Экология: биология взаимодействия. 2.14. (дополнение) Некоторые этапы истории земной жизни
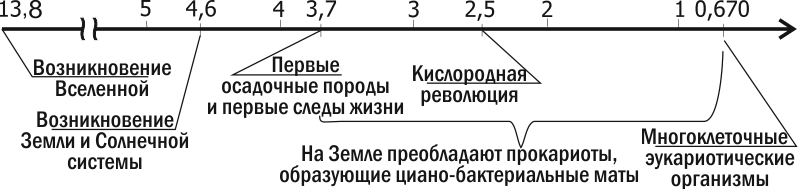
Кислородная революция была важнейшим переломом в истории Земли. Поменялся не только состав атмосферы, но и состав пород, формирующихся на поверхности Земли. Следствием кислородной атмосферы стало образование озонового слоя в атмосфере — предпосылка для заселения суши.
Українська (найновіша версія) / російська версія (оновлення припинено)
2.13. (доповнення) Геохронологічна шкала
Д. Shabanov, M. Kravchenko. Екологія: біологія взаємодії
Глава 2. Біосферологія
2.15. (доповнення) Венера, Земля, Марс
2.14. (доповнення) Деякі етапи історії земного життя
Перше появлення життя при створенні біосфери мало відбутися не у вигляді появи одного якогось організму, а у вигляді їх сукупності, що відповідає геохімічним функціям життя.
В.І. Вернадський
Поширена думка, що на якомусь етапі історії Землі виник «перший» організм, нащадки якого з’їли всі запаси органічних речовин («первинний бульйон») і дали початок усім формам життя. Звісно, такі погляди дуже наївні. Для виникнення окремого організму потрібна була неймовірна випадковість. Життя виникло в результаті поступового ускладнення геохімічних циклів у результаті відбору автокаталитичних реакцій, що забезпечували їх окремі етапи. Ще до виникнення живих організмів у цих циклах відбувалося як утворення, так і руйнування органічних речовин. Це означало, що життя виникало не у формі окремих організмів, а у формі забезпечуючих кругообіг речовин екосистем.
Біохімічні «новинки», що виникли на одному етапі геохімічного кругообігу, могли передаватися і на інші етапи. Так мали розповсюдитися способи запасання енергії, матричного синтезу біополімерів і, нарешті, клітинної організації живих систем.
У сучасних екосистемах відбувається кругообіг, основою якого є утворення органічних речовин автотрофами і їх руйнування гетеротрофами. Відходи життєдіяльності кожної з цих груп організмів є ресурсами для іншої групи. Їх чудове відповідність одне одному — наслідок того, що такий кругообіг старший за саме життя.
Одна з найдавніших осадових порід, відомих сучасній науці, знайдена у формації Ісуа в Гренландії. Вік цих порід — 3,7‑3,8 мільярда років. Ці породи утворювалися на глибині, поруч з аналогом чорного курильника; у них знайдені вуглецеві включення. Ізотопний аналіз цих вуглецевих включень свідчить, що вони є залишками живих організмів. Найдавніші залишки організмів, що мають складну структуру, знайдені в Австралії, у формації Варравуна, віком 3,5 млрд років, і в Південній Африці (формація Онфервахт, 3,4 млрд років; рис. 2.10.1). Це ціанобактерії, дуже схожі на сучасні. Це схожість стосувалася навіть біохімії. Так, у породах, утворених 3,1 млрд років, знайдено продукти розпаду хлорофілу та характерний виключно для ціанобактерій пігмент фикобілін.
