
Подкласс Lepidosauromorpha
Підклас Lepidosauromorpha нині налічує більше 7 тисяч видів ящірок і змій і є другою після птахів найбагатшою видами групою тетрапод. Термін Lepidosauromorpha ввів в ужиток М. Бентон у 1983 р. Представників лепідозаврів, як і примітивних архозавроморф, вважають примітивними діапсидами: наразі Archosauromorpha + Lepidosauromorpha складають Sauria і Sauria + Younginiformes складають Neodiapsida (Laurin, 1991). Імовірно, Lepidosauromorpha відділилися від Archosauromorpha в середині пермі.
Лепідозаври походять від ящіркоподібних лепідозавроморфних таксонів, які процвітали під час пізньої пермі й раннього тріасу. Під час раннього тріасу ці різноманітні форми породили такі спеціалізовані форми як kuehneosaurs, завроптеригії (можливо) і справжні лепідозаври. Усі ці рептилії поширювалися, займаючи екологічні ніші, що звільнилися наприкінці пермського вимирання, раніше займані звіроподібними рептиліями, що переважали раніше. Лепідозаври виникли як невелика група з акцентом на невеликі ніші, які не були заповнені архозаврами.
Характерними рисами лепідозавроморф є:
- незграбна постава зі звивистими рухами (примітивна характеристика, властива початковим тетраподам);
- акцент на бічних хвилеподібних вигинах хребта;
- жорстка грудна клітка, велика грудина;
- немає двоногої спеціалізації;
- зап'ястки й кисті, а також шийні хребці немодифіковані (за аналогією з примітивними діапсидами);
- перекривна луска.
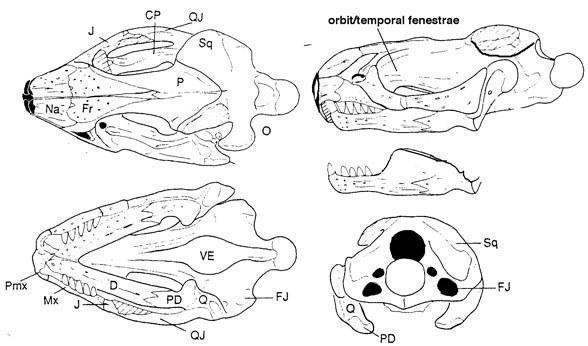
Прогрес лепідозаврів можна пояснити еволюцією стрептостилії (передньо-заднього руху квадратної кістки) і мобільності щелеп і черепа в цілому. Стрептостилія в лепідозаврів досягається шляхом часткової або повної втрати потужної нижньо-скроневої дуги. Діапазон змін нижньо-скроневої дуги діапсид і, зокрема, лепідозаврів відображений на малюнку:
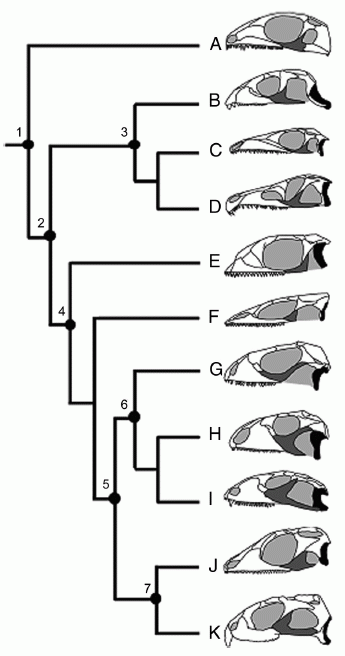
Рис. 1. Спрощена філогенія діапсидних рептилій, що показує варіації в розвитку нижньої скроневої дуги.
