
Кравченко (2013) Экологическая устойчивость популяционных систем ... (Автореферат)
19 июня в Днепропетровске состоялась защита кандидатской диссертации М. А. Кравченко. Здесь представлен развернутый перевод ее автореферата на русский язык и дана ссылка на украинский автореферат в формате pdf. Кравченко М. А. Экологическая устойчивость популяционных систем гибридогенного комплек...
Український автореферат (у форматі pdf) можна завантажити тут. Далі — його російський переклад.
МІНІСТЕРСТВО ОСВІТИ І НАУКИ УКРАЇНИ
ДНІПРОПЕТРОВСЬКИЙ НАЦІОНАЛЬНИЙ УНІВЕРСИТЕТ
ІМЕНІ ОЛЕСЯ ГОНЧАРА
KRAVCHENKO MARYNA OLEKSANDRIVNA
УДК (597.841::574.4):[477.5+292.485]
ЕКОЛОГІЧНА СТІЙКІСТЬ ПОПУЛЯЦІЙНИХ СИСТЕМ
ГІБРИДОГЕННОГО КОМПЛЕКСУ ЗЕЛЕНИХ ЖАБ
(PELOPHYLAX ESCULENTUS COMPLEX)
03.00.16 — екологія
АВТОРЕФЕРАТ
дисертації на здобуття наукового ступеня
кандидата біологічних наук
Дніпропетровськ — 2013
Дисертація є рукописом
Роботу виконано на кафедрі зоології та екології тварин біологічного факультету Харківського національного університету імені В. Н. Каразіна Міністерства освіти і науки України
Науковий керівник
кандидат біологічних наук, доцент
Shabanov Dmytro Andriiovych
Харківський національний університет
імені В. Н. Каразіна, кафедра зоології та
екології тварин, доцент
Офіційні опоненти
доктор біологічних наук, професор
Tsvetkova Nina Mykolaivna
Дніпропетровський національний університет імені Олеся Гончара,
кафедра геоботаніки, ґрунтознавства та екології, професор
кандидат біологічних наук
Reshetylo Ostap Stepanovych
Інститут екології Карпат НАН України,
відділ популяційної екології, старший науковий співробітник
Захист відбудеться «_19_» _червня_ 2013 року о _14.00. на засіданні спеціалізованої вченої ради Д 08.051.04 для захисту дисертацій на здобуття наукового ступеня доктора біологічних наук у Дніпропетровському національному університеті імені Олеся Гончара Міністерства освіти і науки України за адресою: 49010, м. Дніпропетровськ, проспект Гагаріна, 72, корпус 17, факультет біології, екології та медицини, ауд. 611.
З дисертацією можна ознайомитися у науковій бібліотеці Дніпропетровського національного університету імені Олеся Гончара за адресою: м. Дніпропетровськ, вул. Казакова, 8.
Автореферат розіслано «_15_» _травня_ 2013 р.
Вчений секретар
спеціалізованої вченої ради
кандидат біологічних наук, доцент Dubyna A. A.
ЗАГАЛЬНА ХАРАКТЕРИСТИКА РОБОТИ
Актуальність теми. Важливою проблемою сучасної екології є дослідження закономірностей змін біосистем у часі. Відносно нещодавно сучасній біології стали відомі незвичайні біосистеми, що виникають у гібридогенних комплексах видів, наприклад у групі зелених жаб, Pelophylax esculentus complex (огляд: Plötner, 2005). Це популяційні системи, в яких спільним розмноженням об'єднані особини різних батьківських видів та їх гібриди, що відрізняються за складом геномів диплоїдних і триплоїдних особин, а також за складом геномів, які вони клонально передають у статеві клітини. Закономірності динаміки таких біосистем залишаються практично недослідженими. Ця робота присвячена дослідженню змін таких популяційних систем гібридогенного комплексу зелених жаб.
Міжвидова гібридизація зелених жаб була відкрита майже пів століття тому (Berger, 1964), але багато особливостей успадкування у гібридів, відтворення популяційних систем були описані лише на початку XXI століття. У 2004 році на території Східної України були виявлені популяційні системи жаб (Borkin et al, 2004), які за своїм складом є одними з найскладніших серед описаних на даний час (Шабанов, Литвинчук, 2010; Коршунов, 2010). Дослідники цього центру зіткнулися з тим, що для опису характерних для нього популяційних систем відсутня навіть необхідна понятійна база (Шабанов та ін., 2009).
Популяційні системи гібридогенних комплексів видів — це системи, безпосереднє дослідження яких стикається зі значними труднощами. Інтегрувати наявні знання про популяційні системи можна за допомогою імітаційного моделювання. Моделювання допомагає і при вирішенні однієї з найменш вивчених досі проблем: з'ясування того, як можуть зберігати стійкість настільки складні популяційні системи, як зареєстровані у зелених жаб. Дослідження цих питань має значення не лише для вивчення особливостей популяційних систем Pelophylax esculentus complex, але й для розвитку нашого розуміння загальних властивостей інших біосистем, які є об'єктами екології, — популяцій, біоценозів та екосистем.
Зв'язок роботи з науковими програмами, планами, темами. Роботу виконано відповідно до науково-дослідних тем кафедри зоології та екології тварин біологічного факультету Харківського національного університету імені В. Н. Каразіна:
— «Оцінка впливу кліматичних та антропогенних факторів на динаміку спільнот тварин у Північно-Східній Україні» (0106U001580; 2006-2008 рр.),
— «Оцінка факторів, що визначають просторову, етологічну та генетичну структуру популяцій тварин» (0109U001443; 2009-2010 рр.);
— «Неортодоксальне видоутворення (гібридизація, неменделівське успадкування, поліплоїдія та сітчаста еволюція): вивчення на прикладі амфібій» (0109U007439; 2009-2010 рр.);
— «Розробка технологій отримання клонального потомства хребетних тварин шляхом схрещування напівклональних міжвидових гібридів (на прикладі зелених жаб)» (0112U001026; 2012-2014 рр.).
Мета та завдання дослідження. Мета роботи — визначити механізми екологічної стійкості та змін складу популяційних систем гібридогенних комплексів видів на прикладі зелених жаб (Pelophylax esculentus complex).
Для досягнення зазначеної мети в роботі поставлені такі завдання:
— встановити положення популяційних систем зелених жаб (Pelophylax esculentus complex) в ієрархії біосистем; дослідити специфіку таких систем та процесів змін їх складу;
— визначити фактори, що впливають на склад популяційних систем зелених жаб; розробити концептуальну модель динаміки таких систем, що враховує дію цих факторів; створити імітаційну модель трансформацій популяційних систем зелених жаб;
— у ході експериментів з імітаційною моделлю визначити можливі стани популяційних систем зелених жаб та закономірності переходів між ними; порівняти ці результати з емпіричними даними;
— класифікувати стани досліджуваних систем з точки зору їх типології та екологічної стійкості.
Об'єкт дослідження — популяційні системи гібридогенного комплексу зелених жаб (Pelophylax esculentus complex).
Предмет дослідження — трансформації популяційних систем зелених жаб, їх причини та механізми, а також різноманіття станів таких систем.
Методи дослідження — польові обліки зелених жаб, оцінка чисельності популяційної системи з використанням мічення та повторного відлову, первинне визначення видової належності та плоїдності особин, формалізація емпіричних та літературних даних, імітаційне моделювання, експерименти з імітаційною моделлю, порівняння результатів моделювання з емпіричними даними.
Наукова новизна отриманих результатів. Результати, що відображають наукову новизну, полягають у наступному:
вперше:
— запропоновано поняття «геміклональна популяційна система» (ГПС);
— обґрунтовано, що ГПС гібридогенних комплексів видів є особливим рівнем організації біосистем;
— створено концептуальну модель динаміки ГПС, що враховує внутрішні та зовнішні фактори їх динаміки;
— показано, що геміклональне успадкування, характерне для міжвидових гібридів, може призводити до зміни складу ГПС, досягнення ними стану стійкості або їх загибелі;
— розроблено імітаційну модель трансформацій ГПС зелених жаб на основі різницевих рівнянь на базі Microsoft Excel;
— розглянуто при експериментуванні з моделлю можливі шляхи перетворень ГПС зелених жаб;
— розроблено та обґрунтовано динамічну типологію станів ГПС зелених жаб з точки зору екологічної стійкості як здатності зберігати або змінювати свій тип у часі;
удосконалено:
— уявлення про ієрархію рівнів організації біосистем;
— понятійну базу, що стосується гібридогенних комплексів видів з геміклональним відтворенням гібридів;
— уявлення про причини зміни типів ГПС у часі;
— уявлення про різноманіття станів екологічної стійкості біосистем.
Практичне значення отриманих результатів. Концептуальна модель трансформацій популяційних систем гібридогенного комплексу видів (на прикладі зелених жаб) може бути використана для створення інших імітаційних моделей, що описують не лише зелених жаб, а й інші гібридогенні комплекси видів. Зокрема, вона використовується при виконанні ініціативного проекту Харківського національного університету імені В. Н. Каразіна та Каледонського університету Глазго (Великобританія).
