
Subclass Lepidosauromorpha
The subclass Lepidosauromorpha today numbers more than 7 thousand species of lizards and snakes and is the second most species-rich group of tetrapods after birds. The term Lepidosauromorpha was introduced by M. Benton in 1983. Lepidosaur representatives, like primitive archosauromorphs, are considered primitive diapsids: currently Archosauromorpha + Lepidosauromorpha make up Sauria, and Sauria + Younginiformes make up Neodiapsida (Laurin, 1991). Lepidosauromorpha probably split off from Archosauromorpha in the middle Permian.
Lepidosaurs descend from lizard-like lepidosauromorph taxa that flourished during the late Permian and early Triassic. During the early Triassic, these diverse forms gave rise to such specialized forms as the kuehneosaurs, the sauropterygians (possibly), and true lepidosaurs. All these reptiles spread out, occupying ecological niches vacated at the end of the Permian extinction, niches previously occupied by the formerly dominant therapsid-like reptiles. Lepidosaurs arose as a small group with an emphasis on small niches not filled by archosaurs.
Characteristic features of lepidosauromorphs are:
- a clumsy posture with sinuous movements (a primitive characteristic inherited from the earliest tetrapods);
- an emphasis on lateral wave-like bending of the spine;
- a rigid rib cage, a large sternum;
- no bipedal specialization;
- the wrists and hands, as well as the cervical vertebrae, are unmodified (by analogy with primitive diapsids);
- overlapping scales.
The progress of lepidosaurs can be explained by the evolution of streptostyly (the anteroposterior mobility of the quadrate bone) and the mobility of the jaws and skull as a whole. Streptostyly in lepidosaurs is achieved through the partial or complete loss of the powerful lower temporal arch. The range of variation in the lower temporal arch of diapsids, and of lepidosaurs in particular, is shown in the figure:
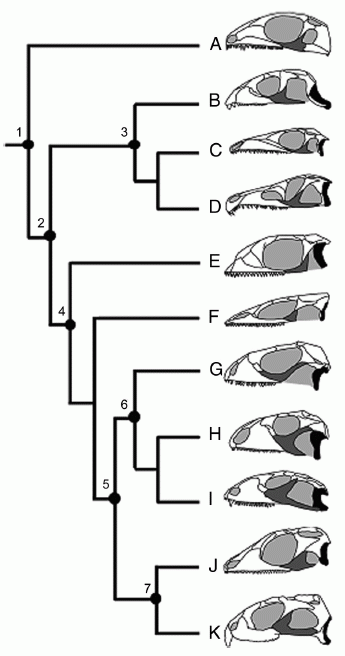
Fig. 1. A simplified phylogeny of diapsid reptiles, showing variation in the development of the lower temporal arch.
The quadrate bone is shown in black, the jugal in gray. (A) the diapsid Petrolacosaurus; (B) the early rhynchosaur Mesosuchus; (C) the archosauromorph Prolacerta; (D) the archosauriform Proterosuchus; (E) the lepidosauriform Kuehneosaurus; (F) the early lepidosaur Marmoretta; (G) the extant squamate Iguana, reflecting a more or less unspecialized form; (H) the huge herbivorous Cretaceous scincomorph Polyglyphanodon; (I) the late Cretaceous lizard Tianyusaurus, unique among squamates for its akinetic rhynchocephalian-like quadrate bone; (J) the rhynchocephalian Gephyrosaurus; and (K) the extant rhynchocephalian Sphenodon. Notes: 1, Diapsida, primitive lower temporal arch, fixed quadrate bone, robust epipterygoid; 2, Sauria, no lower temporal arch, fixed quadrate bone and epipterygoid; 3, Archosauromorpha, no lower temporal arch in basal taxa, but developed in Rhynchosaurs and Archosaurs; 4, Lepidosauriformes and 5, Lepidosauria, no lower temporal arch in basal taxa, quadrate bone and epipterygoid fused with the pterygoid, large squamosal bone; 6, Squamata, overlap of the quadrate and pterygoid is reduced, epipterygoid connected to the pterygoid, lower temporal arch developed only in Tianyusaurus only; and 7, Rhynchocephalia, no lower temporal arch in basal taxa, developed only in some descendants, quadrate, pterygoid, and epipterygoid firmly sutured. From Moazen et al, 2009
Lepidosauromorpha includes the infraclass Lepidosaurs (Lepidosauria) with the superorders Squamates (Squamata) and Beak-heads (Rhynchocephalia or Sphenodontida). Sphenodontians were very widespread and successful from the end of the Triassic to the late Jurassic, but gradually declined during the Cretaceous and Tertiary. At the same time, squamates, which were rare or unknown until the middle Jurassic, remain highly successful today after diversifying in the Cretaceous and Cenozoic. The difference between these two lepidosaur groups lies in their different adaptive strategies: one group evolved a strong, immobile skull adapted for a powerful bite, while the other evolved a skull with reduced strength but high mobility. Snakes acquired the latter adaptation, allowing them to open their mouths extremely wide in order to swallow large prey.
