Студенческие работы после практики II курса - 2008 и кое-что еще
Дедух Д.В., Зарубенко Е. С. Изучение возможности определения форм зеленых лягушек по признакам внешней морфологии и окраски Деряженцева А.А., Дьяконова И.В., Микос И.Г. Изучение флуктуирующей асимметрии рисунка у Rana esculenta complex Усова Е.Е. Скелетохронологическое определение возраста предст...
Студентські роботи після практики II курсу — 2009
Студентські роботи після практики II курсу — 2010
Студентські роботи після практики II курсу — 2011
Студентські роботи після практики II курсу — 2012 (I частина)
Студентські роботи після практики II курсу — 2012 (II частина)
Студентські роботи після практики II курсу - 2013
Студентські роботи після практики II курсу - 2014
Студентські роботи після практики II курсу - 2015
Студентські роботи після практики II курсу - 2016
Студентські роботи після практики II курсу - 2017
ДОСЛІДЖЕННЯ МОЖЛИВОСТІ ВИЗНАЧЕННЯ ФОРМ ЗЕЛЕНИХ ЖАБ ЗА ОЗНАКАМИ ЗОВНІШНЬОЇ МОРФОЛОГІЇ ТА ЗАБАРВЕННЯ
Дедух Д.В., Зарубенко Є. С.
Харківський національний університет імені В. Н. Каразіна, біологічний факультет
пл. Свободи, 4, м. Харків, 61077, Україна
Rana esculenta complex — група роду зелених жаб, що складається з батьківських видів та їх геміклональних гібридів. В околицях біостанції ХНУ спільно мешкають як представники одного з батьківських видів — Rana ridibunda, так і різних (зокрема, як ди-, так і триплоїдних) гібридів — Rana kl. esculenta (Шабанов та ін., 2006).
Зібрали 73 екземпляри статевозрілих зелених жаб з заплави річки Сіверський Донець. Жаб ловили сачком у темну пору доби, збираючи біля берега та в заростях вищої водної рослинності за допомогою ліхтаря. Після виконання роботи досліджені жаби були випущені в місце вилову.
З досліджених нами жаб 15 особин мали «типові» ознаки R. ridibunda: прямий п’ятковий горбок і відсутність жовтого візерунка на стегнах. 17 особин виявилися «типовими» R. esculenta: у них був високий п’ятковий горбок і жовтий візерунок на стегнах. Решта жаб (n=41) мали комбінації ознак, характерних і для R. ridibunda, і для R. esculenta. Застосовували дискримінантний аналіз, під час якого порівняли «нетипових» особин із «типовими» R. ridibunda та R. esculenta за ознаками, що використовуються різними авторами для розрізнення форм зелених жаб. Це відношення довжини тіла до довжини гомілки, відношення довжини п’яткового горбка до довжини гомілки, колір спинної сторони тіла (темно-зелений, зелений, світло-зелений), форма плям (менша за око, більша за око), частота плям (більша за фон, однакова кількість з фоном, менша за фон).
У результаті дискримінантного аналізу ознаки «нетипових» жаб утворили хмару розсіювання, всередині якої розташовувалися ознаки «типових» R. ridibunda та R. esculenta. Це означає, що, користуючись даним набором ознак, неможливо визначити приналежність до певної форми всіх жаб із дослідженого місця проживання.
Дана робота виконана як УІРС у рамках навчально-польової практики з зоології хребетних. Автори висловлюють подяку керівникам роботи ст. викл. Кравченку М.А. та доц. Шабанову Д.А., а також дякують А.А. Атемасову за цінну консультацію.
Дедух Д.В., Зарубенко Є.С. (наукові керівники – Кравченко М.А., Шабанов Д.А.) Дослідження можливості визначення форм зелених жаб за ознаками зовнішньої морфології та забарвлення// «Біологія: від молекули до біосфери». Матеріали III Міжнародної конференції молодих науковців. - Харків: СПД ФО Михайлов Г.Г., 2008. - с. 364-365.
