Альтруизм и парадокс Симпсона. Колонка в КомпьютерреOnline #50
В разных условиях наибольший успех обеспечивают разные формы поведения – иногда чисто эгоистические, иногда – антагонистические, но довольно часто – и альтруистические.
У суперечках з приводу моїх колонок хтось із борців з наукою стверджував, що для еволюційної біології незрозуміла альтруїстична поведінка людей. Ця добра людина не знала, що вивчення еволюції альтруїзму – одна з галузей еволюційної психології, що швидко розвиваються. Багато цікавого з цього приводу написано Олександром Марковим. Хороший огляд цієї теми міститься в його нещодавньому двотомнику про еволюцію людини. Не переказуючи те, що можна прочитати в Маркова, я все ж присвячу пару колонок цій темі.
Ще за часів Дарвіна з'явилося хибне трактування дарвінізму як війни всіх проти всіх. З числа тих, хто вважав, що Дарвін недооцінював взаємодопомогу, найяскравішим був князь Петро Олексійович Кропоткін, географ і теоретик анархізму. Кропоткінська «Взаємодопомога як фактор еволюції», написана в 1902 р., не містить альтернативи чи спростування дарвінізму, вона лише інакше розставляє акценти, приділяючи більшу увагу гармонії у відносинах між особинами. З Кропоткіним не можна не погодитися в тому, що різні форми взаємодопомоги широко поширені в нашому світі. У ході еволюції вони виникали неодноразово.
Чи заперечують дарвінізм і пізніша еволюційна біологія взаємодопомогу? Звичайно, ні. Вони розглядають виникнення пристосувальних ознак, у тому числі, і поведінкових. У різних умовах найбільший успіх забезпечують різні форми поведінки – іноді суто егоїстичні, іноді – антагоністичні, але досить часто – і альтруїстичні.
Як альтруїстична поведінка могла закріпитися в ході еволюції? Наприкінці XIX-початку XX століття її пояснювали добором між групами (видами, популяціями, стадами та ін.) особин. Здавалося природним, що еволюція мала підтримувати об'єднані взаємодопомогою групи.
З поширенням СТЕ пояснювати становлення альтруїзму стало складніше. Еволюцію поведінки стали уявляти як поширення генів, що викликають певні форми поведінки. Чи може поширитися ген, який сприяє поведінці, що зменшує частку носіїв цього гена в наступних поколіннях? Ні! А хіба альтруїзм (принесення своїх інтересів у жертву інтересам родичів) не призведе до такого ефекту? Деякі найбільш завзяті прихильники СТЕ вирішили, що можливість групового добору спростована в принципі. Вони часто посилаються на Річарда Докінза. На щастя, Докінз не заперечує груповий добір як такий. У «Розширеному фенотипі» він лише стверджує, що груповий добір суттєво менш ефективний, ніж добір особин.
Одразу скажу, що існування групового добору доведено. Для прикладу пошлюся на зроблений Марковим переказ дослідження, у якому було доведено наявність добору видів. Пасльонові неодноразово переходили від перехресного запилення до самозапилення. У фрагментах еволюційного дерева пасльонових, де практикувалося самозапилення, і виникнення нових видів, і їхнє вимирання йшло швидше, ніж у їхніх перехреснозапильних родичів. Проте, те, наскільки груповий добір поширений і ефективний - предмет суперечок. Чи можна пояснити альтруїзм без групового добору? Щонайменше дві форми альтруїзму цілком зрозумілі на рівні індивідуального добору.
Споріднений альтруїзм. Чи може еволюція підтримати поведінку самки, за якої вона жертвуватиме життям заради виводка своїх нащадків? Цілком. Прихильники СТЕ пояснюють це так. Уявіть собі ген, що викликає готовність до жертовної поведінки матері. Якщо вона є носієм такого гена, він з високою (і обчислюваною) ймовірністю виявляється притаманний і її нащадкам. Жертвуючи собою заради потомства, така особина за певних умов поширюватиме ген готовності до самопожертви.
Якщо як слід пошкребти реципрокний (взаємний) альтруїзм, з'ясується, що він, насправді, не альтруїзм, а розважливий егоїзм. Ідеться про відносини «ти – мені, я – тобі». У популяціях, де особини здатні впізнавати одна одну індивідуально, можливі відносини, за яких особина, приміром, ділиться їжею з родичем, заробляючи можливість отримати частку від здобичі сусіда, коли той виявиться більш удачливим. Індивідуальний добір сприятиме поширенню в популяції генів, що зумовлюють здатність допомагати ближнім, запам'ятовувати їхню репутацію і вимагати повернення «боргу» за можливості.