Fossil finds of lepidosauromorphs.
The earliest putative lepidosaurs are the late Permian Lanthanolania (Russia, Modesto and Reisz, 2002) and Saurosternon (South Africa, Carroll, 1975).
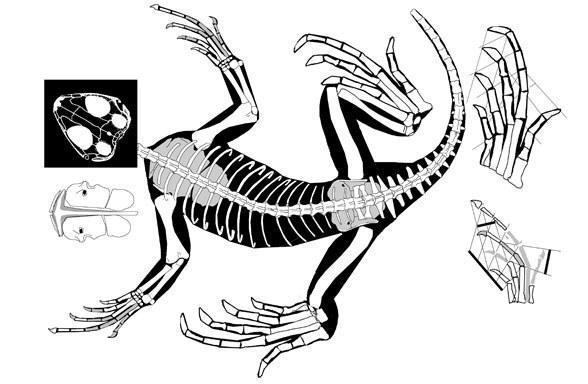
Saurosternon bainii (Huxley 1868)
Paliguana (early Triassic, South Africa, Carroll, 1975) is represented by a single damaged skull with a huge quadrate bone, matching the features of lepidosauromorphs. Paliguana is assigned to Eolacertilia, which also includes the group Marmoretta.
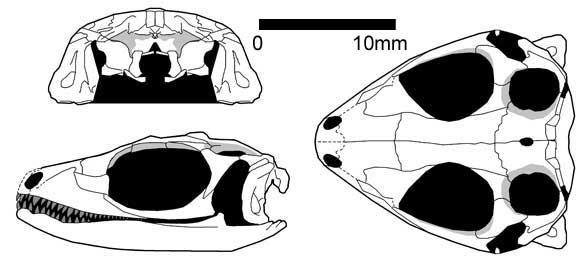
Paliguana whitei (Broom 1903)
In addition, remains attributed to early Triassic lepidosauromorphs have been found in Poland (Czatkowice, Borsuk-Bialynicka
et al., 1999). These deposits formed in an arid environment with localized bodies of water; the fauna included fish, temnospondyls, amphibians, procolophonids, and isolated archosauromorphs.
Kuehneosauria. Kuehneosaurs were specialized, long-ribbed gliders, known from the early Triassic of Poland and the late Triassic of England and North America. A curious feature of the kuehneosaurids was their unusually long gliding membranes, at least twice the length of their body.
