
From the chaos of observations to dynamic typology: a discussion using the example of green frog population systems. Column for Kompyuterra #113
Do not «prop up» your judgments with references to the will of the Creator. Let us ourselves, by the powers of our reason, observe, experiment, model, make forecasts, and choose the state we desire!
{"title": "Noosphere: Rational Forecast, Scientistic Vision, or Common Incantation of Charlatans of Science?", "summary": "From the Chaos of Observations to Dynamic Typology: A Discussion Using the Example of Green Frog Populations", "body": "← Dmitry Shabanov → Noosphere: Rational Forecast, Scientistic Vision, or Common Incantation of Charlatans of Science? From the Chaos of Observations to Dynamic Typology: A Discussion Using the Example of Green Frog Populations What can be inferred by measuring fluctuating asymmetry? Column for Kompyuterra #112 Column for Kompyuterra #113 Column for Kompyuterra #114 After publishing another column, I repeatedly check the KT website: how was it received? Usually, comments from those who understood appear first, followed by proponents of creationism and sacred traditions, as well as professional debaters. Should I respond? I need to understand who I am writing these columns for. Naturally, for the readers (and, of course, a little for myself). Therefore, I must respond. How to do this without getting bogged down in the mud under the columns? Here I will try to describe one work in which I participated. Can it serve as an argument that illustrates an approach that seems correct to me? I will describe a fragment of the work that Marina Kravchenko defended as a candidate dissertation three weeks ago. I am the scientific supervisor of this work. Its main results (from my point of view) have been published with my participation (here is the model itself, and here are the simulation results, which I will discuss). What I am about to describe is not the essence of Marina's dissertation in terms of formal requirements. However, it is the part of it from which, it seems to me, certain worldview conclusions follow. Marina Kravchenko studied the transformations of green frog populations. The dynamics of these systems (we call them HPS, hemiclonal population systems) are practically unstudied. Why? I will explain now. I will start with the assertion that the main properties of well-known biosystems (organisms, populations, communities) are the result of the transmission of genetic information with recombination (shuffling of combinations). The main mechanism of evolution is the selection of organisms, generation after generation, undergoing a difficult path of self-organization under the influence of a hereditary program. One of the important ways to increase the efficiency of evolution of such hereditary programs was fertilization – the fusion of cells from two different organisms. Each of these cells carries hereditary predispositions selected in previous generations. But the cell resulting from their fusion will have a doubled set of genetic information. Therefore, an integral part of life cycles with fertilization is meiosis – cell division in which the amount of genetic information is halved. The alternation of meiosis and fertilization in the life cycle maintains the amount of transmitted genetic information relatively constant. Meiosis and fertilization maintain the recombination of genetic material, accelerating the evolution of adaptations. Genetically unique organisms that produce genetically unique gametes, populations with a common gene pool, interspecific isolation, and the unique evolutionary fate of each species are consequences of this process. The Central European green frogs (and several other hybridogenic species complexes) introduced a fundamental change to this basic process. In these complexes, interspecific hybrids are formed that transmit to future generations a complex of genetic information obtained from one of the parental species as a single unit, without recombination. As a result, the properties of all hierarchical structures whose existence is maintained by reproduction with recombination change. Organisms begin to produce identical, clonal gametes. Hybrids belong to semi-clones with functionally different genomes (hereditary information complexes): some of these genomes are transmitted from generation to generation clonally, while others are used during the individual's life and then destroyed without a trace. The boundaries between populations are blurred, as representatives of the parental species also participate in reproduction. Widespread violations of the basic mode of genetic information transmission lead to the appearance of unusual individuals. These are polyploids with an increased number of genomes, as well as partial interspecific recombinants. Exchange of genetic information occurs between parental species, and the introduction of foreign genes affects even populations that do not participate in hybridization. A more interesting problem for the population ecologist is to determine how the stability of population systems, within which such wonders occur, is