Чи можна пояснити всі прояви альтруїзму цими механізмами? Навряд чи. Що робить нормальний чоловік, побачивши чужу дитину в небезпеці? Кидається на допомогу, незважаючи на небезпеку для себе. Очевидно, така форма поведінки є вродженою, інстинктивно визначеною. (Репліка вбік: деякі читачі відчувають обурення від твердження, що людина – тварина, і багато особливостей її поведінки зумовлені еволюційно. Їм чомусь здається, що така поведінка обов'язково має бути «скотською», аморальною. На щастя, багато наших «тваринно» визначених форм поведінки непогано узгоджуються з вимогами моралі). Здається, для пояснення такого альтруїзму потрібен груповий добір.
Тут є одна проблема. Мені здається, складніше пояснити не те, чому виникають групи, що складаються з альтруїстів, а те, чому ці групи не руйнуються егоїстами. Уявіть собі: у групі, де всі особини допомагають одна одній, перевагу може отримати та, яка прийматиме допомогу, не надаючи її взамін. Такі особини можуть залишати більше нащадків, ніж альтруїсти, і витісняти їх з часом! Чому ж альтруїсти в нашому світі не перевелися зовсім?
Одна з відповідей на це питання пов'язана з парадоксом Сімпсона. Я дізнався про нього зі статті Маркова; відсилаю і вас до неї. Це обговорення статті американських авторів, що створили експериментальну модель для демонстрації цього парадоксу. Він може проявлятися в популяції, яка то поділяється на субпопуляції, на розвиток кожної з яких впливає її склад, то возз'єднується наново. Ось ілюстрація зі згаданої статті.
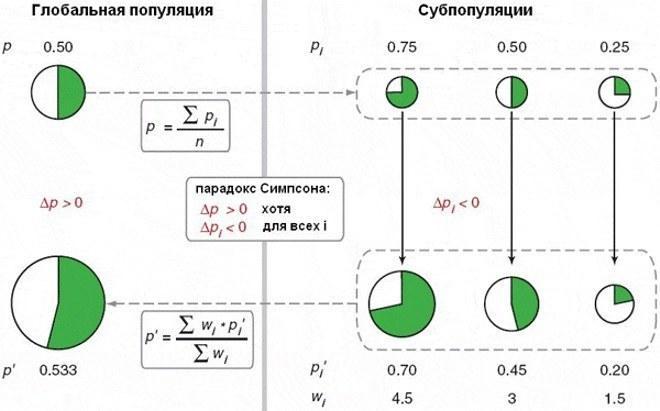
Парадокс Сімпсона: у кожній з трьох субпопуляцій частка альтруїстів (показана зеленими секторами) скорочується, а в цілому вона зростає. Причина – у прискореному зростанні тих субпопуляцій, де вища частка альтруїстів
Автори статті, яку переказує Марков, показали дію парадоксу Сімпсона на культурах бактерій. Вони вирощували суміш із двох штамів. Одні (альтруїсти) виробляли речовину, що вмикає механізм захисту від антибіотика, інші (егоїсти) – ні. При спільному «егоїсти» користувалися речовиною, виробленою "альтруїстами". Колонії, де було багато альтруїстів, росли швидше. Хоча в кожній з них частка альтруїстів скорочувалася, у цілому для популяції вона зростала. Красиво, правда?
Щоб дати студентам можливість «помацати» парадокс Сімпсона, я зробив модель, що його ілюструє, в Excel. Завантажити її можна тут; на жаль (можливо – через мою програмістську недосвідченість) вона виявилася досить громіздкою.
Як ви зрозуміли, парадокс Сімпсона проявляється, коли популяція то поділяється на субпопуляції, то об'єднується знову. Я вирішив почати з якогось простого випадку поділу-об'єднання, і згадав про життєвий цикл багатьох донних морських тварин – від коралових поліпів до асцидій.
Уявіть собі тварин, що утворюють групи на придатних для них ділянках дна. У належний строк вони викидають у воду статеві клітини (гамети). У товщі води розвиваються личинки, що осідають на дно. Шанси кожної особини на виживання залежать не лише від її спадкових задатків, а й від її найближчого оточення (а властивості цього оточення залежать від спадковості особин, що утворюють групу). Якраз те, що потрібно для перевірки роботи парадоксу Сімпсона!
Групи таких донних тварин можуть страждати від хижаків. Приміром, при нападі морської зірки якісь із наших тварин можуть егоїстично ховатися в ґрунт, а якісь – альтруїстично (з точки зору родичів) нападати на агресора. Альтруїсти частіше гинутимуть, але зате хижаки віддаватимуть перевагу групам, де альтруїстів небагато. Чи вистачить цих передумов, щоб парадокс Сімпсона підтримав гени альтруїзму?
