Від хаосу спостережень до динамічної типології: обговорення на прикладі популяційних систем зелених жаб. Колонка для Компьютерры #113
Не «подпирайте» свои суждения ссылками на волю Творца. Давайте сами, силами нашего разума наблюдать, экспериментировать, моделировать, строить прогнозы и выбирать желаемое для нас состояние!
{
"title": "Ноосфера: рациональний прогноз, сциентистське візіонерство чи розхоже заклинання шарлатанів від науки?",
"summary": "Від хаосу спостережень до динамічної типології: обговорення на прикладі популяційних систем зелених жаб",
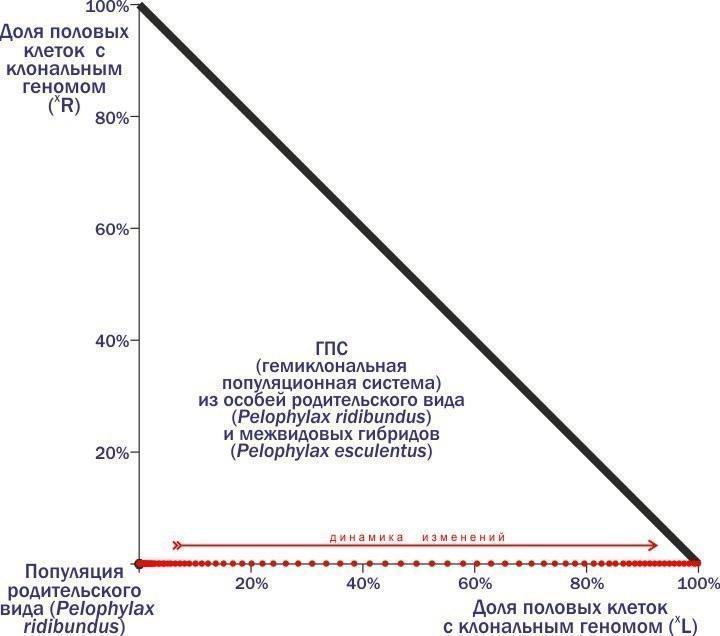
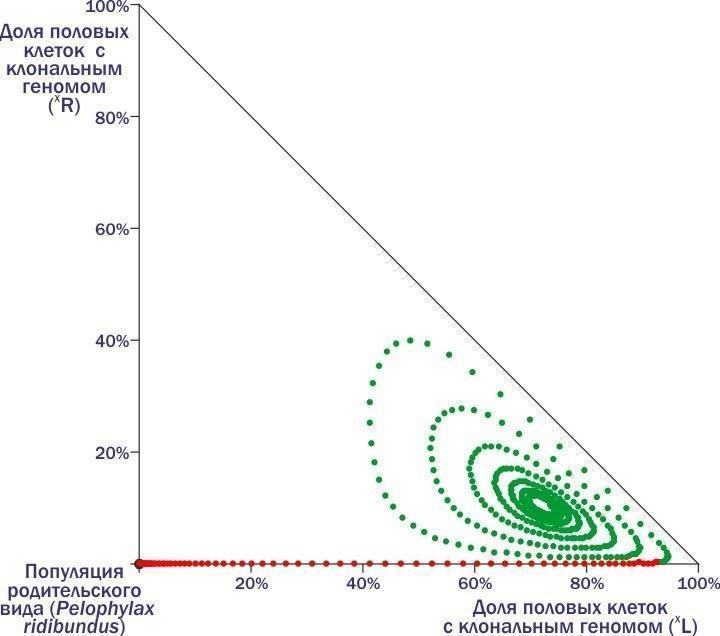
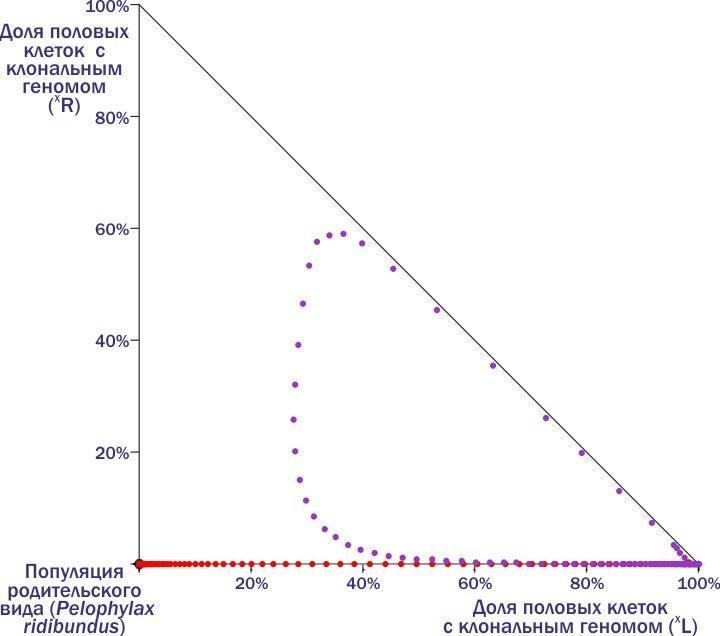
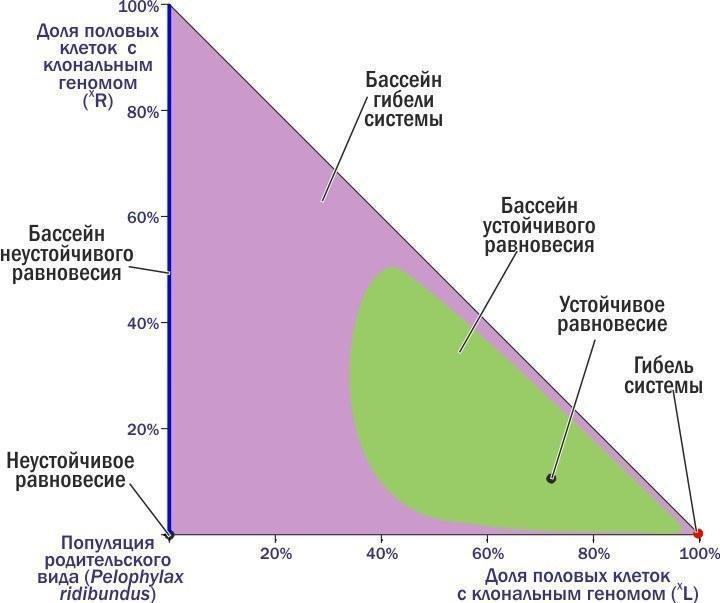
"body": "←\nДмитро Шабанов\n→\n\nНоосфера: рациональний прогноз, сциентистське візіонерство чи розхоже заклинання шарлатанів від науки?\nВід хаосу спостережень до динамічної типології: обговорення на прикладі популяційних систем зелених жаб\nО чому можна судити, вимірюючи флуктуючу асиметрію?\n\nКолонка для Комп'ютерри #112\nКолонка для Комп'ютерри #113\nКолонка для Комп'ютерри #114\n\nВикидавши чергову колонку, я раз за разом заглядаю на сайт КТ: як сприйняли? Частіше спочатку з'являються коментарі від тих, хто зрозумів, а потім підтягуються пропагандисти креаціонізму і сакральних традицій, а також професійні сперечальники. Реагувати? Надо зрозуміти, для кого я пишу колонки. Єстественно, для читачів (ну і, звичайно, трохи для себе). Значить, надо відповідати. Як це робити, не утопая в гримзі під колонками? Тут я спробую розповісти про одну роботу з моїм учасством. Чи зможе вона стати аргументом, що дозволить проілюструвати підхід, який здається мені правильним?\nЯ розповім про фрагмент роботи, яку три тижні тому захистила як кандидатську Марина Кравченко. Я — науковий керівник цієї роботи. Головні (з моєї точки зору) її результати опубліковані з моїм учасством (тут — сама модель, а тут — результати моделювання, про які я розповім).\nТе, що я зараз буду описувати, не є суттю дисертації Марини з точки зору формальних вимог. Зато це та її частина, з якої, як мені здається, слідують певні світоглядні висновки.\nМарина Кравченко вивчала трансформації популяційних зелених жаб. Динаміка цих систем (ми називаємо їх ГПС, геміклональними популяційними системами) практично не вивчена. Чому? Зараз поясню.