Мы используем аналитические cookie только чтобы считать посетителей и оценивать нагрузку — и никогда не продаём ваши данные. Они загружаются только с вашего согласия.
Privacy policy
Подпорки альтруизма. Колонка в КомпьютерреOnline #51
Характерной особенностью альтруистичного поведения многих животных, включая человека, является то, что оно касается только особей, входящих в определенные совокупности.
Дмитро Шабанов
Останню колонку я закінчив тим, що парадокса Сімпсона самого по собі для розвитку істинного альтруїзму недостатньо. Він може привести до збільшення числа носіїв генів альтруїзму лише за досить специфічних умов. Утім, у самому кінці колонки я допустив наявність якихось ще факторів, що підтримують добір на користь альтруїзму.
Я обговорю такі фактори тут. Але до переходу до основного змісту сьогоднішньої колонки застережу читачів від однієї нерідко зустрічуваної помилки. Жанр колонки очевидно обмежений за обсягом. Посилаючись на якісь обставини, я намагаюся вставити в текст мінімальні пояснення, необхідні для їх розуміння. Це означає, що повна картина якоїсь непростої проблеми в колонку не вміститься, я змушений обмежуватися обговоренням лише одного чи кількох її цікавих аспектів. Не поспішайте робити висновок про те, що якщо я не згадав якусь із проблем чи не послався на якийсь феномен, то я заперечую їх існування чи важливість, гаразд? Наприклад, якщо я обговорюю специфіку добору, що сприяє поширенню генів альтруїстичної поведінки, це ніяк не означає ні те, що феномен альтруїзму повністю пояснюється генами, ні те, що я вважаю добір алелів єдиним механізмом еволюції.
Хочете повнішу картину, ніж у цій чи попередній колонці? Читайте Маркова. Ось, до речі, його презентація, на яку мені особливо приємно посилатися. Це розширена версія доповіді на нашій, харківській конференції молодих учених. Ми запрошували його пару років тому як метра, що виступає на відкритті конференції й веде круглий стіл.
До теми! Мене цікавлять специфічні особливості людських популяцій, які могли підвищувати ефективність групового добору на користь генів, що сприяють альтруїстичній поведінці. Тут я поговорю про три з них:
антагоністичні відносини між групами;
невипадковий характер формування груп;
внутрішньогрупові механізми контролю поведінки індивідів.
Я спробую відобразити ці фактори в моделі, яку я робив для минулої колонки. Звісно, це буде досить умовна модель; зрештою, я обґрунтовував її структуру способом життя якихось гермафродитів, що утворюють колонії на дні моря. Вносячи в цю модель фактори, що відображають особливості людських популяцій, я не стверджую, що вона буде коректно відображати нашу передісторію. Тут модель буде використана для іншого. Її задача - показати, що розглядувані фактори дійсно підвищують ефективність групового добору на користь альтруїстів, без кількісних оцінок такого добору. На щастя, моделі, що дозволяють кількісну оцінку, для людських популяцій уже створені. Тим не менш, коли я писав цю колонку й вовтузився з моделлю, я краще зрозумів деякі особливості нашої еволюції. Сподіваюся, що й хтось із читачів також розширить своє розуміння за її допомогою.
Характерною особливістю альтруїстичної поведінки багатьох тварин, включно з людиною, є те, що вона стосується лише особин, що входять у певні сукупності. Сірі щури самовіддано підтримують особин однієї з ними групи й украй ворожі з представниками інших груп. Люди часто ділять оточуючих на «своїх» і «чужих», демонструючи до них зовсім різне ставлення.
Такий альтруїзм, обмежений рамками певної групи, отримав назву «парафіяльного альтруїзму (parochial altruism)». Назва пов'язана з типологією політичних культур за Габріелем Алмондом і Сіднеєм Вербою, американськими політологами. Вони виділили три типи культур: парафіяльна, залежна й культура участі. Парафіяльний альтруїзм - частина парафіяльної культури. У російськомовній літературі (наприклад, у текстах Олександра Маркова) його називають не російським словом, а з використанням транслітерації - «парохіальним альтруїзмом». Іноді Марков пише про «парохіалізм», розуміючи під ним, імовірно, саме агресію щодо чужаків. У використанні транслітерації є свої плюси (парафіяни жодних парафій не образяться) і мінуси (термін стає непрозорим).