maintained. How to study such a problem? Obviously, the basis for such an answer is the collection of empirical data. We and other researchers have established that the composition of HPS can change over time. We and our students observe some of these changes in the field. In any case, such transformations are an extremely long process, and tracking them is difficult work. It is out of the question to study the regularities of these transformations using field research within a reasonable time, to consider the set of possible states and the logic of their transformation. So what should be done? Use modeling. And not analytical modeling, during which we use pre-established dependencies describing the results of interesting processes, but simulation modeling. During simulation modeling, we try to describe, at least approximately, the stages of the studied transformations, determining in which direction the system of interest changes. This is what we did. We described the annual cycle of green frogs, identified the stages that affect the composition of HPS, and set the dependencies characterizing these changes. Based on our own and literature data, we assumed the composition of gametes characteristic of each form of hybrid frogs. We obtained a system of equations describing the parameters of interest. This system turned out to be implemented in two different models. Marina and I created and used a model with difference equations written in Excel, and our mathematician colleagues built a more powerful implementation based on professional languages and packages. The professional model significantly surpasses our training model in power, but is noticeably inferior to it in ease of use and modification. Marina defended her dissertation based on the training model, but at the same time, my colleagues and I continued to work on the 'large' model. I will describe experiments with the model we called the 'training' model, the Excel-based one. It describes the mechanism of reproduction, growth, and competition of frogs with different genome sets. This mechanism is quite 'general'. However, in addition, the model includes a block that allows specifying which gametes are produced by representatives of a particular genotype. This is where semi-clonal inheritance, characteristic of green frogs, can be simulated. Now, to explain the main idea of the column, I will have to describe several experiments with the model and explain the logic of its modification. I will start with an example that was discussed in detail in the column about the Invisible Leg. In the examples I provide, the letters L and R denote genomes (each with 13 chromosomes). Genomes not enclosed in parentheses are normal, recombinant, like those of any of us (the hereditary information complex we pass on to our children is a relatively random mixture of genomes we received from our father and mother). Genomes enclosed in parentheses are clonal, transmitted as a whole. Consider a population of a parental species where all crosses are as follows: ♀RR × ♂RR → ♀RR : ♂RR. Here, approximately the same thing happens as in human populations (and the sex determination system is the same): individuals of a certain species produce individuals of the same species. But imagine that at least one hybrid individual carrying a clonal genome of another species enters such a population – for example, ♀(L)R. By enclosing the genome (L) in parentheses, we indicate that it is transmitted in all gametes. If so, all offspring from the cross of such a hybrid with individuals of the parental species will be hybrid: ♀(L)R × ♂RR → ♀(L)R : ♂(L)R. Now we are dealing not with a population, but with an HPS – a hemiclonal population system, because clonal and recombinant genomes belonging to different species are transmitted from generation to generation. When a male hybrid crosses with a female of the parental species, only hybrid offspring will also arise (since the female genome is involved, all offspring will be females): ♀RR × ♂(L)R → ♀(L)R. Finally, another peculiarity of hybrids is that the offspring from their crosses will be non-viable: ♀(L)R × ♂(L)R → ♀(L)(L) → †. Thus, the crossing of 'parents' (individuals of the parental species) produces 'parents'. Crossing 'parents' and hybrids produces only hybrids, causing the proportion of the latter to increase. Crossings of hybrids do not affect the composition of future generations, as offspring from them die. The result is that the proportion of hybrids in such an HPS increases until only they remain. After that, such a system dies! Many people are surprised that the introduction of just one hybrid individual into a parental population is enough for its demise. Here, one should not think about the contribution to the initial composition of representatives of different forms, but about whether the 'infection' is present in the system, i.e., a genome whose spread leads to catastrophe. Let's see what trajectory the HPS will follow on the phase plane (positions on which reflect the composition of the system). Look. In the corner is the population of the parental species from which we started the experiment.