\nНачну з твердження, що основні властивості добре відомих нам біосистем (організмів, популяцій, спільнот) є наслідком передачі генетичної інформації з рекомбінацією (перебором сочетань).\nГоловний механізм еволюції — відбір організмів, покоління за поколінням, що проходять непростий шлях самоорганізації під впливом спадкової програми. Одним з важливих способів підвищення ефективності еволюції таких спадкових програм стало запліднення — злиття клітин двох різних організмів. Кожна з цих клітин несе спадкові задатки, відібрані в попередніх поколіннях. Але у клітини, яка отримується від їх злиття, набір генетичної інформації виявиться удвоєним. Тому невід'ємною частиною життєвих циклів із заплідненням стає мейоз — клітинне деління, при якому кількість генетичної інформації скорочується вдвічі. Чергування в життєвому циклі мейозу і запліднення підтримує кількість передаваної генетичної інформації відносно постійним.\nМейоз і запліднення підтримують рекомбінацію генетичного матеріалу, прискорюючи виведення пристосувань в еволюції. Генетично унікальні організми, що виробляють генетично унікальні статеві клітини, популяції з спільним генофондом, міжвидова ізоляція і особлива еволюційна доля кожного виду — наслідки цього процесу.\nСередньоєвропейські зелені жаби (і кілька інших гібридогенних комплексів видів) внесли принципову зміну в базовий процес. В цих комплексах утворюються міжвидові гібриди, які передають майбутнім поколінням комплекс генетичної інформації, отриманої від одного з батьківських видів, як єдине ціле, без рекомбінації. І в силу цього властивості всіх ієрархічних структур, існування яких підтримується розмноженням з рекомбінацією, змінюються.\nОрганізми починають виробляти однакові, клональні статеві клітини. Гібриди належать до полуклонів з функціонально різними геномами (комплексами спадкової інформації): одні з цих геномів передаються з покоління в покоління клонально, інші — використовуються протягом життя особини, а потім безслідно знищуються. Кордони між популяціями розмиті, адже в спільному розмноженні беруть участь і представники батьківських видів. Сплошь і рядя порушення базового способу передачі генетичної інформації призводить до появи незвичайних особин. Це поліплоїди з збільшеним кільком геномів, а також часткові міжвидові рекомбінанти. Між батьківськими видами відбувається обмін генетичною інформацією, причому внедрення чужорідних генів торкається і тих популяцій, які не беруть участь у гібридизації.