Чорним показана квадратна кістка, сірим - вилична. (A) діапсида Petrolacosaurus; (B) ранній ринхозавр Mesosuchus; (C) архозавроморф Prolacerta; (D) архозавроформ Proterosuchus; (E) лепідозавроформ Kuehneosaurus; (F) ранній лепідозавр Marmoretta; (G) збережене лускате Iguana, що відображає більш-менш неспеціалізовану форму; (H) величезний травоїдний крейдовий сцинкоморф Polyglyphanodon, (I) пізньокрейдова ящірка Tianyusaurus, унікальна серед лускатих з огляду на акінетичну ринхоцефальну квадратну кістку, (J) ринхоцефал Gephyrosaurus; і (K) нині живий ринхоцефал Sphenodon. Помітки: 1, Diapsida, примітивна нижньо-скронева дуга, фіксована квадратна кістка, міцний епіптеригоїд; 2, Sauria, немає нижньої скроневої дуги, фіксована квадратна кістка й епіптеригоїд; 3, Archosauromorpha, немає нижньої скроневої дуги в базальних таксонах, але розвинена в Rhynchosaurs і Archosaurs; 4, Lepidosauriformes і 5, Lepidosauria, немає нижньої скроневої дуги в базальних таксонах, квадратна кістка, епіптеригоїд зростаються з птеригоїдом, велика луската кістка; 6, Squamata, перекривання квадратної кістки й птеригоїда зменшено, епіптеригоїд пов'язаний з птеригоїдом, нижня скронева дуга розвинена тільки в Tianyusaurus only; і 7, Rhynchocephalia, немає нижньої скроневої дуги в базальних таксонах, розвинена тільки в деяких нащадків, квадратна кістка, птеригоїд і епіптеригоїд міцно зашиті. З Moazen et al, 2009
До складу Lepidosauromorpha входить інфраклас Лепідозаври (Lepidosauria) з надрядами Лускаті (Squamata) і Дзьобоголові (Rhynchocephalia або Sphenodontida). Сфенодонти були дуже поширені й успішні з кінця тріасу до пізньої юри, але поступово зменшилися протягом крейдового й третинного періодів. У той же час, лускаті, які були рідкісними або невідомими до середньої юри, досі є вельми успішними після прогресування в крейді й кайнозої. Різниця між цими групами лепідозаврів полягає в тому, що вони мають різні адаптивні стратегії, в однієї групи розвиваючи сильний, нерухомий, пристосований для потужного укусу череп, а в іншої - череп зі зниженою силою й високою мобільністю. Останнє пристосування отримали змії, які можуть відкрити рот надзвичайно широко для того, щоб проковтнути велику здобич.
Викопні знахідки лепідозавроморф.
Найбільш ранні передбачувані лепідозаври - пізньопермські Lanthanolania (Росія, Modesto and Reisz, 2002) і Saurosternon (Південна Африка, Carroll, 1975).
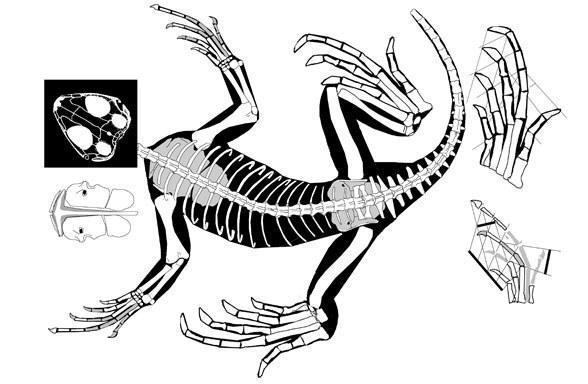
Saurosternon bainii (Huxley 1868)
Paliguana (ранній тріас, Південна Африка, Carroll, 1975) представлений одним пошкодженим черепом з величезною квадратною кісткою, що відповідає ознакам лепідозавроморфів. Paliguana відносять до Eolacertilia, куди також належить група Marmoretta.
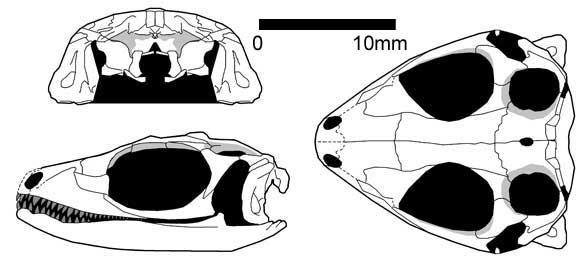
Paliguana whitei (Broom 1903)
Крім цього, останки, що відносяться до ранньотріасових лепідозавроморф, були знайдені в Польщі (Czatkowice, Borsuk-Białynicka
et al., 1999). Ці відкладення сформувалися в посушливому середовищі з локалізованими водоймами, фауна включала риб, темноспондилів, амфібій, проколофонів і окремих архозавроморфів.
Kuehneosauria. Кюнеозаври були спеціалізованими, довгоребристими планерами, відомими з раннього тріасу Польщі й пізнього тріасу Англії й Північної Америки. Цікавою особливістю кюнеозаврид були незвично довгі літальні перетинки, принаймні вдвічі більші за їх розміри.