ДОСЛІДЖЕННЯ ФЛУКТУЮЧОЇ АСИМЕТРІЇ МАЛЮНКА У RANA ESCULENTA COMPLEX
Деряженцева А.А., Дьяконова І.В., Микос І.Г.
Харківський національний університет імені В.Н. Каразіна
пл. Свободи, 4, м. Харків, 61077, Україна
e-mail: bell-iren@mail.ru
Одним із заходів оцінки стійкості розвитку є флуктуюча асиметрія (ФА) (Гелашвілі Д.Б. та ін., 2004). У Rana esculenta complex зареєстровано міжвидову гібридизацію, клональне успадкування, природну поліплоїдію (Plötner, 2005; Шабанов та ін., 2006). Це робить цікавим порівняння двох різних форм жаб з одного місця проживання за характерними для них рівнями ФА.
Досліджено 149 зелених жаб, що належать до Rana esculenta complex. Усі особини були спіймані в річці Сіверський Донець в околицях біостанції ХНУ імені Каразіна (с. Гайдари Зміївського р-ну Харківської області). На підставі морфофізіологічних характеристик жаби розподілялися за тією чи іншою формою. З досліджених екземплярів 116 мали типові для форми Rana ridibunda ознаки: низький і косий п’ятковий горбок, відсутність жовтого візерунка на стегнах, темні резонатори у самців; 24 особини були віднесені до форми Rana esculenta на підставі таких критеріїв: високий п’ятковий горб, жовтий колір на задніх кінцівках, світло-сіре забарвлення резонаторів у самців. 9 особин ми не змогли впевнено віднести до жодної з форм, що, можливо, пов’язано з їхньою поліплоїдністю (морфофізіологічні ознаки формувалися під впливом одразу декількох геномів).
Для визначення характеру асиметрії ми реєстрували такі ознаки: характер дорзомедіальної смуги (ДМП) (симетрична, злегка асиметрична, сильно асиметрична), кількість плям-смуг на правій і лівій нижніх кінцівках від стегна до кінця гомілки, а також кількість плям уздовж ДМП зліва і справа. Щоб визначити, чи можуть ці ознаки використовуватися як міра ФА, ми провели порівняння груп за цими критеріями. Виявилося, що мірою ФА може бути абсолютне («за модулем») значення різниці між кількістю плям зліва і справа вздовж ДМП, яке змінювалося від 0 до 3.
Середнє значення ФА за цією ознакою (тобто різниця між кількістю плям уздовж ДМП праворуч і ліворуч) для R. esculenta становило 0,87, а для R. ridibunda — 0,67 (при порівнянні за критерієм Стьюдента р = 0,18). Таким чином, відмінність гібридів та особин батьківської форми є недостовірною (можливо через недостатній обсяг вибірки), однак, незважаючи на отримані показники, було помічено тенденцію до більшої асиметрії гібридних особин.
Було з'ясовано, що у самців Rana esculenta complex асиметрія виражена сильніше, що, імовірно, обумовлено слабшою зарегульованістю їхнього розвитку порівняно із самками.
Таким чином, ми припускаємо, що кількість плям уздовж ДМП може бути використана для оцінки ФА різних (ді- та триплоїдних) гібридів.
Дана робота виконана в рамках УІРС навчально-польової практики з зоології хребетних під керівництвом старшого викладача Кравченка М.А. та доцента Шабанова Д.А.
Деряженцева А.А., Дьяконова І.В., Микос І.Г. (наукові керівники – Кравченко М.А., Шабанов Д. А.) Вивчення флуктуючої асиметрії малюнка у Rana esculenta complex // «Біологія: від молекули до біосфери». Матеріали III Міжнародної конференції молодих науковців. - Харків: СПД ФО Михайлов Г.Г., 2008. - с. 365-366.