Результати роботи важливі для дослідження механізмів екологічної стійкості біосистем, вони можуть використовуватися для моніторингу стану природних об'єктів, організації їх охорони, визначення ГПС, що потребують збереження.
Модель трансформацій ГПС є способом інтегрувати наявні дані з популяційної екології зелених жаб, а також встановити, які дані є недостатніми, і тим самим сприяти їх отриманню.
Результати роботи використовуються при читанні загальних та спеціальних курсів «Екологія», «Моделювання в екології» та «Батрахологія і герпетологія», а також при проведенні навчально-польової практики з зоології хребетних на біологічному факультеті ХНУ імені В. Н. Каразіна.
Особистий внесок здобувача. Дисертаційна робота є самостійним та оригінальним дослідженням. Здобувач самостійно розробила програму досліджень та обрала методики. Напрями роботи були визначені спільно з науковим керівником. Польові дослідження, обробка даних, розробка концептуальної моделі трансформацій популяційних систем зелених жаб, створення імітаційної моделі та ряд експериментів з нею, узагальнення та інтерпретація отриманих результатів проведено здобувачем самостійно або за її безпосередньої участі.
Низка публікацій, що відображають розглянуті в дисертації теми, підготовлені у складі робочої групи популяційної екології амфібій ХНУ імені В. Н. Каразіна. У цих роботах автор брала участь у виробленні плану досліджень, виконувала постановку завдання для моделювання, створювала моделі, експериментувала з ними та брала участь в інтерпретації результатів.
Апробація результатів дисертації. Матеріали дисертації оприлюднені на III та IV Міжнародних наукових конференціях Zoocenosis-2005 та Zoocenosis-2007 (Дніпропетровськ); науковій конференції «Сучасні проблеми зоології та екології» (Одеса, 2005); IX та XI Міжнародних науково-практичних екологічних конференціях, присвячених популяційній екології (Бєлгород, 2006 та 2010); III та V з'їзді Герпетологічного товариства ім. А. М. Нікольського (Пущино, 2006; Мінськ, 2012); науковій конференції біологічного факультету ХНУ, присвяченій 100-річчю з дня народження Є.І. Семененка (Харків, 2006); Міжнародній конференції «Сучасні проблеми біологічної еволюції» (Москва, 2007); III, IV та VI з'їздах Українського герпетологічного товариства (Мелітополь, 2007; Дніпропетровськ, 2008; Київ, 2011); III Міжнародній конференції молодих учених «Біологія: від молекули до біосфери» (Харків, 2008); з'їзді Екологічної ліги (Київ, 2008), Міжнародній науково-практичній конференції «Біорізноманіття та сталий розвиток» (Сімферополь, 2010); І Всеукраїнській науково-практичній конференції «Сучасна проблематика та методологія біоекологічних досліджень: популяційний підхід» (Івано-Франківськ, 2012).
Публікації. За матеріалами дисертації опубліковано 20 наукових публікацій, у тому числі 11 статей у профільних наукових журналах та збірниках наукових праць (з них 10 у профільних журналах України) і 1 стаття у виданні іноземної держави, а також 9 наукових публікацій у матеріалах та тезах конференцій. Крім того, матеріали дисертації використано у трьох підготовлених за участі дисертанта навчально-методичних виданнях.
Структура та обсяг дисертації. Дисертація складається з переліку умовних позначень, вступу, 6 розділів, висновків, списку використаної літератури, 2 додатків, ілюстрована 7 таблицями та 26 рисунками. Матеріали дисертації викладено на 159 сторінках комп'ютерного набору, з яких основного тексту 127 сторінок. Список використаної літератури містить 165 джерел, серед яких 44 — латиницею.
ОСНОВНИЙ ЗМІСТ РОБОТИ
СУЧАСНИЙ СТАН ПРОБЛЕМИ
Гібридогенез Pelophylax esculentus complex. До європейських зелених жаб, Pelophylax esculentus complex, належать батьківські види: ставкові жаби, Pelophylax lessonae (Camerano, 1882), та озерні жаби, Pelophylax ridibundus (Pallas, 1771), а також їх геміклональні міжвидові гібриди — їстівні жаби, Pelophylax esculentus (Linnaeus, 1758). При гаметогенезі у P. esculentus у статеві клітини в типовому випадку переходить один із батьківських геномів клонально, без рекомбінації (рис. 1).
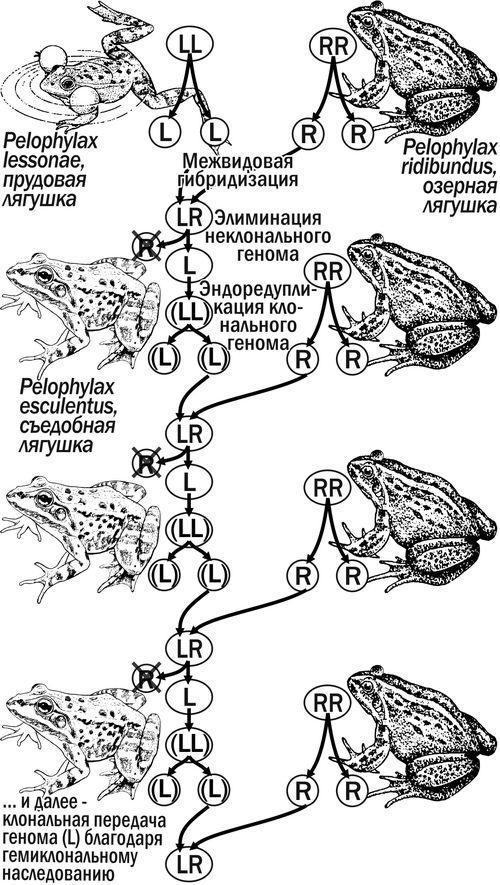
Рис. 1. Поява P. esculentus завдяки міжвидовій гібридизації та його відтворення при схрещуванні з представниками батьківського виду (за Plötner, 2005). Літерою L позначено геном P. lessonae, літерою R — геном P. ridibundus, клональність геному показано розміщенням його символу у дужках
P. esculentus відтворюються найчастіше у популяційних системах, що складаються як з гібридів, так і з особин батьківських видів, які розмножуються спільно. Різні каріогенетичні форми P. esculentus відрізняються за кількістю та складом геномів і за характером гаметогенезу (Plötner, 2005).
Номенклатурні та термінологічні проблеми дослідження Pelophylax esculentus complex. Обговорюється використання родової назви Pelophylax, застосування для гібридних жаб назви, подібної до видової, належність до P. esculentus триплоїдів, поняття «геміклональність» та «популяційна система».
Різноманіття популяційних систем Pelophylax esculentus complex. Обговорюється різноманіття популяційних систем зелених жаб, наведено їх найпоширенішу класифікацію. Найбільше різноманіття популяційних систем було зареєстровано в регіоні, що отримав за участі автора роботи назву Сіверсько-Донецького центру різноманіття P. esculentus complex; більша частина цього центру розташована в Харківській області (Шабанов та ін., 2009; Шабанов, Литвинчук, 2010).
Рівні організації біосистем, їх емерджентні властивості та процеси динаміки. Концепція структурних рівнів організації біосистем є результатом застосування принципів загальної теорії систем до опису біологічних явищ. Виділяти окремий рівень організації біосистем слід тоді, коли на ньому відбувається виникнення нових якостей у результаті взаємодії підсистем (Тахтаджян, 2001). Кожному рівню відповідають характерні процеси динаміки та певний рівень біорізноманіття.
Екологічна стійкість біосистем та фактори, що впливають на неї. Поняття стійкості є одним з найважливіших для екології, але не має загальноприйнятого визначення. Проведено аналіз його використання, встановлено, що найбільш загальною є трактування стійкості як здатності системи зберігати певні ознаки після змін, спричинених певними факторами (Артюхов, 2009). Цей підхід збігається з математичним трактуванням стійкості, яке було запропоновано в 1892 р. професором Харківського університету А. М. Ляпуновим (1950).
Основні принципи моделювання динаміки біосистем. Розглянуто системні принципи, які слугують основою моделювання. Модель має бути спрощеною порівняно з дійсністю, але має відповідати оригіналу за структурою зв'язків між підсистемами. Порівнюються аналітичні та імітаційні моделі. Імітаційні моделі відображають структуру оригіналу та ті процеси, що відбуваються в ньому.
Моделювання біосистем популяційного рівня організації. Математичне вивчення популяційного росту розпочато Л. Фібоначчі на початку XIII ст. Важливі результати при моделюванні динаміки популяцій отримано Т. Мальтусом у XVIII ст., Ф. Ферхюльстом у XIX ст., А. Лоткою та В. Вольтерра у XX ст. З 60-х років минулого століття розпочалася епоха широкого застосування моделювання для дослідження популяцій, екосистем та біосфери загалом (Медоуз та ін., 2007; Коросов, 2002).