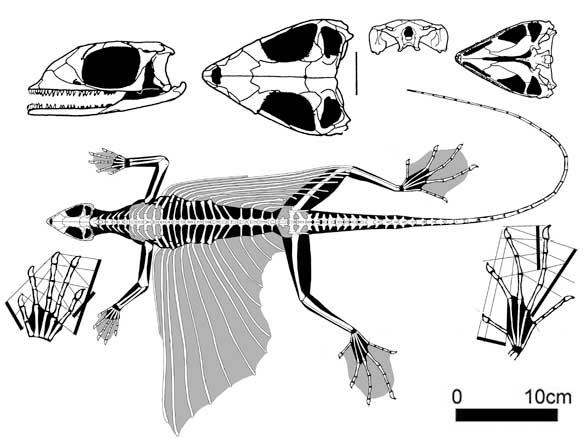
Kuehneosaurus latus (Robinson 1962), пізній тріас, Англія
Кюнеозаври були здатні планувати на шкірній перетинці, натягнутій на 10-11 парах рухомих ребер, зчленованих з платицельними хребцями. Особливо довгі ребра мав північноамериканський Icarosaurus siefkeri з черепом завдовжки всього 25 мм, «літні» ребра в нього були чітко вигнуті:

Icarosaurus siefkeri Colbert, 1966
Кюнеозаври жили на невеликих вологих прибережних морських островах разом з птерозаврами, арозавроформами, ринхоцефалами й рідкісними ссавцями (Robinson, 1962; Fraser, 1994).
До інших мезозойських лепідозавроморф, що не належать до лепідозаврів, відносяться середньотріасовий Coartaredens (Англія, Spencer and Storrs, 2002) і Megachirella (Італія, Renesto and Posenato, 2003); ранньоюрський Tamaulipasaurus (Мексика, Clark and Hernandez, 1994). Філогенетична спорідненість Megachirella і Tamaulipasaurus залишається невирішеною.

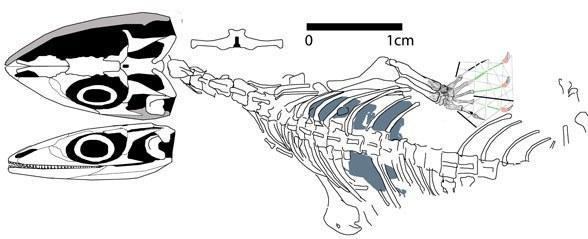
Викопна й реконструйована Megachirella wachtleri (Renesto and Posenato 2003).
Tamaulipasaurus morenoi (Clark and Hernandez 1994).
Надряд Rhynchocephalia (Sphenodonta)
Дзьобоголові - справжні діапсиди некрупних розмірів. Викопні рештки ринхоцефалів відносяться до більш ніж 40 таксонів у діапазоні від пізнього тріасу до нинішнього часу. Географічне поширення: Європа, Північна й Південна Америка, Китай, Індія, Марокко, Південна Африка й Нова Зеландія.
Справжні ринхоцефали з'являються в нижньому тріасі Південної Африки (Palacrodon) і, можливо, Східної Європи, але досягають розквіту у верхньому тріасі Західної Європи, звідки описують більше десятка родів сфенодонтид. Одна форма описана з Кенії. З нижньої юри Індії за уламками щелеп описано два роди ринхоцефалів. У середній юрі Мексики з'являються вже сфенодонти, близькі до сучасної гатерії, у тому числі карликовий Zapatodon [Reynoso, Clark, 1998] і Cynosphenodon. Сфенодонтиди відомі з верхньої юри Північної Америки (три роди) і Західної Європи. З крейди відомі тільки деякі сфенодонти. У кайнозої викопні ринхоцефали невідомі, але нині існує новозеландський ринхоцефал гатерія - Sphenodon punctatus.

Sphenodon punctatus Evans, 1980
В екологічному відношенні мезозойські дзьобоголові були різноманітні. Вони включали довготілих морських плавців (плеврозаври, сафеозаври, Керролл and Wild, 1994), грацильних бігунів (Homoeosaurus, Cocude-Мішель, 1963), броньованих Pamizinsaurus (Reynoso, 1997) і групу великотілих рептилій з копитоподібними кігтьовими фалангами (Priosphenodon, і Apesteguia Novas, 2003). Серед них були комахоїдні форми (Gephryosaurus; Diphydontosaurus), опортуністичні "хижаки" (Sphenodon, Dawbin, 1962; Cree та ін., 1999), імовірно отруйні хижаки (Sphenovipera, Reynoso, 2005, відомий із середньої юри Мексики), а також спеціалізовані травоїдні тварини (наприклад, Toxolophosaurus, Priosphenodon, Apesteguía і Novas, 2003). Незважаючи на те, що деякі їх характеристики менш просунуті, ніж у ящірок, деякі примітивні їх риси є вторинними. Сфенодонти можуть залишатися активними при нижчих температурах, ніж ящірки. Також для дзьобоголових характерний довгий репродуктивний цикл.