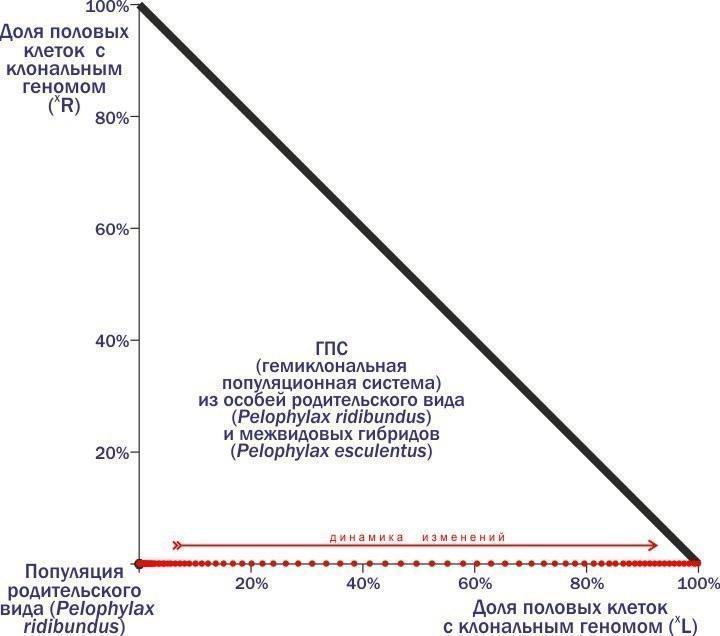
The axes represent the proportions of clonal genomes. In the example considered, we add one individual (L)R to a population of RR individuals. Due to the mechanism we just described, the proportion of individuals with the (L) genome in the considered HPS begins to increase. Its trajectory is marked by red dots; the direction of dynamics is shown by an arrow. It ends with the death of the HPS when it reaches the right corner: if 100% of its composition consists of (L) genome carriers, all crosses lead to the appearance of non-viable offspring. When individuals carrying the (R) genome are added to the parental species population, nothing interesting happens. Crossing such hybrids with individuals of the parental species yields 'parents'. This means that the parental species population is in a stable equilibrium with respect to the introduction of (R) genomes and unstable with respect to (L) genomes. What happens if the (R) genome enters a population where most individuals are carriers of the (L) genome?
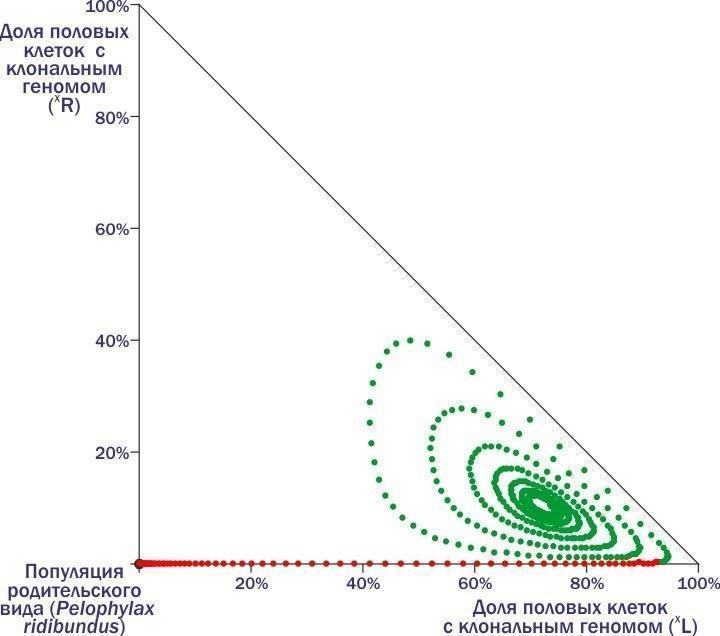
In the place where the red dots turn green, one individual with the clonal genome (R) is added to the HPS. Describing loops, the model HPS moves towards a state of stable equilibrium – somewhere in the center, towards which its trajectory tends. And to reach it, the HPS must receive the (R) genome within a fairly specific time frame. Earlier – it will get lost. Later – although it will cause sharp changes in composition, it will still not save the system from death.