Забігаючи наперед, скажу, що в міру налагодження моделі я зрозумів, що відмінностей у смертності в різних групах недостатньо для підтримки альтруїстів. У моделі виникають субпопуляції, чисельність яких визначається випадково, від 0 до якоїсь змінної величини. Підвищена частка альтруїстів частіше досягається в дрібних групах. Після того, як смертність скоротить чисельність усіх груп, внесок «щасливчиків» у загальний результат стає малосуттєвим. Мені довелося модифікувати модель, додавши до неї розмноження всередині субпопуляцій. Це реалістичне припущення. Донні тварини, про яких ми говорили, від коралів до асцидій, крім статевого розмноження часто використовують і нестатеве. У хороших умовах удачлива особина брунькується, утворюючи групу своїх генетичних копій. До часу статевого розмноження гамети у воду випустить не одна особина, а кілька.
Якщо хочете – завантажте файл Excel і пограйтеся з моделлю самі. Тут я розповім лише про кілька зроблених з її допомогою висновків.
У моделі незалежно задається вплив алелів гена A на шанси особини вижити і розмножитися всередині тієї групи, до якої вона потрапила, і рівень смертності та кількості нащадків, характерних для груп у цілому. Якщо алель A забезпечує альтруїстичну поведінку, його носії мають програвати своїм конкурентам на індивідуальному рівні. Такий результат показаний на малюнку. Стрибки на графіку пов'язані з високою часткою випадковості при формуванні субпопуляцій, загибелі і розмноженні особин. При іншому запуску моделі траєкторія виявиться дещо іншою, але кінцевий результат (викорінення гена альтруїзму) найімовірніше залишиться тим самим.
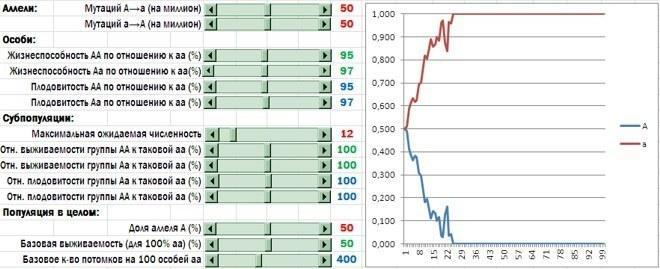
Носії алеля A програють носіям алеля a. Цей ефект сильніше виражений у генотипу AA, слабше - у Aa (інакше кажучи, має місце кодомінування). Спочатку частки алелів A і a рівні, але протягом перших 25 років добір видаляє алель A з популяції
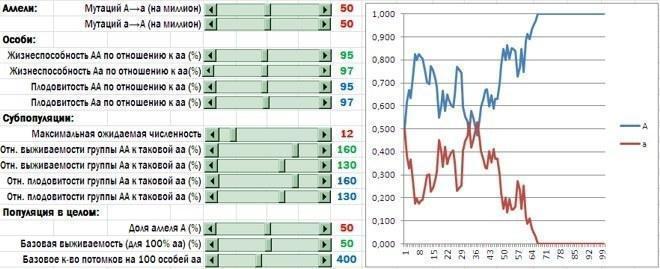
Субпопуляції з високою часткою альтруїстів (носіїв алеля A) отримали перевагу у виживаності і плодючості. Тепер найчастіше в популяції закріплюється алель альтруїзму (хоча на результат конкуренції алелів сильно впливає випадковість)
Що в підсумку? Перший висновок я вже назвав. Груповий добір на користь альтруїстів працює набагато ефективніше, коли він проявляється не лише в меншій смертності груп, що складаються з альтруїстів, а й у їхній більшій плодючості.
Другий висновок полягає в тому, що груповий добір пересилює добір на рівні особин лише тоді, коли він пов'язаний з вагомими відмінностями між групами. Якщо особини-альтруїсти поступаються егоїстам на одиниці відсотків від рівня їхньої виживаності і плодючості, то парадокс Сімпсона стійко проявлятиметься при перевагах груп альтруїстів у десятки відсотків.
Третє (картинками не ілюструю; хочете – перевірте). Альтруїсти частіше перемагають, ніж програють, тільки якщо субпопуляції виявляються маленькими. Якщо групи будуть досить великими, і при їх утворенні не діятиме жодний механізм, що збирає альтруїстів разом, частка альтруїстів у всіх групах буде приблизно однакова. Переважить ефект добору проти альтруїстів усередині субпопуляцій. На жаль, оскільки групи, у яких може проявитися парадокс Сімпсона, невеликі й ефемерні, на їхню еволюцію серйозний вплив справляє проста випадковість.
Отже, одним парадоксом Сімпсона закріплення альтруїзму в еволюції не поясниш. Але чи всі чинники ми врахували?