Рис. 2. Сфенодонти
Характерні особливості будови сфенодонтів: акродонтні зуби з уповільненим заміщенням, повна нижня скронева дуга, нерухома квадратна кістка (монімостилія), укорочені шийні хребці, хвостові хребці часто поділені надвоє автотомальною септою, що полегшує довільну автотомію (відрив хвоста). При живленні користуються рухами нижньої щелепи.
Ряд Pleurosauridae. З морської юри й нижньої крейди Західної Європи відомі два або три роди спеціалізованих водних плеврозаврів. Вони мають акродонтні зуби й довгий тулуб і хвіст, що перевищує довжину тіла. У нижньоюрського Palaeopleurosaurus у тілі було тільки 37 хребців, у той же час плеврозавр Pleurosaurus – найбільший не тільки з плеврозаврів, але й ринхоцефалів у цілому (довжина – 1,5 м).

Palaeopleurosaurus posidoniae

Pleurosaurus goldfussi
Надряд Squamata
Лускаті налічують більше 7000 сучасних видів, від крихітних геконів до комодоських варанів і анаконди. Squamata - це спеціалізовані планери, норники, бігуни, скелелази й плавці. Історично (Romer, 1956), лускаті були розділені на дві групи, "Lacertilia" (ящірки, амфісбени) і Ophidia (змії), але філогенетичний аналіз показав, що «Lacertilia» у цьому сенсі не є монофілетичною групою.
Основні апоморфії лускатих, ураховуючи "м'які" органи: редукція хрящових компонентів передньої мозкової коробки, зв'язок слізного протоку з якобсоновим органом, відособлення останнього від головної частини носової капсули, занурення септомаксили, що прикриває у сквамат якобсонів орган, усередину носової капсули, різке укорочення носових кісток, переміщення надскроневої кістки назад від відповідного відростка тім'яної кістки, процельність хребців, наявність тироїдної вирізки в тазовому поясі (є також у кюнеозаврид), злиття астрагала й п'яткової кістки з утворенням мезотарзального суглоба й гачкуватість V метатарзальної кістки. Крім того, ящірки сучасного типу мають велике задньоскроневе вікно, а більшість ящірок - також характерні вирізки по передньому краю коракоїда й нижньої частини лопатки, відсутність кісткових черевних ребер (гастралій).
Ящірки, або форми, близькі до них, відомі з найверхнішої пермі Південної Африки, але представники сучасних родин з'являються тільки в крейді, а більш примітивні ящірки сучасного вигляду, що належать до викопних родин сучасних підрядів - у верхній юрі. Ящірки відрізняються не тільки втратою нижньої скроневої дуги й стрептостилією, але також і наявністю в квадратній кістці вушної вирізки, грацильною слуховою кісточкою й зв'язком парокципітального відростка з квадратною кісткою. Характерним у ящірок сучасного вигляду є те, що хвостові хребці часто поділені надвоє автотомальною септою, що полегшує довільну автотомію (відрив хвоста).
Підряд Eolacertilia. Доюрські «ящірки» відрізняються багатьма примітивними ознаками, до числа яких належить парність усіх кісток даху черепа, субтекодонтність зубів, амфіцельність хребців, пластинчастий тазовий пояс; метатарзальна кістка пальця в них, хоча й сильно укорочена, але не гачкувата. До них належить родина Paliguanidae з родами Palaeagama, Paliguana і Saurosternon з найверхнішої пермі й нижнього тріасу Південної Африки, а також Blomisaurus з нижнього тріасу Кіровської області. Більше відрізняється від палігуанід Boreopricea з нижнього тріасу о-ва Колгуєв, що має сильно подовжений череп, але подібна до Saurosternum парністю грудини [Татарінов, 1978]. Усе це дрібні ящірки з довжиною тіла без хвоста не більше 15-25 см.
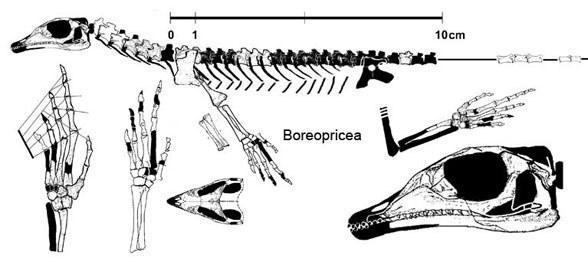
Boreopricea funerea (Tatarinov 1978, Benton and Allen 1997)
До недавнього часу палігуаніди вважалися найдавнішими представниками ящіркового філогенетичного стовбура. Але нещодавно з Мезені, датованого, можливо, верхами казанського або низами татарського ярусу, був описаний за черепом ще один можливий представник дрібних еолацертилій - Lanthanolania ivakhnenkoi (Modesto and Reisz 2003). У цього ящера є верхнє скроневе вікно, облямоване знизу трикутною заочноямковою кісткою, а нижнє скроневе вікно, певно, широко відкрите знизу, зуби дрібні, однорідні.