СКЕЛЕТОХРОНОЛОГІЧНЕ ВИЗНАЧЕННЯ ВІКУ ПРЕДСТАВНИКІВ ЗЕЛЕНИХ ЖАБ RANA ESCULENTA COMPLEX З ХАРКІВСЬКОЇ ОБЛАСТІ
Усова Є.Є.
Харківський національний університет імені В.Н. Каразіна, кафедра зоології та екології тварин, пл. Свободи, 4, м. Харків, 61077, Україна
e-mail: е_usova@mail.ru
Група зелених жаб — Rana esculenta complex — одна з найцікавіших для вивчення груп тварин Харківської області. До неї входять два батьківські види, а також різні ди-, три- і навіть тетраплоїдні форми їх гібридів (Plotner, 2005). Співіснування гібридів з батьківськими видами підтримується завдяки геміклональному успадкуванню (передачі гібридами певних геномів клонально, без рекомбінації), що представляє значний науковий інтерес. При спільному проживанні ди- та триплоїдних гібридів виникає можливість порівняння їх життєздатності. Адекватним методом для цього є скелетохронологічне визначення віку та швидкості росту.
Нами вивчено 62 особини зелених жаб, що зберігаються в колекції ЗІН РАН (Санкт-Петербург), Музею природи ХНУ та кафедри зоології й екології тварин ХНУ. Вивчено 50 особин Rana esculenta (гібридів); 9 — Rana ridibunda та 3 — Rana lessonae. Вилов жаб проводився в Харківській області з 2004 по 2007 р. Видова приналежність і склад геномів у генотипі для більшості досліджених особин визначалися методом проточної ДНК-цитометрії в ІНЦ РАН (Санкт-Петербург) С.Н. Литвинчуком та Ю.М. Розановим. Частина особин (представники R. lessonae) визначені морфологічно за комплексом ознак (Лада, 1995; Шабанов та ін., 2006).
Для визначення віку були використані зрізи декальцинованих фаланг пальців, які потім фарбували гематоксиліном Ерліха (Ромейс, 1953). Зрізи були отримані за допомогою заморожувального мікротома. Визначення віку ми проводили шляхом підрахунку кількості ліній зрощення в періостальній зоні кістки з урахуванням можливої резорбції спочатку відкладених шарів. Як правило, перша лінія зрощення резорбується повністю, а друга — частково. Вік особин можна обчислювати за формулою: вік = кількість повністю видимих (не уражених резорбцією) ліній зрощення + 2.
Ми встановили, що в Харківській області вік статевозрілих зелених жаб, спійманих на нересті, знаходиться в межах від 4-х до 10-ти років, з переважанням 5-ти та 6-ти річних особин. У досліджених гібридних особин зареєстровано істотне різноманіття за темпами їхнього росту. Найбільший вік мають не найбільші особини, а найбільші особини не є найстарішими. Наприклад, зареєстровано особин з довжиною тіла понад 740 мм у віці 5 років та 8–10-річних з довжиною тіла, що не перевищує 689 мм. Нами не було зареєстровано істотних відмінностей у швидкості росту між диплоїдними та триплоїдними гібридами, що є попереднім свідченням нормальної життєздатності триплоїдних гібридів.
У ході подальших досліджень ми плануємо збільшити кількість досліджених жаб та визначити динаміку росту окремих особин. При вивченні жаб з одного місця проживання (пойми Сіверського Донця в околицях біостанції ХНУ) порівняння швидкості росту та тривалості життя представників різних форм дасть можливість оцінити їх життєздатність та внесок у відтворення геміклональної популяційної системи.
Робота виконана під керівництвом доцента кафедри зоології та екології тварин ХНУ, к. б. н. Шабанова Д.А. Автор висловлює подяку за допомогу в роботі Шабановій А.В., Коршунову А.В. та Шабанову Д.А.
Усова Є.Є. (науковий керівник – Шабанов Д. А.) Скелетохронологічне визначення віку представників зелених жаб Rana esculenta complex з Харківської області// «Біологія: від молекули до біосфери». Матеріали III Міжнародної конференції молодих науковців. - Харків: СПД ФО Михайлов Г.Г., 2008. - с. 396-397.
