic endomitosis produces diploid eggs in the natural clone loach, Misgurnus anguillicaudatus (Teleostei: Cobitidae). J. Exp. Zool. 305A, 513-523. (doi:10.1002/jez.a.283) 44 Morishima, K., Yoshikawa, H. & Arai, K. 2008 Meiotic hybridogenesis in triploid Misgurnus loach derived from a clonal lineage. Heredity 100, 581-586. (doi:10.1038/ hdy.2008.17) 45 Butlin, R., Schon, I. & Griffiths, H. I. 1998 Introduction to reproductive modes. In Sex and parthenogenesis: evolutionary ecology of reproductive modes in non-marine ostracodes (ed. K. Martens), pp. 1-24. Leiden, The Netherlands: Backhuys Publ. 46 Lutes, A. A., Neaves, W. B., Baumann, D. P., Wiegraebe, W. & Baumann, P. 2010 Sister chromosome pairing maintains heterozygosity in parthenogenetic lizards. Nature 464, 283-287. (doi:10.1038/nature08818) 47 Smith-White, S. 1948 Polarised segregation in the pollen mother cells of a stable triploid. Heredity 2, 119-129. (doi:10.1038/hdy.1948.7) 48 Werlemark, G., Uggla, M. & Nybom, H. 1999 Morphological and RAPD markers show a highly skewed distribution in a pair of reciprocal crosses between hemi- sexual dogrose species, Rosa sect. Caninae. Theor. Appl. Gen. 98, 557-563. (doi:10.1007/s001220051104) 49 Nybom, H. 2007 Unique reproduction in dogroses (Rosa sect. Caninae) maintains successful and highly heterozygous genotypes. In Apomixis (eds E. Horandl, U. Grossniklaus, P. J. van Dijk & T. F. Sharbel), pp. 281-298. Koenigstein, Germany: Koeltz Scientific Books. 50 Stock, M., Moritz, C., Hickerson, M., Frynta, D., Dujsebayeva, T., Eremchenko, V., Macey, J. R., Papenfuss, T. J. & Wake, D. B. 2006 Evolution of mito- chondrial relationships and biogeography of Palearctic green toads (Bufo viridis subgroup) with insights in their genomic plasticity. Mol. Phylogenet. Evol. 41, 663-689. (doi:10.1016/j.ympev.2006.05.026) 51 Stock, M., Lampert, K. P., Moller, D., Schlupp, I. & Schartl, M. 2010 Monophyletic origin of multiple clonal lineages in an asexual fish (Poecilia formosa). Mol. Ecol. 19, 5204-5215. (doi:10.1111/j.1365-294X.2010. 04869.x) 52 Litvinchuk, S. N. et al. 2011 Influence of environmental conditions on the distribution of Central Asian green toads with three ploidy levels. J. Zool. Syst. Evol. Res. 9, 233-239. (doi:10.1111/j.1439-0469.2010.00612.x) 53 Bell, G. 1982 The Masterpiece of nature: the evolution and genetics of sexuality. London, UK: Croom Helm. 54 Cuellar, O.1977 Партеногенез у тварин. Science 197, 837-843. (doi:10.1126/science.887925)55 Lynch, M. 1984 Дестабілізуюча гібридизація, генотипи загального призначення та географічний партеногенез. Quat. Rev. Biol. 59, 257-290. (doi:10.1086/413902)56 Kearney, M. 2003 Чому секс такий непопулярний у австралійській пустелі? Trends Ecol. Evol. 18, 605-607. (doi:10.1016/ j.tree.2003.09.021)57 Vrijenhoek, R. C. & Davis Parker Jr, D. 2009 Географічний партеногенез: генотипи загального призначення та заморожена варіація ніші. У Lost sex: the evolutionary biology of parthenogenesis (eds I. Schoen, K. Martens & P. van Dijk), с. 99-131. Гейдельберг, Німеччина: Springer.58 Pala, I., Coelho, M. M. & Schartl, M. 2008 Компенсація дози шляхом глушіння копій генів у триплоїдній гібридній рибі. Curr. Biol. 18, 1344-1348. (doi:10.1016/j. cub.2008.07.096)59 Buggs, R. J. A. et al. 2011 Транскриптомний шок генерує еволюційну новизну у новоутвореній, природній аллоплоїдній рослині. Curr. Biol. 21, 551-556. (doi:10. 1016/j.cub.2011.02.016)60 Berger, L., Uzzell, T. & Hotz, H. 1988 Визначення статі та статеві співвідношення у західних палеарктичних водяних жабах: XX та XY гібридні самки у Панононському басейні? Proc. Natl Acad. Sci. USA 140, 220-239.61 Frank, S. A. & Hurst, L. D. 1996 Мітохондрії та чоловічі захворювання. Nature 383, 224. (doi:10.1038/383224a0)62 Archetti, M. 2005 Накопичення шкідливих мутацій у геномі гібридогенетичних організмів. J. Theor. Biol. 234, 151-152. (doi:10.1016/j.jtbi.2004. 11.019)63 Muller, H. 1964 Відношення рекомбінації до мутаційного прогресу. Mut. Res. 1, 2-9.64 Felsenstein, J. 1974 Еволюційна перевага рекомбінації. Genetics 78, 737-756.65 Charlesworth, B. & Charlesworth, D. 2000 Деградація Y-хромосом. Phil. Trans. R. Soc. Lond. B 355, 1563-1572. (doi:10.1098/rstb.2000.0717)66 Guex, G. D., Hotz, H. & Semlitsch, R. D. 2002 Шкідливі алелі та диференціальна життєздатність нащадків природних геміклональних жаб. Evolution 56, 1036-1044.67 Bi, K. & Bogart, J. P. 2006 Ідентифікація інтергеномних рекомбінацій у унісексуальних саламандрах роду Ambystoma за допомогою геномної in situ hybridization. Cytogenet. Genome Res. 112, 307-312.

![рис. 1, [7‑9,21]) деякі триплоїдні хребетні поєднують клональні і статеві елементи у своєму способі розмноження, наприклад, у клептогенезі або різних формах гибридогенезу [10,22‑24], у тому числі мейотичному гибридогенезі, який вимагає репродукти](https://s3.prozo.re/batrachos/images/stock_01_YLpndq3.original.jpg)
![рис. 2, c) полягає в тому, що весь материнський внесок (NOR +, NOR—) виключається, а потім батьківський NOR + дуплікується внаслідок премейотичного ендоміозу. Мейотичні пари (NOR +/NOR +; [25]) таким чином представляють псевдобіваленти, передбача](https://s3.prozo.re/batrachos/images/stock_02_N08hPsi.original.jpg)