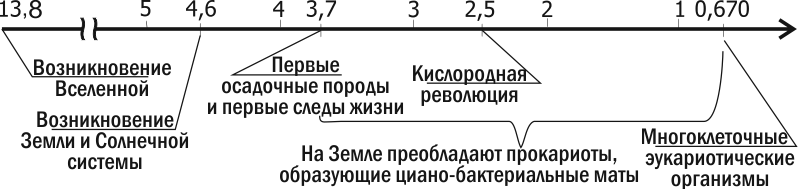
Рис. 2.10.1. Основні етапи еволюції життя на Землі
Для протерозою (а в певній мірі й археї) характерні осадові породи, які називаються строматолитами (буквально «каменними килимами»). Вони мають шарувату структуру і часто відкладалися окремими брилами. Походження строматолитів довго залишалося неясним. Їх утворення прояснила знахідка сучасних строматолитоутворювачів у Заливі Акули (Shark Bay) в Австралії. Це ізольована від океану лагуна з дуже солоною водою. На мілководді розташовані осадові гліби, поверхню яких покрито ціанобактеріальним матом. На його поверхні знаходяться ціанобактерії, а під їх шаром — дуже різноманітні бактерії інших груп, а також архебактерії. Щоб показати цю двоїстість, мат називають ціанобактеріальним, використовуючи дефіс, що він складається з ціанобактерій і інших бактерій.
Мінеральні речовини, що осідають на поверхню мата і утворюються при його життєдіяльності, відкладються шарами (приблизно 0,3 мм на рік) на його основу.
Інший тип бактеріальних екосистем відомий на території України. На Арабатській стрілці, косі в затокі Сіваш, розташовані затоплені водою та висихаючі лимани. На поверхні ґрунту в них розташований ціанобактеріальний мат товщиною до кількох сантиметрів.
Живі організми не лише залежать від середовища свого існування, а й самі впливають на нього. Земля спочатку мала атмосферу відновного характеру, у якій були стабільні окислені гази (вуглекислий газ CO₂, водяна пара H₂O, оксид сірки SO₂) і гази‑відновники (вуглекислий газ CO, водень H₂, сірководень H₂S, аміак NH₃, метан CH₄, ціан HCN, хлороводень HCl тощо). Протягом тривалого періоду історії Землі на її поверхні могли утворюватися легко окислювані породи, такі як графіт (C), лазурит (Na₂S), пірит (FeS₂) та інші.
Перші організми Землі, як автотрофи, так і гетеротрофи, були анаеробами (пристроєними до життя в безкисневих умовах). Під час фотосинтезу автотрофів виділявся вільний кисень (O₂), який був токсичним для анаеробних організмів. Спочатку він швидко окислювався відновниками, які в надлишку знаходилися в середовищі. Після того як кисень окислив основний запас відновників у середовищі, склалися відносно нейтральні умови. У результаті бактеріальні екосистеми пристосувалися до існування в умовах надлишку кисню, і поширення отримали аероби (організми, що живуть у кисневих умовах). Оскільки фотосинтез відбувався у воді, кисень міг окислювати розчинені в ній речовини, сприяючи їх осадженню. Найважливіше, що при цьому відбувалося окислення двовалентного (добре розчинного) заліза до тривалентного, що випадало в осад. Так утворювалися джеспіліти — смугові залізні руди, що є важливим джерелом цього металу для сучасного людства. Вони складаються в основному з гематиту (Fe₂O₃) і магнетиту (FeO×Fe₂O₃).
З поширенням фотосинтетик‑аеробів накопичення кисню в атмосфері продовжилося. Близько 2 млрд років тому гравітаційна диференціація Землі призвела до того, що практично все незв’язане в осадових породах залізо перейшло в ядро нашої планети. Припинення надходження заліза на поверхню Землі означало, що живі організми можуть окислювати практично всю біосферу і накопичити надлишок кисню в атмосфері. Цей перелом (що відбувався 2,5–2 млрд років тому; рис. 2.10.1) називається кисневою революцією. Проте не слід думати, що така революція була одноразовою зміною. Вона проходила через тривале балансування, коли в різних частинах біосфери склалися десь окислювальні, десь відновлювальні умови.