МАТЕРІАЛ ТА МЕТОДИ ЙОГО ДОСЛІДЖЕННЯ
Методи дослідження природних популяційних систем зелених жаб. Матеріал було зібрано під час польових сезонів 2004-2012 рр. на території Сіверсько-Донецького центру різноманіття Pelophylax esculentus complex. Обговорено методи визначення жаб, мічення, обліку за методом Петерсона, визначення плоїдності.
Емпіричний та теоретичний матеріал, використаний при створенні моделі. Застосовано оригінальні дані, результати дослідження різноманіття ГПС групи популяційної екології амфібій кафедри зоології та екології тварин Харківського національного університету імені В. Н. Каразіна та літературні дані.
Методика експериментів з імітаційною моделлю трансформацій ГПС. Обговорюється використання імітаційної моделі ГПС зелених жаб. Для якісної оцінки адекватності моделі слід встановити, чи достатньо факторів, передбачених у моделі, щоб викликати динаміку, що відповідає змінам природних систем, а також чи збігається розподіл станів ГПС, отриманих у результаті моделювання, з описаним у ході збору емпіричних даних.
ТРАНСФОРМАЦІЇ ГЕМІКЛОНАЛЬНИХ ПОПУЛЯЦІЙНИХ СИСТЕМ ЗЕЛЕНИХ ЖАБ ТА ПРОБЛЕМИ ЇХ ВИВЧЕННЯ
ГПС як особлива категорія біосистем. Гібридогенні комплекси видів утворюють особливі біосистеми, що складаються з особин різних видів, об'єднані спільним розмноженням (рис. 1). Загальноприйнята назва «популяційні системи» є для них недостатньо конкретною. Пропонується поняття геміклональної популяційної системи (ГПС). ГПС — це сукупність особин батьківських видів у складі гібридогенного комплексу, а також їх гібридів, об'єднаних спільним перебуванням та розмноженням, у якому з покоління в покоління передаються як клональні, так і рекомбінантні геноми. Розташування ГПС в ієрархії біосистем показано на рис. 2.
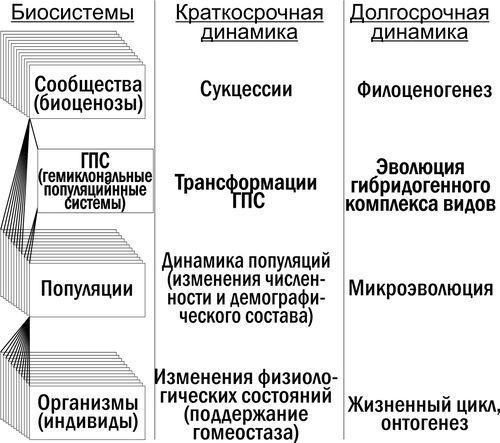
Рис. 2. {"translated":"Положення ГПС у ієрархії біосистем. Між популяційним і біогеоценотичним рівнем для деяких груп видів (точніше — для гибридогенних комплексів видів) «вбудовується» рівень ГПС
ГПС зелених жаб Іськового ставка: приклад зареєстрованих трансформацій. Іськовий ставок розташований поблизу біологічної станції ХНУ імені В. Н. Каразина та НПП «Гомольські ліси». ГПС зелених жаб цього ставка стала модельною для дослідження трансформацій завдяки досить довгій історії її спостережень. У 1995 р. російський батрахолог Г. А. Лада зареєстрував тут популяційну систему, що складалася майже виключно з диплоїдних P. esculentus. Ця ГПС зазнала антропогенного впливу: у 2000 р. ставок був спущений, у 2001 частково заповнений. У 2006 р. ми зареєстрували ГПС з критично низькою ефективною чисельністю, яка складалася з диплоїдних самців P. esculentus. Розмноження відбувалося завдяки самкам P. ridibundus, які потрапляли у ставок з інших водойм. Головастики та сеголетки у ставку практично відсутні. У наступні роки кількість жаб на Іськовому ставку зменшувалася, але в ньому з’явилися самки P. esculentus. У 2011 та 2012 рр. тут реєструються самки, головастики та сеголетки.
Можливі механізми змін ГПС Іськового ставка. Криза, через яку пройшла ГПС Іськового ставка, свідчить, що під час спуску водойми з нього зникли всі клональні геноми, крім чоловічого геному P. lessonae: (YL). У ставку переважно відбувалися схрещування ♀XRXR × ♂(YL)XR → ♂(YL)XR, усі нащадки яких є самцями (YL)XR. Збільшення їх кількості в ГПС до критичного рівня могло привести її до загибелі. Але, на щастя, в ГПС Іськового ставка потрапили й інші клональні геноми.
Оцінка чисельності ГПС зелених жаб Іськового ставка. Для детального опису подальших змін складу ГПС Іськового ставка застосовано облік статево зрілих особин за Петерсоном (Коли, 1979). У 2011 році нерестового стада складало 358 ± 55 особин, кількість самок була менша за кількість самців приблизно в 10 разів і була недостатньою для точної оцінки. У 2012 році чисельність ГПС стала значно більшою: 697 ± 224 особин. Облік самців і самок окремо показує, що співвідношення ♂♂:♀♀ ≈ 5:1. Таким чином, зафіксовано зріст чисельності ГПС і збільшення частки самок. У ГПС Іськового ставка переважають диплоїдні P. esculentus, але знайдені одиничні P. ridibundus і триплоїдні P. esculentus.
Таким чином, зафіксовано зміни складу природної ГПС. Теоретичний аналіз дозволяє вважати, що ці зміни належать до особливої категорії процесів динаміки біосистем. Для дослідження таких процесів доцільно використовувати імітаційне моделювання.
РАЗРОБКА КОНЦЕПТУАЛЬНОЇ МОДЕЛІ ТРАНСФОРМАЦІЙ ГЕМИКЛОНАЛЬНИХ ПОПУЛЯЦІЙНИХ СИСТЕМ ЗЕЛЕНИХ ЖАБ
Призначення та принцип використання моделі трансформацій ГПС. Можливе застосування моделей для розвитку уявлень про трансформацію ГПС показано на рис. 3. Процес пізнання є системою з багатьма зворотними зв’язками. Додаткові зв’язки з’являються при формалізації системи уявлень, що склалася під час дослідження певного феномена. Формалізація необхідна для розробки концептуальної моделі: опису структури підсистем і зв’язків між ними. На основі концептуальної моделі можна розробляти різні імітаційні моделі. Результати моделювання мають бути порівнянні з емпіричними даними.
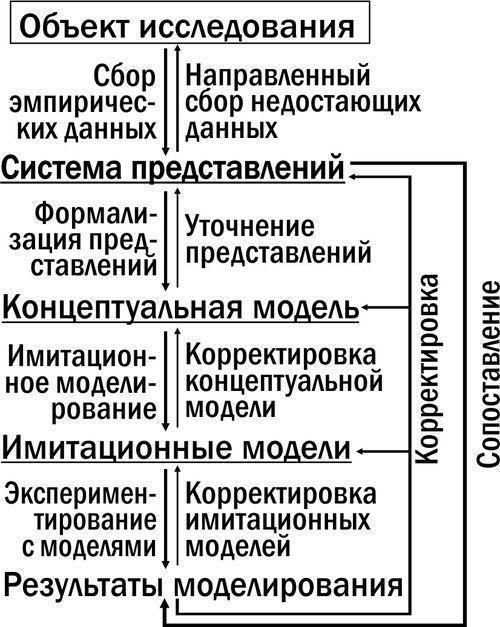
Рис. 3. Роль моделювання у розвитку системи наукових уявлень про об’єкт дослідження. Різні моделі підкреслені
Теоретичні уявлення, закладені в основу моделі. Гіпотеза, на якій базується модель, полягає в тому, що різні типи ГПС зелених жаб можна розглядати як етапи їх трансформацій. Так, потрапляння в популяцію P. ridibundus хоча б однієї особини P. esculentus з клональним геномом (YL) при нормальній життєздатності самців (YL)XR призведе ГПС до загибелі у випадку, якщо в неї не потраплять інші клональні геноми. Результати теоретичного аналізу можливих шляхів зміни ГПС зелених жаб, отримані на етапі формалізації уявлень при розробці концептуальної моделі, показані на рис. 4.