ОСОБЛИВОСТІ СТРУКТУРИ ФОЛІДОЗУ ГОЛОВИ У СТРИБУЧИХ ЯЩІРОК (LACERTA AGILIS) ОКОЛИЦЬ БІОСТАНЦІЇ ХНУ
Садовнича М. А., Чусова О. А.
Харківський національний університет імені В. Н. Каразіна, біологічний факультет
пл. Свободи, 4, м. Харків, 61077, Україна
Визначення особливостей фолідозу (щиткоутворення) широко застосовується в систематиці рептилій та аналізі їх популяційної структури. При цьому враховується кількість, форма та взаєморозташування окремих щитків голови та тіла.
У роботі розглянуто особливості щиткування лобової частини голови 16 особин спритної ящірки (Lacerta agilis), зібраних у НПП «Гомолишанські ліси» (Зміївський р-н, Харківська обл.) та в околицях біостанції ХНУ (Зміївський р-н, с. Гайдари). При описі фолідозу голови спритних ящірок за основу взято зведену таблицю різноманітності морфологічних ознак передньоносових (пн), лобоносових (лн) лобноносового (лн) та інших щитків, складена за результатами дослідження географічної мінливості спритних ящірок у межах колишнього СРСР (Яблоков, Ларіна, 1985).
Під час обстеження зібраних екземплярів відзначено ознаки, не включені до зазначеного зведення. Перший з них — варіант будови передньоносових щитків, при якому між ними спереду відносно глибоко вклинюється міжносовий (мн) щиток (рис. 1). Такий стан спостерігався у 5 із 16 досліджених нами особин.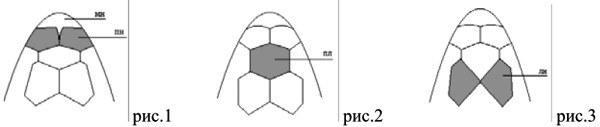
Другою зареєстрованою ознакою, яка не входила до опублікованої схеми опису, є варіант будови лобно-носового щитка, за якого він має форму правильного шестикутника. Звернені передньолатерально та задньолатерально вершини цього щитка з'єднуються з передніми вершинами передлобних щитків під кутом приблизно 110° (рис. 2). Ця особливість є найбільш поширеним варіантом будови лобноносового щитка для вивченої вибірки ящірок і спостерігалася у 15 особин із 16 вивчених.
Нарешті, відзначено новий варіант будови передлобових щитків, при якому вони мають ромбоподібну форму і стикаються лише в одній точці у своїй задній частині (рис. 3). Ця особливість фолідозу спостерігалася в однієї з розглянутих нами особин.
У ході роботи не було зареєстровано зв'язку між забарвленням тварин та особливостями їхнього щиткування.
Ймовірно, наявність «незвичайних» морфологічних ознак фолідозу голови характеризує особливості розглянутої нами вибірки ящірок. Надалі ці ознаки можуть бути використані для фенетичного аналізу популяцій спритних ящірок.
Робота виконана на біологічній станції ХНУ ім. В.Н.Каразіна під керівництвом ст. викл. Кравченко М.А. та доц. Шабанова Д.А., яким ми висловлюємо щиру подяку.
Садовничая М. А., Чусова О. А. Особливості структури фолідозу голови у спритних ящірок (Lacerta agilis) околиць біостанції ХНУ// «Біологія: від молекули до біосфери». Матеріали III Міжнародної конференції молодих науковців. - Харків: СПД ФО Михайлов Г.Г., 2008. - с. 398-399.
ЩОДО РОЗРОБКИ МЕТОДІВ ФОРМАЛІЗОВАНОЇ ОЦІНКИ ДОЦІЛЬНОСТІ ОХОРОНИ ПРИРОДНИХ ОБ’ЄКТІВ
Кравченко М. А.