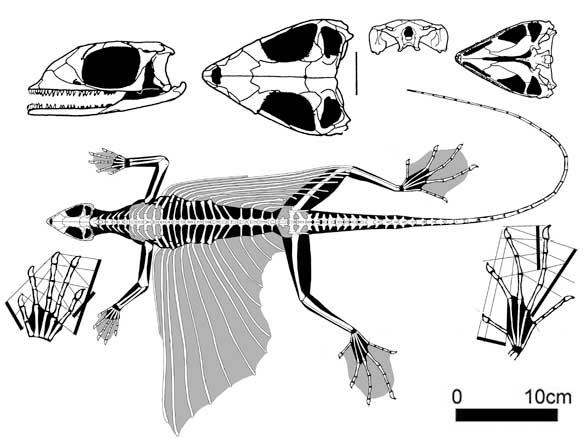
Kuehneosaurus latus (Robinson 1962), late Triassic, England
Kuehneosaurs were able to glide on a skin membrane stretched over 10-11 pairs of movable ribs, articulated with platycoelous vertebrae. The North American Icarosaurus siefkeri had especially long ribs, with a skull only 25 mm long; its "flight" ribs were distinctly curved:

Icarosaurus siefkeri Colbert, 1966
Kuehneosaurs lived on small, humid coastal marine islands together with pterosaurs, archosauriforms, rhynchocephalians, and rare mammals (Robinson, 1962; Fraser, 1994).
Other Mesozoic lepidosauromorphs that do not belong to lepidosaurs include the middle Triassic Coartaredens (England, Spencer and Storrs, 2002) and Megachirella (Italy, Renesto and Posenato, 2003); and the early Jurassic Tamaulipasaurus (Mexico, Clark and Hernandez, 1994). The phylogenetic relationships of Megachirella and Tamaulipasaurus remain unresolved.

Fossil and reconstructed Megachirella wachtleri (Renesto and Posenato 2003).
Tamaulipasaurus morenoi (Clark and Hernandez 1994).
Superorder Rhynchocephalia (Sphenodonta)
The beak-heads are true diapsids of small size. Fossil remains of rhynchocephalians belong to more than 40 taxa, ranging from the late Triassic to the present day. Geographic distribution: Europe, North and South America, China, India, Morocco, South Africa, and New Zealand.
True rhynchocephalians appear in the lower Triassic of South Africa (Palacrodon) and possibly Eastern Europe, but reach their heyday in the upper Triassic of Western Europe, from where more than a dozen genera of sphenodontids have been described. One form has been described from Kenya. From the lower Jurassic of India, two genera of rhynchocephalians have been described from jaw fragments. In the middle Jurassic of Mexico, sphenodontians already appear that are close to the modern tuatara, including the dwarf Zapatodon [Reynoso, Clark, 1998] and Cynosphenodon. Sphenodontids are known from the upper Jurassic of North America (three genera) and Western Europe. From the Cretaceous, only a few sphenodontians are known. No fossil rhynchocephalians are known from the Cenozoic, but the New Zealand rhynchocephalian tuatara - Sphenodon punctatus - survives to this day.

Sphenodon punctatus Evans, 1980
Ecologically, Mesozoic beak-heads were diverse. They included long-bodied marine swimmers (pleurosaurs, saphaeosaurs, Carroll and Wild, 1994), gracile runners (Homoeosaurus, Cocude-Michel, 1963), the armored Pamizinsaurus (Reynoso, 1997), and a group of large-bodied reptiles with hoof-like claw phalanges (Priosphenodon, and Apesteguia Novas, 2003). Among them were insectivorous forms (Gephryosaurus; Diphydontosaurus), opportunistic "predators" (Sphenodon, Dawbin, 1962; Cree et al., 1999), probably venomous predators (Sphenovipera, Reynoso, 2005, known from the middle Jurassic of Mexico), as well as specialized herbivores (for example, Toxolophosaurus, Priosphenodon, Apesteguia and Novas, 2003). Although some of their characteristics are less advanced than those of lizards, some of their primitive features are in fact secondary. Sphenodontians can remain active at lower temperatures than lizards. Beak-heads are also characterized by a long reproductive cycle.

Fig. 2. Sphenodontians
Characteristic structural features of sphenodontians: acrodont teeth with slow replacement, a complete lower temporal arch, an immobile quadrate bone (monimostyly), shortened cervical vertebrae, caudal vertebrae often divided in two by an autotomy septum, which facilitates voluntary autotomy (tail loss). When feeding, they use movements of the lower jaw.