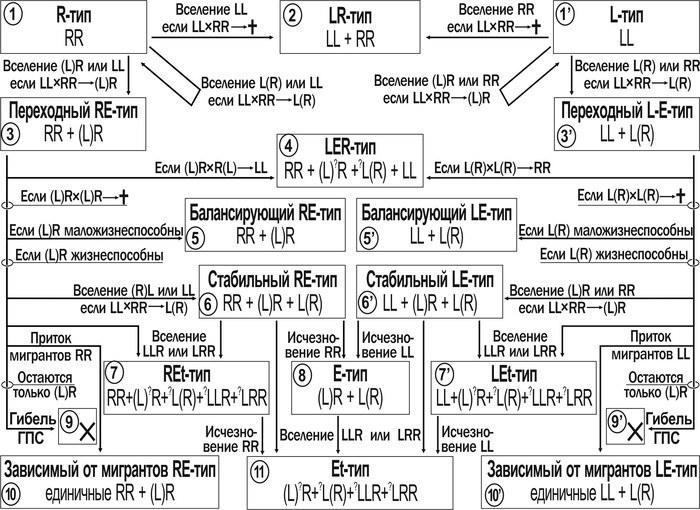
Рис. 4. Апріорні (відносно імітаційного моделювання) уявлення про можливі шляхи трансформації ГПС зелених жаб. Схема симетрична щодо батьківських видів: P. lessonae і P. ridibundus
Долю ГПС визначає те, які клональні геноми в неї потрапили, а також те, чи є життєздатними гибридні особини, чи ж вони поступаються у життєздатності особинам батьківських видів. Важливо зазначити, що за нашими та літературними даними особини батьківських видів, що утворюються від схрещування гибридів (їх виникнення називається гибридолізом), гинуть до досягнення статевої зрілості: ♀(XL)XR × ♂(YL)XR → [♂XLYL → †].
Екологічні особливості зелених жаб, суттєві для моделювання їх популяційних систем. Розглянуто річний цикл зелених жаб, описані процеси, які відбуваються в популяціях і мають враховуватись при побудові моделі.
Екологічні фактори, дія яких має враховуватись у моделі трансформацій ГПС зелених жаб. Модель має бути максимально простою і при цьому відповідати дійсності в важливих з точки зору динаміки ГПС обставинах. Доцільно не імітувати всю різноманітність факторів, що діють на досліджувані об’єкти (рис. 5). Відповідно до підходу, запропонованого А. Н. Колмогоровим, у моделі розглядаються змінні, які відображають інтегральну дію факторів середовища. Ці змінні ми називаємо параметрами життєздатності.
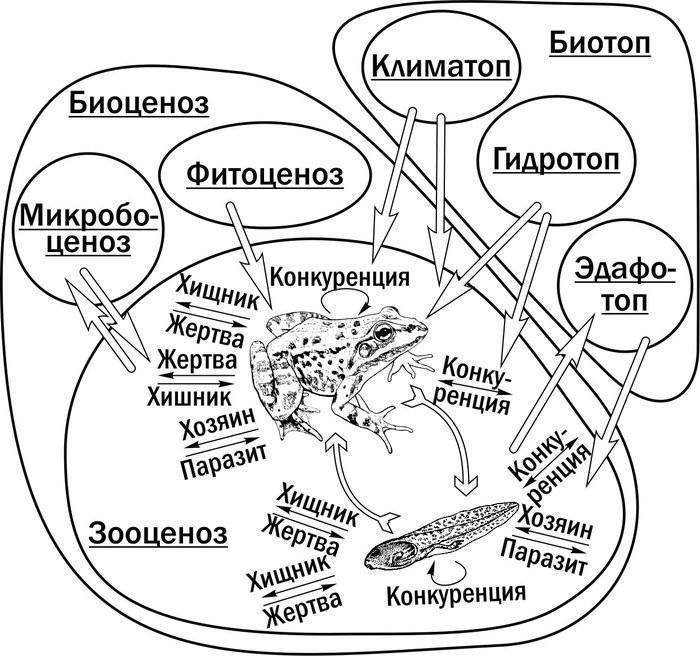
Рис. 5. Найважливіші групи екологічних факторів, що впливають на динаміку ГПС зелених жаб, накладені на схему біогеоценозу за В. Н. Сукачевим. Популяції зелених жаб складаються з геміпопуляцій (за В. Н. Беклемішевим) личинок (головастиків) і дефінітивних особин (жаб)
Різницеві рівняння для опису популяційної динаміки. Для дослідження ГПС пропонується модель з дискретним часом і перекриваючимися поколіннями, побудована на різницевих рівняннях. На кожному кроці кількість представників кожної з груп жаб (виділених за генотипом і віком) обчислюється за даними про таку кількість на попередньому кроці з урахуванням впливу факторів.
Параметри життєздатності. У моделі використуються наступні параметри життєздатності: тривалість життя, вік першого нересту, виживаність, конкурентоспроможність, використання ресурсів, ймовірність розмноження, плодовитість і ймовірність повторного розмноження того ж року для самців. Вказані способи визначення цих параметрів для кожного генотипу та кожної вікової групи.
Алгоритм конкурентного скорочення чисельності. Для характеристики концептуальної моделі наведемо її фрагмент: алгоритм скорочення чисельності у випадку нестачі ресурсів. Сукупність особин з генотипами 1G, 2G, … jG представлена віковими групами: 1Ga‑1, 1Ga, 1Ga+1,…jGa‑1, jGa, jGa+1. Для кожної групи розраховані параметри життєздатності (s(jG) тощо) і конкурентоспроможності (c(jG) тощо), а також необхідна кількість ресурсів (u(jG) тощо). Припускається, що сумарне використання ресурсів не перевищує місткість середовища: .
1. За минулорічною чисельністю та виживаністю розраховується «вхідна» чисельність усіх груп: w(jGa)t = n(jGa‑1)t‑1× s(jGa).
2. Розраховується необхідна кількість ресурсів: Ot = Σ(w(jGa)t × u(jGa)).
3. Якщо Ot ≤ K (ресурсів достатньо), то для всіх груп n(jGa)t = w(jGa)t.
4. Якщо Ot > K, розраховується нормована за конкурентоспроможністю чисельність усіх груп: q(jGa)t = w(jGa)t × c(jGa)) і кількість необхідних їм ресурсів: Yt = Σ(q(jGa)t × u(jGa)).
5. Якщо Yt = K, тоді n(jGa)t = q(jGa)t. Якщо Yt > K, то кількість усіх n(jGa)t визначається відповідно 6‑му кроку алгоритму, а якщо Yt < K – 7‑му кроку.
6. Якщо Yt > K, усі групи скорочуються однаково: n(jGa)t = q(jGa)t × K/Yt.
7. Якщо Yt < K, (w(jGa)t - n(jGa)t)/(w(jGa)t - q(jGa)t) = (Ot - K) / (Ot - Yt), n(jGa)t = w(jGa)t - (w(jGa)t - q(jGa)t) × (Ot - K) / (Ot - Yt).
Перевіркою розрахунків є обчислення Vt = Σ(n(jGa)t × u(jGa)). Якщо Vt = K, то скорочення проведено правильно. Чисельність кожної групи у випадку нестачі ресурсів скоротилася пропорційно їх конкурентоспроможності.
Алгоритм утворення пар при розмноженні. Шанси на розмноження кожної особини залежать від її параметрів життєздатності. Скорочення участі у розмноженні тієї статі, яка представлена в ГПС у надлишку, проводиться аналогічно скороченню чисельності при конкуренції.
Склад нащадків при різних схрещуваннях. Специфікою гибридогенних комплексів видів є незвичні (внаслідок напівклонального успадкування) результати схрещувань за участю гибридних особин (рис. 1). Один із варіантів складу потомства від схрещувань (прийнятий у моделі за замовчуванням) показаний у табл. 1.
Таблиця 1
Генотипи, що виникають при схрещуванні різних диплоїдних форм жаб
♀XRXR
♀(XL)XR
♀XL(XR)
♀(XL)(XR)
♀XLXL
Гамети
XR
(XL)
(XR)
(XL) : (XR)
XL
♂XRYR
XR : YR
XRXR : XRYR
(XL)XR : (XL)YR
XRXR : XRYR
(XL)XR : (XL)YR : XRXR : XRYR
(XL)XR : (XL)YR
♂(XL)YR
(XL)
(XL)XR
XLXL
(XL)(XR)
XLXL : (XL)(XR)
XLXL
♂(YL)XR
(YL)
(YL)XR
XLYL
(YL)(XR)
XLYL : (YL)(XR)
XLYL
♂YL(XR)
(XR)
XRXR
(XL)(XR)
XRXR
(XL)(XR) : XRXR
XL(XR)
♂XL(YR)
(YR)
XRYR
(XL)(YR)
XRYR
(XL)(YR) : XRYR
XL(YR)
♂(XL)(YR)
(XL) : (YR)
(XL)XR : XRYR
XLXL : (XL)(YR)
(XL)(XR) : XRYR
XLXL : (XL)(YR) : (XL)(XR) : XRYR
XLXL : XL(YR)
♂(YL)(XR)
(YL) : (XR)
(YL)XR : XRXR
XLYL : (XL)(XR)
(YL)(XR) : XRXR
XLYL : (XL)(XR) : (YL)(XR) : XRXR
XLYL : XL(XR)
♂XLYL
XL : YL
XL(XR) : YL(XR)
XLXL : XLYL
XL(XR) : YL(XR)
XLXL : XLYL : XL(XR) : YL(XR)
XLXL : XLYL"}ДОСЛІДЖЕННЯ ЗАКОНСЛУГОВАНЬ ТРАНСФОРМАЦІЙ ГПС ЗЕЛЕНИХ ЖАБ ПРИ ВИКОРИСТАННІ ІМІТАЦІЙНОЇ МОДЕЛІ
Реалізація моделі на основі Microsoft Excel. На основі концептуальної моделі, яка описана в попередньому розділі, у середовищі Microsoft Excel розроблена імітаційна модель. До складу цієї моделі входять наступні функціональні блоки: блок для вводу початкового складу ГПС (з клітинками, у яких слід вказати чисельність усіх вікових груп усіх кариогенетичних форм), блок для вводу сценарію імітації (з клітинками, що відповідають вводу певної кількості жаб певної вікової групи та кариогенетичної форми на певному кроці імітації), блок для вводу параметрів життєздатності для всіх груп жаб, а також кількості ресурсів, які доступні в середовищі, блок для вводу результатів усіх можливих схрещувань, блок для покрокового розрахунку чисельності всіх груп жаб; зведені таблиці динаміки різних форм; діаграми для візуалізації отриманої динаміки.