Ображатися тут немає на що. Парафії бувають різними, але в цілому зв'язок підтримки «своїх» і ненависті до «чужих» з належністю до певних релігійних громад показаний для різних людських культур. Цікавий висновок у цитованій статті такий: схильність до парафіяльного альтруїзму не корелює з частотою молитов, але значимо пов'язана з частотою відвідування церков і громад. Із шести вивчених конфесій найсильніше цей ефект проявився в парафіян російської православної церкви.
Отже, парафіяльний (парохіальний) альтруїзм проявляється одночасно в любові до свого, споконвічного (її можна називати патріотизмом) і в ненависті до чужого (ксенофобії, шовінізмі тощо, тощо). Є серйозні підстави припускати, що в розвитку такої поведінки великий внесок генетичних, біологічно зумовлених факторів. Люди, схильні до такої поведінки, особливо затребувані під час воєн і конфліктів. Ми пам'ятаємо їх як героїв, що поєднували самовідданість, щиру відданість братам по зброї й ненависть до ворогів. У мирний час така психологія набагато менш адекватна. Тому, зокрема, я стримано ставлюся до спроб пропагандистів патріотизму виховувати молодь на прикладі героїв війни...
Як відобразити парафіяльний альтруїзм у моїй моделі? Наслідком антагонізму між субпопуляціями є те, що частина з них гине. Якщо ви не забули, на кожному ході моєї моделі відбувається випадкове формування семи субпопуляцій. Для них обчислювалася групова виживаність, тим вища, чим більше було в цій субпопуляції альтруїстів. Я використав просту міру «сили» субпопуляції: добуток її чисельності та виживаності. Усі субпопуляції, «сила» яких нижча за середній рівень, гинуть: їх знищують численніші й міцніше згуртовані конкуренти.
Знаєте, така перебудова моделі не привела до суттєвого підвищення ефективності підтримки альтруїзму добором. Принаймні, у тому варіанті, який я вибрав, перемагають не лише групи з високою часткою альтруїстів, але й просто-напросто численніші групи. Це реалістично: великі групи, навіть складені з менш героїчних бійців, мають свої переваги. Але якщо формування субпопуляцій відбувається випадково, частка альтруїстів у великих групах виявляється ближчою до середньої по популяції. У таких умовах на динаміку алелів сильніше впливає проста випадковість.
Але чи завжди справедливе припущення, що співвідношення егоїстів і альтруїстів у субпопуляціях визначається випадково? Чи не буває так, що частка альтруїстів у багатьох групах систематично перевищує той рівень, який треба очікувати згідно з пуассоновим розподілом? Слід урахувати процеси, що приводять до таких відхилень. Перший - близькоспоріднене розмноження в малих групах. Другий - асортативність при утворенні груп, тобто наявність певних переваг при їх формуванні.
У моделі відображена асортативність, здатність альтруїстів і егоїстів групуватися «за інтересами». При формуванні субпопуляцій особини додаються в них одна за одною. Імовірності додавання особин з генотипом AA («чистих альтруїстів»), aa («чистих егоїстів») і Aa (ні те, ні се) залежить від частот алелів A і a в популяції. Зі зростанням чисельності альтруїстів у субпопуляції зростає ймовірність додавання до неї наступних альтруїстів (за рахунок зменшення ймовірності додавання особин Aa); те саме стосується й накопичення егоїстів.
Після описаних змін ефективність підтримки альтруїстів груповим добором різко збільшилася. Але перед тим, як показати результат, розповім ще про одну зміну. Не лише індивіди впливають на властивості групи, до складу якої вони входять. Групи теж впливають на поведінку особин, що їх складають. Наведу два приклади таких впливів. Перший. У колоніях суспільних комах робочі особини, які не повинні розмножуватися, контролюють одна одну. Якщо, наприклад, робоча оса егоїстично почне відкладати власні яйця (замість альтруїстичного догляду за яйцями цариці), її приплід буде знищений іншими робочими.