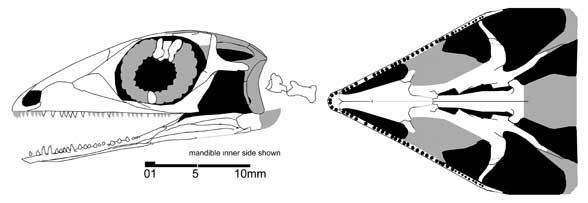
Lanthanolania ivakhnenkoi (Modesto and Reisz 2003)
Татарінов відносить до еолацертилій і кюнеозаврид, однак філогенетичний аналіз розглядає цей таксон як більш примітивний (Lepidosauria+Kuehneosauridae=Lepidosauriformes). Погляди на філогенію лепідозаврів неоднозначні, і пряма спорідненість еолацертилій і ящірок сучасного вигляду потребує додаткового підтвердження.
Ящірки сучасного вигляду. Найдавніші «справжні» ящірки виявлені у верхній частині середньої юри Англії, де знайдені рештки парамацелодид, сцинкоморф (Saurillodon) і ангвіоморф (Parviraptor). До цієї групи може належати й рід Marmoretta, також із середньої юри Англії. Крім того, із середньої юри Мексики описаний амфісбеноподібний діапсид Tamaulisaurus morenoi. Знахідки верхньоюрських ящірок уже численніші, вони походять зі США, Західної Європи, Середньої (Центральної) Азії й Китаю. Найбагатша верхньоюрська фауна ящірок походить із Західної Європи, звідки відома родина ігуан Euposauridae, дві родини гекот (Ardeosauridae і Bavarisauridae), родина кордилоїдів Paramecellodidae. Усі юрські ящірки належать до вимерлих родин, їх зв'язки з більш давніми лепідозавроморфами не вловлюються.
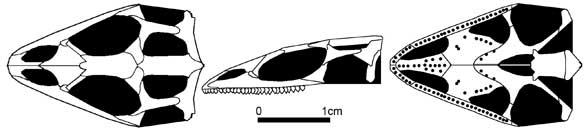
Marmoretta oxoniensis (Evans 1991)
Не менш різноманітна фауна верхньокрейдових ящірок, налічуючи також більше 40 видів, розкопана також у Монголії, у якій ця фауна відрізняється високим ендемізмом і великою кількістю вимерлих родин. У верхній крейді Монголії зареєстровані вимерлі Arretosauridae, Hoplocercidae, можливо, Ardaeosauridae, Slavoiidae, Adamisauridae, Carusiidae, Paramacellodidae, Glyptosauridae, Necrosauridae, можливий дорсетозаврид, параваранід. Із сучасних родин у верхній крейді Монголії відмічені агаміди з групи круглоголовок, іноді виділювані в особливу родину, агаміди вимерлої підродини Priscagaminae, ізодонтозаврові агаміди, різноманітні тейїди, імовірний гімнодактилід, лацертид, гекконід, ксенозаврид і вараніди.
Заслуговує спеціальної уваги одна з найдивовижніших знахідок викопних крейдових ящірок — це Sineoamphisbaena hexataburis з формації Джадохта Внутрішньої Монголії (Китай). Синеоамфісбена - некрупна (довжина черепа близько 4 см) ящірка з коротким і дуже широким черепом, маленькою верхньою скроневою ямою, відособленою знизу заочноямковою кісткою, високою максилою, що контактує з носовою кісткою, і плевродонтними зубами.
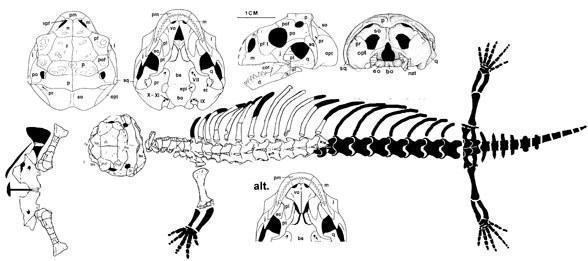
Sineoamphisbaena hextabularis (Wu et al. 1993)
У кайнозої швидко з'являються сучасні роди ящірок, останні вимерлі родини - Arretosauridae і Necrosauridae - зникають до кінця еоцену-початку олігоцену.