Модель у вигляді файлу Microsoft Excel знаходиться у вільному доступі у репозиторії Центральної наукової бібліотеки ХНУ імені В. Н. Каразина за адресою https://dspace.univer.kharkov.ua/handle/123456789/2037.
Демографічні параметри модельної популяції Pelophylax ridibundus. Розглянуто роботу моделі щодо імітації простої популяції P. ridibundus. Вибрано параметри життєздатності, за яких у результаті роботи моделі виникає стабільна демографічна структура, що відповідає наявним даним, отриманим завдяки застосуванню скелетохронології та інших методів вивчення популяцій зелених жаб.
Приклад роботи моделі. Розглянемо популяцію P. ridibundus, що населяє регіон, де всі P. lessonae виникають в результаті гибридолізу (при схрещуванні гибридів) і є нежиттєздатними (т. е. (L)R × (L)R → LL → †). Попадання в таку популяцію гибридних особин, що передають геном P. lessonae, призведе до того, що весь нащадок від схрещування гибридів з особинами батьківського виду складатиметься з гибридів: (L)R × RR → (L) R. Два інші типи схрещувань, що відбуваються в такій ГПС, це: RR × RR → RR і (L) R × (L) R → LL → †. Через наростання кількості гибридів у ГПС її здатність до розмноження знижується до нуля, і така ГПС вмирає (рис. 6).
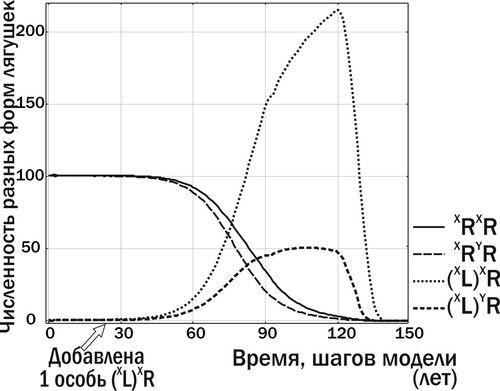
Рис. 6. Попадання в популяцію P. ridibundus однієї особини P. esculentus з клональним геномом (XL) призводить ГПС до загибелі
Попадання гибридів, що передають клональний геном P. ridibundus, у популяцію P. ridibundus, не викличе жодних довгострокових змін: усі нащадки будуть належати до батьківського виду. Але якщо такий геном потрапить у ГПС, показану на рис. 6, у якій вже значна кількість особин передає клональний геном P. lessonae, ГПС може перейти у стан стабільної рівноваги (рис. 7).
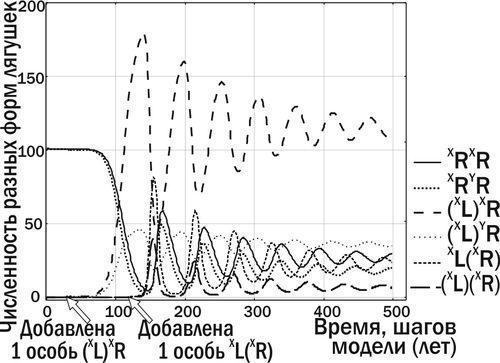
Рис. 7. Попадання в ГПС, показану на рис. 6, особини P. esculentus з клональним геномом (XR) призводить до того, що ГПС переходить у стан стійкої рівноваги
Запропонований у моделі комплекс факторів виявився достатнім для моделювання трансформацій, аналогічних тим, що були зафіксовані в природі. Наприклад, ситуація, показана на рис. 6 (за короткий час до загибелі), у певній мірі аналогічна стану, у якому ГПС Іськового ставища перебувала у 2006 році. Зараз ця ГПС знаходиться на шляху до стану стабільної рівноваги, що аналогічно перетворенням, показаним на рис. 7.
Імітаційне моделювання теоретично передбачених трансформацій ГПС зелених жаб. За допомогою імітаційної моделі проведено перевірку теоретичних уявлень, розроблених при створенні концептуальної моделі (рис. 4). В цілому ці уявлення отримали підтвердження, проте щодо ряду деталей (наприклад, умов, у яких відбір на користь особин батьківського виду може компенсувати перевагу гибридів у розмноженні) були уточнені. Це уточнення наявних уявлень було б неможливим без використання імітаційної моделі.
Фактори, що визначають напрямок трансформацій ГПС Pelophylax esculentus complex. Проведено якісний аналіз відповідності між вихідними значеннями параметрів моделі та станами модельної ГПС, до яких вона переходить у результаті імітацій. Встановлено (рис. 8), що зміни параметрів життєздатності та співвідношення різних форм у початковій ГПС впливають, перш за все, на кількісне співвідношення різних форм у складі ГПС. Коли накопичення кількісних змін переходить певний поріг, модельна ГПС переходить у інший стан. Зміна результатів схрещувань (що відображають дію геміклонального успадкування) та початкового складу клональних геномів у модельній ГПС визначають, перш за все, якісний склад кариогенетичних форм, що реєструються в ГПС наприкінці імітацій.
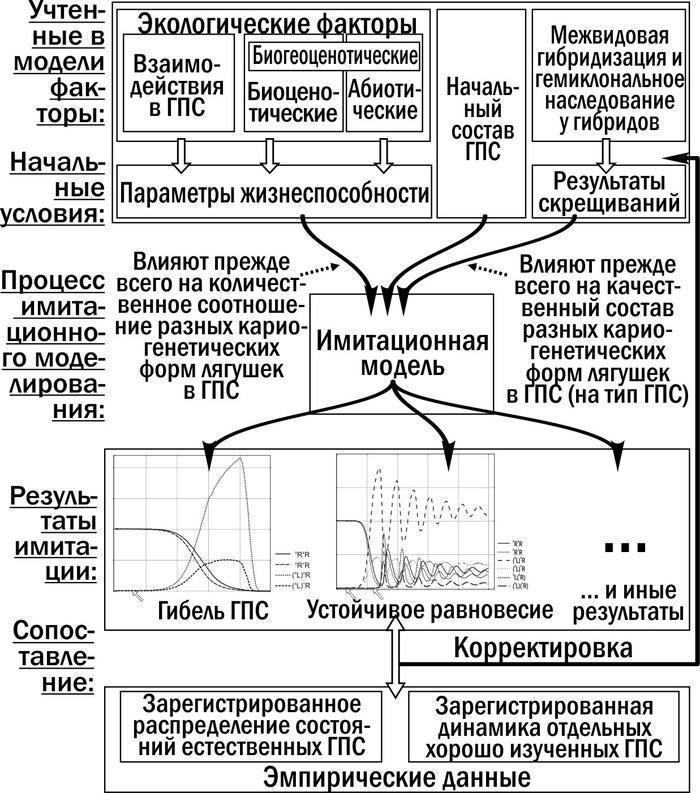
Рис. 8. Схема порівняння результатів роботи моделі з емпіричними даними
РАЗНОМАНІТТЯ СТАНІВ ПОПУЛЯЦІЙНИХ СИСТЕМ ЗЕЛЕНИХ ЖАБ З ПЕРЕГЛЯДУ ЇХ ЕКОЛОГІЧНОЇ СТІЙКОСТІ
Аналіз фазового простору ГПС RE-типу з жіночими клональними геномами. Способом аналізу трансформацій ГПС є візуалізація їх траєкторій у фазовому просторі станів системи (рис. 9). Перехід до стійкої рівноваги, аналогічний зображеному на рис. 7, показано на рис. 10. У випадку, коли клональний геном P. ridibundus надто пізно потрапляє у ГПС з переважаючими гибридами, що передають геном P. lessonae, ГПС вмирає, а її траєкторія описує петлю у фазовому просторі (рис. 11).