Другий. У групах людей, де прийнято допомагати одне одному, егоїст, що відмовляється від прийнятих норм поведінки, може відчути суспільне засудження, яке змусить його змінити свою поведінку. Згадайте: у минулій колонці я згадував реципрокний (взаємний) альтруїзм. Його наслідком є здатність особин оцінювати одна одну за готовністю допомагати ближнім. Наслідком роботи цього механізму може стати феномен репутації. Особини, що діють альтруїстично, заробляють хорошу репутацію, що сприяє підвищенню їхнього статусу (і, наприклад, збільшенню кількості їхніх нащадків). У таких групах можуть поводитися альтруїстично не лише носії генів, що сприяють альтруїстичній поведінці, але й генетичні егоїсти!
У моїй моделі це відображається так. Після формування субпопуляції в ній визначається співвідношення «чистих альтруїстів» (AA) і «чистих егоїстів» (aa). Якщо альтруїстів більше, особини Aa поводяться як альтруїсти (і з точки зору життєздатності групи, і з точки зору своїх шансів на виживання), якщо більше егоїстів - як егоїсти. При рівності кількості особин з груп AA і aa характеристики представників генотипу Aa виявляються проміжними між двома крайнощами.
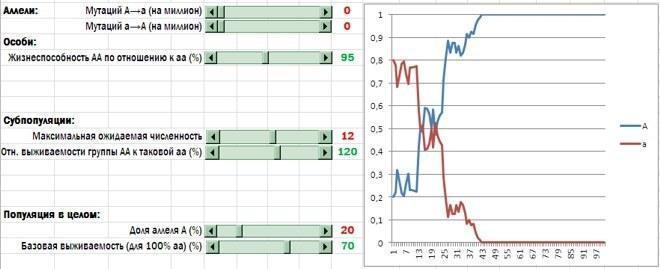
Отже, я вніс у модель аналогії групового антагонізму, асортативності формування груп і дії внутрішньогрупових норм поведінки. Етап безстатевого розмноження особин, що залишилися, став не потрібний, і я його прибрав. Ось змінена (і значно спрощена!) модель. Хоча вона описує інший спосіб популяційного відтворення, ніж характерний для людини, для нас важливо, що зроблені зміни сприяли посиленню добору на користь альтруїстів.
Порівняйте цей скрин з рисунками в минулій колонці. Описані тут зміни значно спростили модель і привели до посилення підтримки альтруїзму. Тепер альтруїсти систематично перемагають і при таких значеннях вихідних параметрів моделі, які в іншому випадку майже не залишали б їм шансів
Так, до речі, наведені тут міркування можуть допомогти не лише в розумінні еволюції альтруїзму. Вони, крім іншого, допомагають шукати відповіді й на інші питання, наприклад, на таке. Чому релігій багато, і вони - різні?
Можна сказати, що всі люди тягнуться до однієї й тієї самої істини, але знаходять її для себе по-різному. Інша (широко поширена!) версія полягає в тому, що існує одна істинна віра, а всі інші - результат бісівської спокуси, що збиває єретиків з істинного шляху. Мені подобається те пояснення, яке вважає наявність релігій та інших колективних ритуалів суттєвою частиною механізму, що створив нас у ході еволюції.
І дивіться, що виходить. Розглянуті нами фактори посилюють дію один одного. Антагонізм між групами приводить до того, що успішнішими з них виявляються ті, що об'єднані парафіяльним альтруїзмом; парафіяльний альтруїзм посилює антагонізм між групами. Невипадкове формування груп і виникнення механізмів, що підтримують їхню цілісність, сприяє перемозі субпопуляцій, об'єднаних парафіяльним альтруїзмом. У міру еволюції альтруїзму в міжгруповій конкуренції перемагають усе більш згуртовані групи. Прихильність вірі батьків - частина механізму, що забезпечував нашу еволюцію. Але в сліпої віри - злі очі...
Комплекс з розглянутих нами механізмів об'єднаний позитивними зворотними зв'язками. Однією з цікавих особливостей позитивних зворотних зв'язків є те, що їхня дія рано чи пізно переводить розглядувану систему в новий стан. Після того, як у якійсь групі тварин почалася еволюція за описаним тут механізмом, процес спрямованих змін у ній мав виявитися самопідтримуваним. Спостерігачу збоку здалося б, що він спрямований до певного результату.