Харківський національний університет імені В.Н. Каразіна, біологічний факультет, кафедра зоології та екології тварин
пл. Свободи, 4, м. Харків, 61077, Україна
e-mail: m_kravchenko@inbox.ru
В умовах швидких змін біосфери, спричинених як антропогенними впливами, так, можливо, й іншими причинами, актуальною стає проблема вибору природних об’єктів, пріоритетних з точки зору їх охорони. У даній роботі ми розглянемо деякі принципи, що визначають підстави для охорони природних об’єктів (популяцій, видів, біогеоценозів тощо).
Будь-який об’єкт можна охороняти як заради його корисності, так і з етичних міркувань, з морального обов’язку. Оцінкою корисності об’єкта може бути його вартість, а оцінкою етичних підстав для його охорони – його цінність. Наприклад, концепція раціонального використання природних ресурсів оцінює потенційні об’єкти охорони з точки зору їхньої корисності (актуальної або потенційної) (Марушевський, 2008), а т.зв. «екологічна етика» та інші натуроцентричні концепції розглядають насамперед етичні підстави для охорони природи (Рьюз, 1992; Борейко, 2004). На нашу думку, при виборі об’єктів для охорони необхідно враховувати як їхню вартість, так і їхню етичну цінність. Концепція, на основі якої має здійснюватися такий вибір, повинна дозволяти хоча б якісно оцінювати ці параметри.
Етичні підстави для охорони об’єктів пов’язані з їхньою унікальністю та можливістю їхнього існування в довгостроковій перспективі, потенційним безсмертям (Кравченко, Шабанов, 2006). Відповідно до сказаного, ми пропонуємо такий спосіб порівняння підстав для охорони тих чи інших об’єктів
Сама необхідність охорони певного об’єкта є наслідком тієї чи іншої небезпеки, що загрожує його існуванню. У переважній більшості випадків охорона не може повністю усунути загрозу знищення об’єкта, а лише зменшує її ймовірність. Ефективність охорони можна оцінити як зниження ймовірності знищення об’єкта завдяки охоронним заходам.
Оцінити унікальність об'єкта можна за ймовірністю виникнення його аналога, ідентичного даному об'єкту за істотними для нас властивостями. Мірою потенційного безсмертя об'єкта може бути ймовірний термін його існування в тому випадку, якщо він збережеться в короткостроковій перспективі (тій, на яку плануються заходи щодо його охорони).
Викладений підхід поєднує економічні підстави для охорони об’єктів із формалізованою оцінкою їх етичної цінності. Крім двох економічних оцінок (вартості об’єкта, тобто можливої вигоди, отриманої від його існування, а також вартості заходів щодо його охорони), що вимагають залучення економістів, у наведеній формулі використовуються ймовірнісні величини, пов’язані з особливостями виникнення та загибелі об’єктів, що охороняються. Оцінка таких ймовірностей має бути завданням фахівців з вивчення даної категорії систем. Вона може здійснюватися як на підставі статистичної обробки емпіричних даних, що описують долю аналогічних об’єктів, так і на підставі результатів моделювання відповідних природних систем.
Автор висловлює подяку доценту, к.б.н. Д.А. Шабанову, під керівництвом якого виконано дану роботу.
Кравченко М. А. До розробки методів формалізованої оцінки доцільності охорони природних об’єктів // «Біологія: від молекули до біосфери». Матеріали III Міжнародної конференції молодих науковців. - Харків: СПД ФО Михайлов Г.Г., 2008. - с. 442-443.
ЩО НОВОГО МОЖНА ДІЗНАТИСЯ ПРО ЗЕЛЕНИХ ЖАБ ЗА ДОПОМОГОЮ ІМІТАЦІЙНОГО МОДЕЛЮВАННЯ?
1 Кравченко М.А., 2 Луцик А.А., 1 Шабанов Д.А.