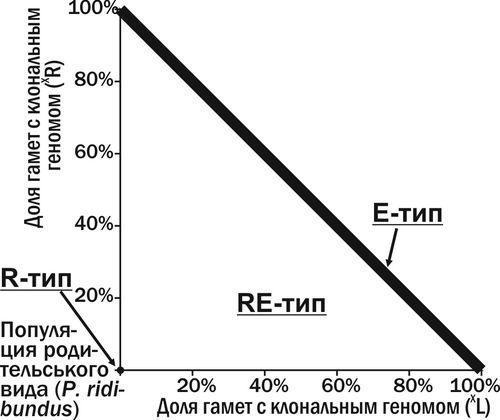
Рис. 9. Типи ГПС (за складом) у фазовому просторі станів ГПС з P. ridibundus та гибридами, що передають жіночі клональні геноми
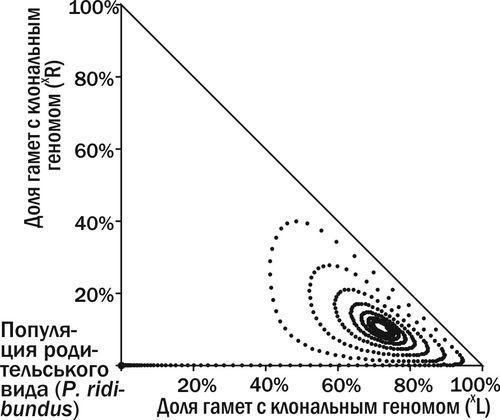
Рис. 10. ГПС переходить у стан стійкої рівноваги (фазовий простір як на рис. 9)
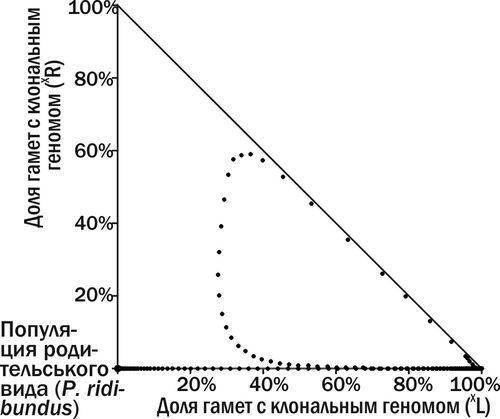
Рис. 11. Геном (XR) пізно потрапив у ГПС, де накопичуються носії геному (XL). ГПС вмирає, описавши петлю у фазовому просторі
У фазовому просторі можна виділити зони, що відповідають певним атракторам (точкам «притягання») та їх басейнам (зонам, з яких розвиваються системи «притягуються» до атракторів (рис. 12). Таке відображення дозволяє розділити можливі стани ГПС на динамічні типи. На відміну від типів популяційних систем, виділених на основі наявності різних форм у їх складі, динамічні типи визначають і склад, і майбутню динаміку ГПС.
Рис. 12. Зони (позиції рівноваги та їх басейни) вивченого фазового простору ГПС зелених жаб
Типологія станів ГПС зелених жаб з точки зору їх екологічної стійкості. Динамічні типи ГПС, показані на рис. 12, відрізняються за своєю екологічною стійкістю. Відповідно до підходу, запропонованого А. М. Ляпуновим, у даній роботі екологічна стійкість визначається як здатність зберігати або змінювати тип системи з часом. Типи стійкості розглянутих нами систем з використанням фізичних аналогій показані на рис. 13.

Рис. 13. Фізичні аналогії типів станів ГПС зелених жаб (та інших біосистем) з точки зору їх стійкості
Порівняння різноманіття станів ГПС за результатами моделювання та за емпіричними даними. У експериментах з моделлю встановлено, що склад ГПС може залишатися постійним у трьох категоріях випадків. Перша категорія — відсутність гибридних особин. Друга категорія — передача у ГПС клональних геномів P. ridibundus і P. lessonae, а також існування особин з двома різними клональними геномами, т. е. (L)(R). Ймовірно, саме такий склад був характерним для ГПС Іськового ставища до 2000 р. Третя категорія — протилежно спрямовані впливи на склад ГПС, що взаємно нейтралізують один одного. Такі системи, що складаються з P. ridibundus і P. esculentus, зареєстровані в басейні р. Харків у м. Харкові. За даними моделювання, таке становище є стійким, якщо конкуруючі форми відрізняються за характером смертності під час онтогенезу.
Як з точки зору динамічних типів, так і з точки зору складу ГПС, модельні прогнози відповідають якісній картині, яку можна спостерігати в природних ГПС за сучасними даними про поширення ГПС зелених жаб у Північно-Донецькому центрі їх різноманіття.
Значення екологічної стійкості різних типів ГПС зелених жаб для визначення їх природоохоронної цінності. Відповідно до концепції, яку назвали раціональною природоохоронною етикою (Kravchenko, 2007; Kravchenko, Shabanov, 2009), підставами для охорони певних природних об’єктів є їх унікальність та можливість тривалого існування. Результати визначення динамічних типів ГПС можуть бути використані для оцінки їх природоохоронної цінності. Найбільшої ступені захисту заслуговують ГПС, що перебувають у стані стійкої рівноваги.
ВИСНОВКИ
1. Встановлено, що гибридогенні комплекси видів, зокрема — група зелених жаб (Pelophylax esculentus complex), утворюють геміклональні популяційні системи (ГПС). На відміну від популяцій, ГПС — це сукупності особин різних видів і їх гибридів. На відміну від спільнот і гільдій, ГПС об’єднані спільним розмноженням і передачею клональних і рекомбінантних геномів батьківських видів. Гібридні особини в ГПС можуть належати до різних кариогенетичних форм; особини батьківських видів і різні гибриди відрізняються за своїми екологічними особливостями. Існування ГПС підтримується міжвидовою гибридизацією і геміклональним успадкуванням у гибридів. Таким чином, ГПС є специфічним рівнем організації біосистем, характерним для гибридогених комплексів видів.
2. У ході багаторічних спостережень за модельною ГПС зелених жаб, що населяють Іськове ставище в околицях НПП «Гомольшанські ліси», зафіксовано трансформації її складу. Ця ГПС, яка перебувала у стабільному стані і складалася виключно з диплоїдних Pelophylax esculentus, була виведена з нього спуском ставища, опинилася на межі загибелі через те, що в ній залишився лише один клональний геном, але наразі відновлюється завдяки потраплянню в неї інших клональних геномів.
3. Розроблена при виконанні роботи концептуальна модель трансформацій ГПС Pelophylax esculentus complex враховує основні фактори динаміки цих систем. Дія сукупності біогеоценотичних і внутрішніх факторів, що впливають на трансформації ГПС, описується в цій моделі змінами параметрів життєздатності, які задаються для кожної вікової групи всіх кариогенетичних форм жаб: тривалість життя, вік першого нересту, виживаність, конкурентоспроможність, використання ресурсів, ймовірність розмноження, плодючість і ймовірність повторного розмноження в той же рік для самців. Вказання генотипів потомства від схрещування батьків, що належать до всіх передбачених у моделі кариогенетичних форм, дозволяє врахувати в моделі вплив геміклонального успадкування у гибридів.
4. Створена на основі запропонованої концептуальної моделі імітаційна модель у експериментах показала можливі шляхи трансформацій ГПС зелених жаб. Доведено, що передбачений у моделі комплекс факторів достатній для моделювання трансформацій, аналогічних тим, що були зафіксовані в природі. Встановлено, що склад кариогенетичних форм жаб, що входять до ГПС, залежить, перш за все, від особливостей геміклонального успадкування у гибридів, а кількісне співвідношення різних форм обумовлюється в першу чергу значеннями параметрів життєздатності.
5. Різні типи ГПС, виділені на підставі їх складу, можна розглядати як різні етапи процесу їх перетворень. Встановлено, що умовою стійкого стану ГПС, у склад якої входять гибриди, є передача одночасно клональних геномів обох видів і існування гибридних особин з двома клональними геномами. Це підтверджується реєстрацією таких особин у складі природних ГПС. Різноманітність станів ГПС за результатами моделювання в цілому відповідає результатам дослідження природних ГПС.
6. Аналіз можливих трансформацій ГПС з використанням імітаційної моделі дозволяє встановлювати динамічні типи ГПС. Ці типи відображають не лише особливості складу ГПС, а й перспективи її подальших трансформацій. Аттракторні динамічні типи є певними станами ГПС, яким відповідають окремі точки у фазовому просторі станів цих біосистем. Перехідним типам відповідають певні басейни (зони) у фазовому просторі; ГПС, що потрапляють у такі басейни, з часом змінюються у напрямку аттракторних типів. Описано чотири аттракторних типи екологічної стійкості (стійке, циклічне, підвижне рівновагу та загибель системи) і три перехідних типи (нестійке рівновагу, спрямовану зміну і байдужу рівновагу). Таким чином, встановлено, що динамічні типи ГПС відрізняються за екологічною стійкістю: здатністю зберігатися або змінюватися з часом.