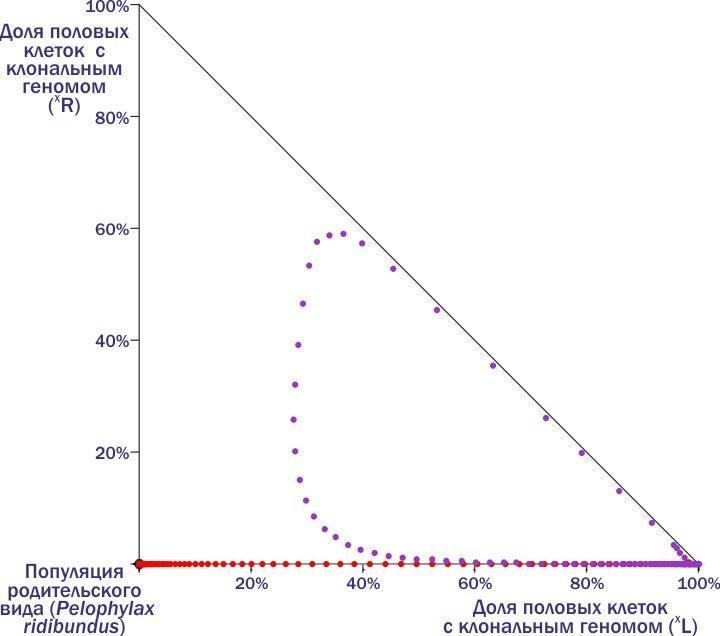
Here, at the moment when the red dots turn purple, a carrier (R) enters the HPS. The system's trajectory describes a loop... and still leads to its death. Comparing the two previous figures, we can see that outside the widest green loop in the previous figure and inside the purple loop in the last one lies the boundary between the regions from which the HPS enters two different final states – stable equilibrium and death. These states correspond to attractor (pulling the dynamic system towards them) points on the phase plane. Thus, we can divide the phase plane into basins of different attractor states! Until now, different HPS have been classified by their composition.
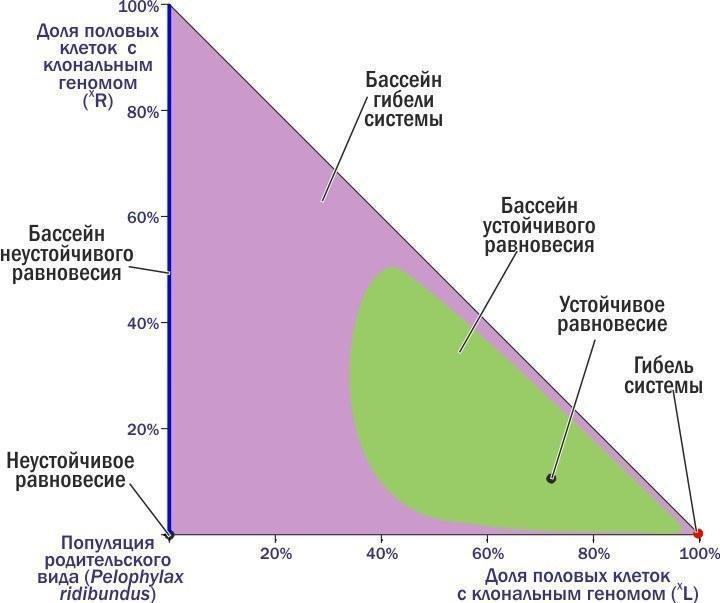
The approach described here allows us to identify dynamic types that are characterized not only by a specific composition but also by a common expected fate. These types are basins in the phase space of the states of the systems of interest. Look at what we have done. We have not introduced any external criteria of 'good' or 'bad' into the discussion of the dynamics of the system of interest. We have simply established that some states systematically transition into others, and divided their totality into groups. Now we can hope that the dynamic types we have identified reflect the actual characteristics of the studied systems. We can choose which states are optimal for us. The next step is to understand how to transition from observed states to desired ones (if they appear). Debaters have argued with me in the comments under previous columns that one cannot even distinguish a healthy person from a sick one without knowing God's will, and even less can one understand what biosphere state we should strive for. The example provided here shows, I hope, that the typology of states of a complex system can be fully constructed without reference to supernatural goal-setting. Yes, my example relates not only to HPS. The typologies of many 'everyday' dynamic systems that we use are obtained through procedures that are essentially similar. The only difference is that the analysis