Харківський національний університет імені В.Н. Каразіна,
1 біологічний факультет, кафедра зоології та екології тварин
2 механіко-математичний факультет, кафедра вищої математики та інформатики
e-mail: 1 m_kravchenko@inbox.ru; d.a.shabanov@gmail.com, 2 aalutsyk@gmail.com
Описуючи різноманіття живого світу, ми спрощуємо різноманіття спостережуваних явищ до набору типових схем. Так, прийнято вважати, що організми в природі належать до певних видів і входять до складу популяцій, що складаються з особин цього виду. Переважна більшість роздільностатевих диплоїдних організмів передають своїм нащадкам геноми, що є результатом рекомбінації тих двох геномів, які кожен з цих організмів отримав від батьків. Однак серед описаних схем існує чимало винятків. Одне з них — гібридизація з утворенням геміклональних гібридів, що передають нащадкам геноми одного з двох батьківських видів у чистоті. Зокрема, така гібридизація характерна для зелених жаб.
До групи європейських зелених жаб належать два батьківські види: ставкова, Pelophylax lessonae (Camerano, 1882) та озерна, Pelophylax ridibundus (Pallas, 1771) жаби, а також їхні гібриди, названі їстівними жабами, Pelophylax esculentus (Linnaeus, 1758). Назви цих форм надані відповідно до результатів останньої ревізії системи амфібій (Frost et al., 2006 та більш ранні роботи). Більш відомі їхні традиційні назви — Rana lessonae Camerano, 1882, Rana ridibunda Pallas, 1771 та Rana esculenta Linnaeus, 1758. Чому гібридна форма носить ім'я, подібне до видового? Гібридні жаби можуть тривалий час існувати без повторних схрещувань батьківських видів і утворювати популяції, в яких вони успішно розмножуються протягом багатьох поколінь.
Описуючи розмноження зелених жаб, ми позначаємо геном P. lessonae (що складається з 13 хромосом) символом L, а геном P. ridibundus (також 13 хромосом) – R. Геміклональне успадкування у P. esculentus виражається в тому, що тільки один із геномів диплоїдних гібридних жаб переходить у гамети. Такий геном називається клональним; ми пропонуємо позначати цю обставину, беручи символ відповідного геному в дужки. Другий геном у диплоїдних гібридів елімінується на тій чи іншій стадії гаметогенезу. Наприклад, у багатьох місцевостях Східної України спільно мешкають і розмножуються P. ridibundus та P. esculentus. Такі та подібні до них сукупності зелених жаб некоректно називати «популяціями», оскільки їх складають тварини, що не належать до одного виду. Ми пропонуємо називати такі біосистеми геміклональними популяційними системами – ГПС. Особини батьківського виду виробляють гамети з рекомбінантними геномами цього виду, а гібриди – лише гамети з клональним геномом (L). У результаті все потомство від схрещування батьківського виду та гібридів з клональним геномом іншого виду виявляється гібридним: RR × (L)R → (L)R. Клональний геном при цьому передається з покоління в покоління без рекомбінації.
Часто ГПС має набагато складніший склад. В околицях біостанції ХНУ (с. Гайдари Зміївського р-ну Харківської обл.) мешкає ГПС, до складу якої входять жаби RR, (L)R, L(R), (L)(R), LLR, LRR, а серед ювенільних особин зустрічаються також LL і LLRR. У три- та тетраплоїдних гібридів клональні геноми в дужки не взяті, оскільки їх гаметогенез повністю не вивчений; особини, позначені як (L)(R), одночасно виробляють гамети, що несуть геноми P. lessonae та P. ridibundus у чистоті.
ГПС – відносно нова категорія біосистем для сучасної науки. Процеси, що підтримують їхнє відтворення, залишаються значною мірою невивченими. Для їх вивчення ми застосували імітаційне моделювання з використанням мультиагентної комп'ютерної моделі. Одним з авторів даного повідомлення (А.А. Луциком) під керівництвом доц. М.В. Володимирової та проф. Г.Н. Жолткевича була створена програма Batrahometrics, що моделює трансформації ГПС зелених жаб. За допомогою цієї моделі можна описати початковий стан ГПС (що включає чисельність особин певних форм і вікових груп); вказати правила, за якими стан ГПС у році n визначає його ж у році n+1; визначити розподіл ймовірностей стану, в якому ця ГПС опиниться через певний проміжок часу.