ПОДЯКИ
Автор щиро дякує своєму науковому керівнику доценту D. A. Shabanovу. Починаючи з 2004 р., робота автора проходила у тісній співпраці з членами робочої групи з дослідження популяційної екології амфібій кафедри зоології та екології тварин ХНУ імені В. Н. Каразина. Автор дякує A. V. Korshunovу, G. A. Mazepa, E. E. Usovій, M. V. Stroilovій, D. V. Dedukha, A. A. Bondarevій, A. V. Mikhailovій, E. V. Melesko та іншим харківським батрахологам. У розвитку викладених у даній роботі ідей важливу роль зіграли колеги автора з інших наукових установ: S. N. Lytvynchuk, L. Ya. Borkin, G. A. Lada, S. Yu. Morozov‑Leonov, E. M. Pysanets. У освоєнні імітаційного моделювання автор отримала неоцінену допомогу від A. G. Kozlenko, A. A. Lutsyk, M. V. Vladimirovai та G. N. Zholtkevych. Автор щиро дякує всім цим колегам.
СПИСОК ВИКОРИСТАНИХ ДЖЕРЕЛ
Статті у профільних виданнях
1. Kravchenko M. A. Оцінка унікальності популяцій Bufo viridis Laurenti, 1768 (Amphibia, Anura) з точки зору раціональної екологічної етики / M. A. Kravchenko, D. A. Shabanov // Вісник Дніпропетровського ун-ту. Серія Біологія. Екологія — 2005. — Вип. 13. — Т. 2. — № 3/2. — С. 91 — 95. (Дисертантка брала участь у розробці концепції, відповідно до якої самостійно провела нову обробку раніше опублікованих даних і самостійно інтерпретувала результати).
2. Дослідження популяційних систем зелених жаб (Rana esculenta complex) в Харківській області: історія, сучасний стан і перспективи / D. A. Shabanov, A. I. Zynenko, A. V. Korshunov [та ін.] // Вісник Харківського нац. ун-ту ім. В. N. Kаразіна. Серія: біологія. — 2006. — Вип. 3 (№ 729). — 208 — 220. (Дисертантка зібрала частину використаних у роботі емпіричних даних, спільно з авторами інтерпретувала результати).
3. Kravchenko M. O. Етичні підстави для охорони природи: пошук раціональності / M. O. Kravchenko // Біологія та валеологія. — 2007. — Вип. 9. — С. 158–165.
4. Kravchenko M. A., Shabanov D. A. Можливі шляхи трансформації популяційних систем Pelophylax esculentus complex (Ranidae, Anura, Amphibia) / M. A. Kravchenko, D. A. Shabanov // Праці Українського герпетологічного товариства. — 2008. — № 1 — С. 15 — 20. (Дисертантка брала участь у постановці задачі, обробила емпіричний матеріал, брала участь у інтерпретації результатів).
5. Shabanov D. A., Korshunov O. V., Kravchenko M. O. Які ж зелені жаби населяють Харківську область? Термінологічний і номенклатурний аспекти проблеми / D. A. Shabanov, O. V. Korshunov, M. O. Kravchenko // Біологія та валеологія. — 2009. — Вип. 11. — С. 116 — 125. (Дисертантка брала участь у обробці літературних даних, обґрунтувала необхідність введення поняття ГПС, брала участь в інтерпретації результатів).
6. Usova O. E. Визначення демографічних параметрів зелених жаб (Pelophylax esculentus complex, Amphibia, Ranidae) для математичного моделювання їх популяційних систем / O. E. Usova, M. O. Kravchenko // Біологія та валеологія. —2010. — Вип. 12. — С. 67 — 74. (Дисертантка брала участь у постановці задачі роботи, формалізувала емпіричні дані, отримані співавтором, розробила використану імітаційну модель і брала участь в інтерпретації даних).
7. Kravchenko M. A. Моделювання трансформацій геміклональних популяційних систем зелених жаб (Pelophylax esculentus complex; Amphibia, Ranidae) за допомогою рекурентних різницевих рівнянь / M. A. Kravchenko, D. A. Shabanov // Вісник Харківського нац. ун-ту ім. В. N. Kаразіна. Серія: біологія. — 2010. — Вип. 12 (№ 920). — С. 70 — 82. (Дисертантка брала участь у постановці задачі, розробила концептуальну та імітаційну модель, провела експерименти з нею і брала участь в інтерпретації результатів).
8. Melesko E. V. Популяційна система зелених жаб (Pelophylax esculentus complex) Іськового ставища в Зміївському районі Харківської області: історія вивчення і оцінка чисельності / E. V. Melesko, A. D. Suvorova, M. A. Kravchenko [та ін.] // Біологічний вісник. — 2010. — Т. 14 — № 1. — С. 63 — 68. (Дисертантка брала участь у постановці задачі, обробці емпіричних і літературних даних, а також в інтерпретації результатів).
9. Kravchenko M. A. Дослідження стійкості геміклональних популяційних систем гибридогенного комплексу зелених жаб за допомогою імітаційного моделювання / M. A. Kravchenko, D. A. Shabanov, M. V. Vladimirova [та ін.] // Вісник Дніпропетровського університету. Біологія. Екологія. — 2011. – Вип. 19. — Т. 1. — С. 51 — 64. (Дисертантка брала участь у постановці задачі, самостійно обробила літературні дані, самостійно виконала експеримент з моделлю і брала участь у інтерпретації результатів).
10. Melesko O. V. Дослідження стану популяційної системи зелених жаб (Pelophylax esculentus complex) Іськового ставка Зміївського району Харківської області / O. V. Melesko, M. O. Kravchenko // Вісник Прикарпатського національного університету імені Василя Стефаника. Серія: біологія. — Івано‑Франківськ: Вид‑во Прикарпатського нац. ун-ту імені Василя Стефаника, 2012. — Вип. XVII. — С. 90–94.(Дисертантом була науковим керівником роботи студентки, брала участь у постановці задачі та інтерпретації результатів).
Статті в інших виданнях
11. Kravchenko M. A. Перетворення популяційних систем Rana esculenta complex як особливий тип процесів природного розвитку / M. A. Kravchenko // Питання герпетології. — Санкт‑Петербург, 2008. — С. 204 — 209.
Тези доповідей і матеріали конференцій
12. Kravchenko M. A. До розробки методів оцінки цінності популяцій з точки зору раціональної екологічної етики / M. A. Kravchenko, D. A. Shabanov // Біорізноманіття і роль зооценозу в природних і антропогенних екосистемах: Мат. III Міжнар. науков. конф. — Дніпропетровськ: Видавництво ДНУ, 2005. — С. 78 — 80. (Дисертантом самостійно обробила літературний матеріал і брала участь в інтерпретації результатів).
13. Shabanov D. A. Унікальність популяцій безхвостих амфібій як підстава для їх охорони / D. A. Shabanov, A. V. Korshunov, M. A. Kravchenko // Сучасні проблеми зоології та екології. — 2005. — С. 328 — 329. (Дисертантом брала участь у розробці викладеної концепції).
14. Korshunov A. V. Перетворення популяційних систем зелених жаб як особлива категорія процесів природного розвитку / A. V. Korshunov, M. A. Kravchenko, D. A. Shabanov // Сучасні проблеми популяційної екології: Мат. IX Міжнар. науково‑практ. еколог. конф. — Белгород: Видавництво ПОЛИТЕРРА, 2006. — С. 101 — 102. (Дисертантом брала участь у розробці концепції).
15. Kravchenko M. A. Стійкість і перетворення популяційних систем Rana esculenta complex / M. A. Kravchenko // Мат. конф. «Сучасні проблеми біологічної еволюції. До 100‑річчя Державного Дарвінського музею». — Москва: Видавництво ГДМ, 2007. — С. 113 — 115.
16. Kravchenko M. A. Ієрархія рівнів біорізноманіття на прикладі гібридогенного Rana esculenta complex / M. A. Kravchenko, D. A. Shabanov // Zoocenosis‑2007. Біорізноманіття та роль тварин в екосистемах. — Дніпропетровськ: ДНУ, 2007. — С. 381 — 383. (Дисертантом брала участь у аналізі даних).
17. Kravchenko M. A. Що нового можна дізнатися про зелені жаби за допомогою імітаційного моделювання? / M. A. Kravchenko, A. A. Lutsyk, D. A. Shabanov // Біологія: від молекули до біосфери: Мат. III Міжнар. конф. молодих вчених. — Харків, 2008. — С. 9 — 11. (Дисертантом представила результати роботи, виконаної разом зі спеціалістом з моделювання).