Order Pleurosauridae. From the marine Jurassic and lower Cretaceous of Western Europe, two or three genera of specialized aquatic pleurosaurs are known. They have acrodont teeth and a long body and tail exceeding the length of the body. The lower Jurassic Palaeopleurosaurus had only 37 vertebrae in its body, while the pleurosaur Pleurosaurus - the largest not only of the pleurosaurs, but of rhynchocephalians as a whole (length - 1.5 m).

Palaeopleurosaurus posidoniae

Pleurosaurus goldfussi
Superorder Squamata
Squamates number more than 7000 living species, from tiny geckos to Komodo dragons and anacondas. Squamata are specialized gliders, burrowers, runners, climbers, and swimmers. Historically (Romer, 1956), squamates were split into two groups, "Lacertilia" (lizards, amphisbaenians) and Ophidia (snakes), but phylogenetic analysis has shown that "Lacertilia" in this sense is not a monophyletic group.
The main apomorphies of squamates, including "soft" organs: reduction of the cartilaginous components of the anterior braincase, a connection between the lacrimal duct and Jacobson's organ, separation of the latter from the main part of the nasal capsule, the sinking of the septomaxilla, which in squamates covers Jacobson's organ, into the interior of the nasal capsule, sharp shortening of the nasal bones, the shifting of the supratemporal bone backward from the corresponding process of the parietal bone, procoelous vertebrae, the presence of a thyroid notch in the pelvic girdle (also present in kuehneosaurids), the fusion of the astragalus and calcaneum forming a mesotarsal joint, and hookedness of the fifth metatarsal bone. In addition, lizards of the modern type have a large post-temporal fenestra, and most lizards also have characteristic notches along the anterior edge of the coracoid and the lower part of the scapula, and the absence of bony abdominal ribs (gastralia).
Lizards, or forms close to them, are known from the very uppermost Permian of South Africa, but representatives of modern families appear only in the Cretaceous, while more primitive lizards of modern appearance, belonging to extinct families of modern suborders, appear in the upper Jurassic. Lizards are distinguished not only by the loss of the lower temporal arch and by streptostyly, but also by the presence of an ear notch in the quadrate bone, a slender ear ossicle (columella), and a connection between the paroccipital process and the quadrate bone. A characteristic feature of modern-looking lizards is that the caudal vertebrae are often divided in two by an autotomy septum, which facilitates voluntary autotomy (tail loss).
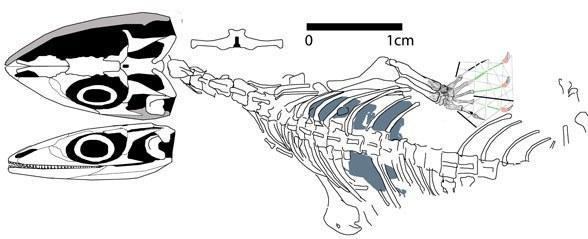
Suborder Eolacertilia. Pre-Jurassic "lizards" are distinguished by many primitive features, including the pairing of all the roofing bones of the skull, subthecodont teeth, amphicoelous vertebrae, a plate-like pelvic girdle; the metatarsal bone of the toe, although greatly shortened, is not hooked. This includes the family Paliguanidae, with the genera Palaeagama, Paliguana, and Saurosternon from the very uppermost Permian and lower Triassic of South Africa, as well as Blomosaurus from the lower Triassic of the Kirov region. Boreopricea, from the lower Triassic of Kolguyev Island, differs more from the paliguanids, having a strongly elongated skull, but is similar to Saurosternon in the pairing of the sternum [Tatarinov, 1978]. All of these are small lizards, with a body length excluding the tail of no more than 15-25 cm.