\nЦікавіша для популяційного еколога проблема — вияснення того, як підтримується стійкість популяційних систем, в рамках яких відбуваються такі чудеса. Як досліджувати таку проблему?\nОчевидно, основа для такого відповіді — збирання емпіричних даних. І нами, і іншими дослідниками встановлено, що склад ГПС може з часом змінюватися. Деякі з таких змін ми з нашими студентами спостерігаємо в полі. В будь-якому випадку такі трансформації — надзвичайно тривалий процес, а їх відстежування — важка робота. Про те, щоб за допомогою польових досліджень за розумний час вивчити закономірності цих трансформацій, розглянути сукупність можливих станів і логіку їх перетворення, говорити не приходиться. Що ж робити?\nЗастосовувати моделювання. Причому не аналітичне, в ході якого ми використовуємо заздалегідь встановлені залежності, що описують результати цікавих нас процесів, а імітативне. В ході імітативного моделювання ми намагаємось хоча б приблизно описати етапи вивчених перетворень, визначаючи, в якому напрямку змінюється цікавить нас система.\nТак ми і зробили. Описали річний цикл зелених жаб, виділили в ньому етапи, що впливають на склад ГПС, задали залежності, що характеризують такі зміни. На основі власних і літературних даних припустили, який склад статевих клітин характерний для кожної з форм гібридних жаб. Отримали конструкцію з рівнянь, що описують цікавить нас параметри.\nЦя конструкція виявилася реалізованою в двох різних моделях. Ми з Мариной зробили і використали модель на різностних рівняннях, написану в Excel, а наші колеги-математики зібрали більш потужну реалізацію на основі професійних мов і пакетів. Професійна модель значно перевершує нашу, тренувальну, по потужності, але помітно поступається їй по зручності використання і перебудові. Марина захистила дисертацію, зроблену на тренувальній моделі, але при цьому ми з колегами-математиками продовжували працювати над моделлю «великої».\nРозкажу про експерименти з тією моделлю, яку назвав «тренувальною», Excel-евською. В ній прописаний механізм розмноження, росту і конкуренції жаб з різними наборами геномів. Цей механізм носить вполне «загальний» характер. Але крім того, в моделі є блок, який забезпечує можливість задавати, які статеві клітини виробляють представники того або іншого генотипу. Тут-то і можна передбачити полуклональне спадкування, характерне для зелених жаб.