18. Kravchenko M. A. До розробки методів формалізованої оцінки доцільності охорони природних об’єктів / M. A. Kravchenko // «Біологія: від молекули до біосфери»: Матеріали III Міжнародної конференції молодих вчених. — Харків, 2008. — С. 442 — 443.
19. Kravchenko M. A. Математичне моделювання динаміки різних кариогенетичних форм зелених жаб у геміклональних популяційних системах / M. A. Kravchenko // Видові популяції і спільноти в антропогенними трансформованих ландшафтах: стан і методи його діагностики: Мат. XI Міжнар. науково‑практич. екологіч. конф. — Белгород, 2010. — С. 210.
20. Kravchenko M. A., Shabanov D. A. Рівні біорізноманіття Pelophylax esculentus complex / M. A. Kravchenko, D. A. Shabanov // Біорізноманіття і стале розвиток. Тези Міжнар. науково‑практ. конф. — Сімферополь: КНЦ, 2010. — С. 68 — 71. (Дисертантом брала участь у аналізі даних).
АНОТАЦІЯ
Kravchenko M. O. Екологічна стійкість популяційних систем гібридогенного комплексу зелених жаб (Pelophylax esculentus complex)». — На правах рукопису.
Дисертація на здобуття наукового ступеня кандидата біологічних наук за спеціальністю 03.00.16 — екологія. — Дніпропетровський національний університет імені Олеся Гончара. — Дніпропетровськ, 2012.
У дисертації надані оригінальні результати багаторічних досліджень складу популяційних систем гібридогенного комплексу зелених жаб (Pelophylax esculentus complex). Обґрунтовано, що геміклональні популяційні системи (ГПС) є особливим рівнем організації біосистем, що розташований між популяційним та біогеоценотичним рівнями.
Для ГПС зареєстровані процеси трансформацій — зміни складу з часом. Вони відрізняються від процесів нециклічної динаміки, характерних для інших рівнів біосистем: динаміки популяцій, сукцесій та філоценогенезу.
Розроблено концептуальну модель, що враховує основні фактори динаміки цих систем: диференціальне виживання і розмноження представників батьківських видів та їх різних гібридів, що залежать від впливу компонентів біогеоценозу, міжвидову гібридизацію і геміклональне спадкування у гібридів.
На підставі концептуальної моделі на базі MS Excel створена імітаційна модель трансформацій ГПС зелених жаб. Передбачений набір факторів виявився достатнім для того, щоб у моделі породжувалися трансформації, аналогічні зареєстрованим в природі.
Різні типи ГПС можна розглядати як окремі етапи їх перетворень. В експериментах з моделлю встановлені можливі шляхи трансформацій ГПС зелених жаб. На підставі аналізу таких переходів запропонована динамічна типологія ГПС, у якій кожному типу відповідає певний басейн у фазовому просторі станів цих біосистем. Належність ГПС до певного динамічного типу відбиває особливості її складу, дозволяє зробити припущення про напрям її трансформацій.
На підставі результатів моделювання було встановлено, що різні динамічні типи ГПС відрізняються з точки зору їх екологічної стійкості — здатності зберігатися або змінюватися з часом. Описано чотири атракторних типи екологічної стійкості (стійка, циклічна, рухлива рівновага і загибель системи) і три перехідні типи (нестійка рівновага, спрямована зміна і байдужа рівновага).
Ключові слова: екологія, Pelophylax esculentus complex, P. ridibundus, P. lessonae, P. esculentus, геміклональні популяційні системи, імітаційне моделювання, стійкість.
АНОТАЦІЯ
Kravchenko M. O. Екологічна стійкість популяційних систем гібридогенного комплексу зелених жаб (Pelophylax esculentus complex) — На правах рукопису.
Дисертація на здобуття наукового ступеня кандидата біологічних наук за спеціальністю 03.00.16. — екологія. — Дніпропетровський національний університет імені Олеся Гончара. — Дніпропетровськ, 2012.
У дисертації представлені результати багаторічних досліджень структури популяційних систем гібридогенного комплексу зелених жаб (Pelophylax esculentus complex). Обґрунтовано, що геміклональні популяційні системи (ГПС) є особливим рівнем організації біосистем, розташованим вище популяційного і нижче біогеоценотичного рівня. Від першого його відрізняє те, що популяційні системи зелених жаб включають не представників одного виду, а представників двох батьківських видів і їх гібридів. Від другого — те, що популяції батьків і гібридів об’єднані спільним розмноженням. Особливістю такого розмноження є передача без рекомбінації одного з геномів батьківських видів від гібридів до нащадків — геміклональне спадкування.
Для ГПС зареєстровані процеси трансформацій — зміни складу з часом. Вони відрізняються від процесів нециклічної динаміки, характерних для інших рівнів біосистем: динаміки популяцій, сукцесій і філоценогенезу.
Для формалізації знань і проведення подальших досліджень трансформації ГПС розроблена концептуальна модель, що враховує основні фактори динаміки цих систем: диференціальне виживання і розмноження представників батьківських видів і їх різних гібридів, що залежать від впливу компонентів біогеоценозу, міжвидову гібридизацію і геміклональне спадкування у гібридів.
На підставі концептуальної моделі на базі Microsoft Excel створена імітаційна модель трансформацій ГПС зелених жаб. Передбачений набір факторів виявився достатнім для того, щоб у моделі породжувалися трансформації, аналогічні зареєстрованим у природі. Це свідчить про несуперечливість концептуальної моделі і достатність передбачених у ній факторів динаміки ГПС. Фактори зовнішнього середовища не розглядалися окремо, а оцінювався їх сукупний вплив на показники життєздатності жаб різних каріогенетичних і вікових форм.
Різні типи ГПС, виділені на підставі каріогенетичного складу особин і такі, що є вузловими точками в концептуальній моделі, можна розглядати як окремі етапи їх перетворень. В експериментах з моделлю підтверджені і уточнені можливі шляхи трансформацій ГПС зелених жаб — логічного переходу від одного типу до іншого. На підставі аналізу таких переходів була запропонована динамічна типологія ГПС. Кожному динамічному типу відповідає певний басейн у фазовому просторі станів цих біосистем. Належність ГПС до певного динамічного типу відображає не лише особливості її складу, але і дозволяє зробити припущення про її подальші трансформації, оцінити ймовірність загибелі або нормального функціонування ГПС у певному місцеперебуванні.
На підставі результатів моделювання було встановлено, що різні динамічні типи ГПС відрізняються з точки зору їх екологічної стійкості — здатності зберігатися або змінюватися з часом. Описано чотири аттракторних типи екологічної стійкості (стійка, циклічна, рухлива рівновага і загибель системи) і три перехідних типи (нестійка рівновага, спрямована зміна і байдужа рівновага).
Ключові слова: екологія, Pelophylax esculentus complex, P. ridibundus, P. lessonae, P. esculentus, геміклональні популяційні системи, імітаційне моделювання, стійкість.
ANNOTATION
Kravchenko M. O. Ecological sustainability of population systems in the water frogs of the Pelophylax esculentus hybridogenetic complex. — Manuscript.
Dissertation for a degree of Candidate of Biological Sciences by speciality 03.00.16. – ecology. — Oles’ Gonchar Dnipropetrovsk National University. — Dnipropetrovsk, 2012.
Original results of the long‑term studies of population systems of water frogs hybridogenetic complex (Pelophylax esculentus complex) are provided. Present work justifies that hemiclonal population systems (HPS) represent a special level of biosystems, which bridges population and biogeocenotic levels.
We registered transformation (i.e., change their composition over time) of the HPS. These processes differ from noncyclic dynamics processes characteristic of other levels of biosystems: population dynamics, succession or phylocoenogenesis.
Further, we developed a conceptual model that takes the main factors of the HPS dynamics into account. This model considers differential survival and reproduction of the individuals belonging to the parental species and their various hybrids, which depend on the impact of components biogeocoenosis, as well as interspecific hybridization and hemiclonal inheritance in hybrids.
Ultimately, on the basis of that conceptual model we developed an original simulation model (MS Excel) to trace transformations of the water frogs HPS. The provided set of factors appeared to be sufficient to observe the model transformations similar to the processes registered in nature. Different types of HPS can be regarded as separate stages of their transformation sequence. Based on subsequent analysis of these models transitions we propose dynamic typology of the HPS. Each type has a corresponding dynamic pool in the phase space of states of a HPS. Belonging to a particular HPS dynamic type reflects the peculiarities of its structure and allows to make assumptions about its future transformation direction.
Based on the simulation results we found that different types of dynamic HPS differed in terms of their ecological sustainability – the ability to persist or change over time. Four attractor types of ecological stability (stable, cyclic, agile and loss of balance) and three types of transient (unstable equilibrium, directional change and indifferent equilibrium) were described.
Key words: ecology, Pelophylax esculentus complex, P. ridibundus, P. lessonae, P. esculentus, hemiclonal population systems, simulation modeling, stability.