Киснева революція була важливим переломом в історії Землі. Змінився не лише склад атмосфери, а й склад порід, що формуються на поверхні Землі. Наслідком кисневої атмосфери стало утворення озонового шару — передумова для заселення суші.
І для прокаріот, і для еукаріот характерне утворення складних систем. У прокаріот це бактеріальні екосистеми з тісно пов’язаних особин різних видів і навіть царств (як еубактерій, так і архебактерій). Морфофункціональна різниця клітин у них — наслідок їхньої незалежної еволюції. У еукаріот це багатоклітинні організми — клони нащадків однієї клітини, різниці між якими обумовлені реалізацією різних варіантів однієї і тієї ж спадкової програми. Для розвитку еукаріотичної багатоклітинності (точніше — багатотканевості) потрібна набагато більша складність керуючих клітинних систем.
Останніми роками вдалося переконливо довести симбіогенетичну теорію походження еукаріотичної клітини. Кожна еукаріотична клітина містить геноми різного походження: у клітинах тварин і грибів це геноми ядра і мітохондрій, а у клітинах рослин — також і пластид. Невелика кільцева ДНК міститься (за багатьма даними) і в базальному тіле жгутиків клітин еукаріотичних.
Метод молекулярних годин (обліку нейтральних, з точки зору природного відбору, і ненаправлених змін послідовностей ДНК) свідчить, що еукаріоти виникли тоді ж, коли і прокаріоти. Незважаючи на це, очевидно, що протягом значної частини історії Землі домінували прокаріоти. Перші клітини, що за розмірами відповідають еукаріотичним (так звані акритархи), мають вік 3 млрд років, але їхня природа залишається неясною. Майже беззаперечні залишки еукаріот мають вік близько 2 млрд років. Лише після кисневої революції на більшій частині поверхні нашої планети склалися сприятливі для еукаріот умови. Епоха їхнього панування почалася близько 1 млрд років тому.
Ймовірно, «головним» предком еукаріотичних клітин стали архебактерії, які перейшли до живлення шляхом поглинання часток їжі. Зміна форми клітини, необхідна для такого поглинання, забезпечував цитоскелет, що складається з актина і міозину. Спадковий апарат такої клітини перемістився в глибину від її змінної поверхні, зберігши при цьому зв’язок з мембраною. Це стало причиною виникнення ядерної оболонки з ядерними порами (пов’язаної завдяки ендоплазматичній мережі з зовнішньою мембраною клітини).
Бактерії, поглинені клітиною‑хазяїном, могли продовжити своє існування всередині неї. Так, предками мітохондрій стала група фотосинтезуючих бактерій, пристосованих до життя в кисневих умовах — пурпурові альфа‑протеобактерії. Всередині клітини‑хазяїна вони втратили здатність до фотосинтезу і взяли на себе окислення органічних речовин. Завдяки їм еукаріотичні клітини стали аеробними. Симбіози з іншими фотосинтезуючими клітинами стали причиною набуття рослинними клітинами пластид. Ймовірно, жгутики еукаріотичних клітин виникли в результаті симбіозу клітин‑хазяїв з бактеріями, які, як сучасні спірохети, були здатні до вигинаючих рухів.
Сучасна фауна дає безліч прикладів існування проглочених органел і клітин у цитоплазмі клітин хижака. «Вирощують» у собі проглочені водорості деякі інфузорії, радіоляри, кишечнополостні, плоскі черв’яки, молюски та представники інших груп. Живі в кишечнику термітів жгутиколетючі вступають у тісний симбіоз зі спірохетами, які прикріплюються до їхньої поверхні. У деяких з таких симбіотичних комплексів зафіксовано навіть скорочення генетичного матеріалу у симбіонта і його залежність від речовин, синтезованих клітиною‑хазяїном. Подібні процеси відбувалися і при виникненні еукаріотичних клітин.