\nА тепер, щоб пояснити основну ідею колонки, мені доведеться розповісти про декілька експериментів з моделлю і пояснити логіку її перебудови. Почну з прикладу, який докладно розбирала в колонці про Невидиму Ногу. В прикладах, які я привожу, букви, L і R — позначення геномів (кожний з 13 хромосом). Геноми, позначення яких не беруться в дужки, — звичайні, рекомбінантні, такі, як у кожного з нас (той комплекс спадкової інформації, що ми передаємо дітям, є відносно випадковою сумішшю геномів, які ми отримали від батька і матері). В дужки беруться клональні геноми, які передаються як єдине ціле.\nРозглянемо популяцію батьківського виду, всі скрещування в якій такі:\n♀RR × ♂RR → ♀RR : ♂RR.\nТут відбувається приблизно те саме, що і в популяціях людей (і система визначення статі та ж): особини певного виду виробляють особин того ж виду. Але уявімо собі, що в таку популяцію потрапляє хоча б одна гібридна особина, що несе клональний геном іншого виду - наприклад, ♀(L)R. Заключенням геному (L) в дужки ми вказуємо, що він потрапляє у всі статеві клітини. Раз так, усе потомство від скрещування такого гібрида з особинами батьківського виду буде гібридним:\n♀(L)R × ♂RR → ♀(L)R : ♂(L)R.\nТепер ми вже маємо справу не з популяцією, а з ГПС — геміклональною популяційною системою, адже в ній передаються з покоління в покоління клональні і рекомбінантні геноми, що належать до різних видів. При скрещиванні самця-гібрида з самкою батьківських видів також виникне тільки гібридне потомство (оскільки геном жіночий, усе потомство буде самками):\n♀RR × ♂(L)R → ♀(L)R.\nНарешті, ще однією особливістю гібридів є те, що потомки від їх скрещування будуть нежизнеспособними:\n♀(L)R × ♂(L)R → ♀(L)(L) → †.\nТаким чином, скрещивание «батьків» (особин батьківського виду) породжує «батьків». Від скрещування «батьків» і гібридів виникають тільки гібриди, внаслідок чого доля останніх зростає. Скрещивания гібридів не впливають на склад майбутніх поколінь, так як потомство від них гине.\nРезультатом є те, що доля гібридів в такій ГПС зростає, поки в ній не залишаються тільки вони. Після цього така система гине!\nБагато людей дивується, що для загибелі батьківської популяції достатньо потрапляння в неї однієї-єдиної гібридної особини. Тут треба думати не про внесок в початковий склад представників різних форм, а про те, чи присутня в системі «інфекція», тобто геном, розповсюдження якого веде до катастрофи.\nДавайте подивимся, яку траєкторію опише ГПС на фазовій площині (положення на якій відображає склад системи).\n