До числа спрощень, прийнятих при побудові моделі, належить постійність чисельності нерестового стада (1000 особин) та ідентичність властивостей особин, що належать до певної форми (визначеної набором її геномів). Залежно від набору геномів особина характеризується певною привабливістю для партнерів, плодючістю, виживаністю до настання статевої зрілості та після дозрівання. На кожному кроці роботи моделі, що відповідає одному року, відбувається ймовірнісне утворення пар та ймовірнісна загибель наявних особин. Відстежуючи передачу клональних та рекомбінантних геномів з покоління в покоління, програма Batrahometrics відстежує зміну складу ГПС у часі.
На вході кожного окремого експерименту (моделювання трансформацій певної ГПС протягом певного часу) враховується відносно невелика кількість параметрів (чисельність, вік і статевий склад жаб окремих форм, а також, за необхідності, динаміка їхнього прибуття до модельної популяційної системи). При моделюванні змін програма враховує зміни протягом кожного «року» (кроку роботи моделі) 16 000 незалежних параметрів, пов'язаних з окремими особинами в модельному ГПС. Результатом експерименту є та чи інша послідовність трансформацій складу ГПС, а також відносно невелика кількість основних параметрів, що описують досягнутий нею стан.
Отже, на вході та на виході моделі використовується невелика кількість параметрів, а на кожному кроці аналізується набагато більша їх кількість. Це пов'язано з тим, що в щорічних змінах ГПС є як закономірні (детерміновані), так і випадкові (стохастичні) компоненти. Носій того чи іншого генотипу може розмножитися і змінити всю популяційну систему, а може і просто загинути. Відстеження «долі» кожної окремої особини дає можливість моделювати вплив випадковості на трансформації ГПС. Провівши експеримент з однаковими початковими умовами достатню кількість разів, можна визначити розподіл ймовірностей кінцевого стану.
Навіщо потрібні такі експерименти? Їхнє головне призначення — не визначення долі конкретних ГПС, а перевірка адекватності наших уявлень про системи такого роду. Ми дізнаємося про певні особливості об’єктів нашого дослідження і будуємо систему своїх уявлень про них. Щоб побудувати модель, таку систему треба формалізувати (що саме по собі є вельми корисною роботою, яка виявляє можливі порушення логіки в загальноприйнятих поглядах). Модель будується на основі системи уявлень про об’єкт і дає можливість порівняти різноманіття об’єктів, що реєструється в природі, з різноманіттям станів моделі, що реєструється в експерименті. Якщо набір станів, у які може переходити ГПС у моделі, відрізняється від набору станів, що спостерігаються у природних об’єктах, то уявлення, покладені в основу моделі, потребують зміни. Відповідність моделі об’єкту не доводить правильність системи уявлень про об’єкт, але слугує аргументом на її користь.
В описаному використанні моделі проявляється можливість переходу від редукціоністського, поелементного опису властивостей досліджуваного об’єкта до вивчення його характеристик як цілого (визначення сукупності та розподілу ймовірностей шляхів його розвитку).
На думку авторів, подібний підхід може бути застосований і при дослідженні інших біологічних систем різних рівнів організації.
Автори дякують М.В. Володимирову та Г.Н. Жолткевичу за неоціненну допомогу в постановці завдання даної роботи та її виконанні.
Кравченко М.А., Луцик А.А., Шабанов Д.А. Що нового можна дізнатися про зелених жаб за допомогою імітаційного моделювання? (Пленарна доповідь на відкритті конференції) // «Біологія: від молекули до біосфери». Матеріали III Міжнародної конференції молодих науковців. - Харків: СПД ФО Михайлов Г.Г., 2008. - с. 9-11.