Спочатку спадковий апарат еукаріотичних клітин був влаштований приблизно так само, як у прокаріот (на цій стадії досі знаходяться дінофлагеляти, група одноклітинних жгутикових водоростей). Пізніше, у зв’язку з необхідністю управління більшою і складнішою клітиною, змінилася організація хромосом, а ДНК виявилася пов’язана з білками‑гістонами. Прокаріотична організація збереглася у геномах внутрішньоклітинних симбіонтів, проте частина їх функцій (у жгутиків — майже всі) була передана ядерному геному.
Різні групи еукаріотичних організмів виникали в результаті різних актів симбіогенезу. У результаті симбіогенезу еукаріотичної клітини з ціанобактеріями виникли червоні водорості. Зелені водорості виникли в результаті симбіозу з бактеріями‑прохлорофітами. Ця нещодавно виявлена група включає всього кілька сучасних видів, але при цьому перебуває у близькому родинному зв’язку з хлоропластами зелених водоростей і вищих рослин. Нарешті, хлоропласти золотистих, діатомових, бурих і криптомонадних водоростей виникли внаслідок двох послідовних симбіозів, про що свідчить наявність у них 4 мембран. Ендосимбіонтами їхніх предків стали еукаріоти, у яких перебували симбіотичні золотисті бактерії.
Розвиток життя привів до кардинального перетворення земної поверхні. Що ми побачимо, оглянувшись навколо себе за межами людських поселень? Той чи інший ландшафт, покритий характерною для кожного регіону рослинністю. У переважаючій більшості місць гірські породи покриті шаром ґрунту. Десь земну поверхню прорізають водотоки — струмки і річки. Тварини помітні значно менше, ніж рослини, але, присмотрівшись до рослинності, ми майже напевно побачимо комах, а піднявши очі вгору, розпізнаємо птахів на фоні блакитного неба. Що в цій картині є наслідком придатності нашої планети? Все! Освоюючи сушу, життя суттєво змінило її. Навіть блакитний колір неба є наслідком накопичення в атмосфері кисню. Результатом дії живих організмів є ґрунт. Він утримує на поверхні суші воду і біогени у формі, оптимальній для їх споживання організмами.
Плащовий стік з континентів, при якому вода рухалась багатьма непостійними руслами, був замінений русловим. До заселення суші дощова вода збиралася в потоки, які розмивали і виносили залишки гірських порід. Рихлі породи швидко зносились до океану, де потік води різко сповільнювався, і починало осідати осад. У сучасному світі подібні умови виникають там, де річки, що несуть воду з великою кількістю часток, впадають у море. Це призводить до формування річкових дельт — вдаючих у море ділянок «ні суші, ні моря». Прикладом таких ландшафтів може бути дельта Дунаю. Зараз такі ділянки покриває буйна рослинність, яка стабілізує в них тимчасові русла. До появи судинної рослинності цього ефекту не було, і приливно‑відливні хвилі мали постійно перетворювати таке перехідне середовище, полегшуючи вихід з води на сушу. Суша ж була сукупністю інтенсивно вивітрюваних залишків твердих порід. Масовий вихід на сушу став результатом мутуалізму (див. пункт 4.7) рослин і грибів — мікориза виявлена ще у риніофітів. Пройшовши через проміжну середу, рослинність заселила континенти. Залишки рослинних тканин стали основою для формування ґрунту. Ґрунтовий покрив закрив від вивітрювання гірські породи, а рослини захистили ґрунт від розмивання. Стік з поверхні континентів став русловим. Ґрунт затримує дощову воду, а листова поверхня збільшує площу для її випаровування (приблизно вдвічі перевищуючи площу поверхні суші). Водний обмін на континентах виявляється зміненим, причому в сприятливому для організмів напрямку.
Додаткові матеріали:
Учбова модель: Киснева революція
Колонка: Переджиття
2.13. (доповнення) Геохронологічна шкала
Д. Shabanov, M. Kravchenko. Екологія: біологія взаємодії
Глава 2. Біосферологія
2.15. (доповнення) Венера, Земля, Марс