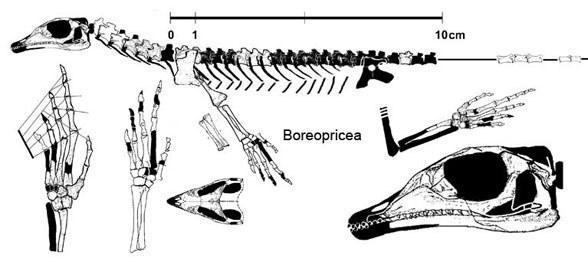
Boreopricea funerea (Tatarinov 1978, Benton and Allen 1997)
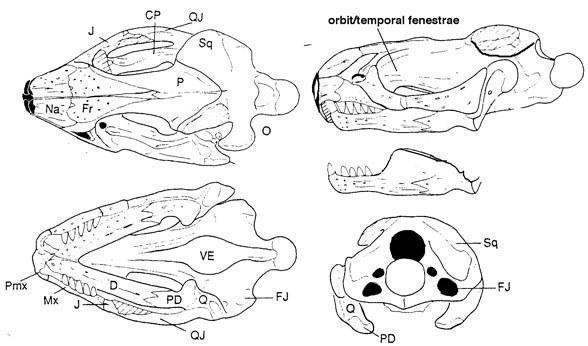
Until recently, the paliguanids were considered the earliest representatives of the lizard phylogenetic stem. But recently, from Mezen, dated possibly to the uppermost Kazanian or lowermost Tatarian stage, another possible small eolacertilian representative was described from a skull - Lanthanolania ivakhnenkoi (Modesto and Reisz 2003). This reptile has an upper temporal fenestra bordered below by a triangular postorbital bone, while the lower temporal fenestra is apparently widely open below; the teeth are small and homogeneous.
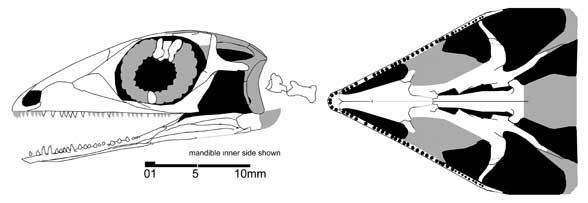
Lanthanolania ivakhnenkoi (Modesto and Reisz 2003)
Tatarinov assigns it to the eolacertilians and kuehneosaurids as well, but phylogenetic analysis treats this taxon as more primitive (Lepidosauria+Kuehneosauridae=Lepidosauriformes). Views on lepidosaur phylogeny are inconsistent, and the direct relationship between eolacertilians and modern-looking lizards requires additional confirmation.
Modern-looking lizards. The earliest "true" lizards are found in the upper part of the middle Jurassic of England, where the remains of paramacellodids, scincomorphs (Saurillodon), and anguimorphs (Parviraptor) have been found. The genus Marmoretta, also from the middle Jurassic of England, may belong to this group as well. In addition, an amphisbaenian-like diapsid, Tamaulisaurus morenoi, has been described from the middle Jurassic of Mexico. Finds of upper Jurassic lizards are already more numerous, coming from the USA, Western Europe, Central Asia, and China. The richest upper Jurassic lizard fauna comes from Western Europe, from which the iguanid family Euposauridae is known, as well as two gecko families (Ardeosauridae and Bavarisauridae), and the cordyloid family Paramacellodidae. All Jurassic lizards belong to extinct families; their connections with more ancient lepidosauromorphs cannot be traced.
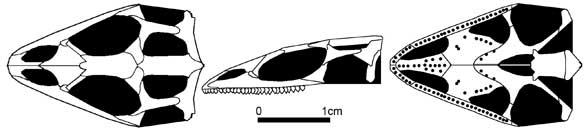
Marmoretta oxoniensis (Evans 1991)
No less diverse is the fauna of upper Cretaceous lizards, likewise numbering more than 40 species, also excavated in Mongolia, where this fauna is distinguished by high endemism and a large number of extinct families. In the upper Cretaceous of Mongolia, the extinct Arretosauridae, Hoplocercidae, possibly Ardaeosauridae, Slavoiidae, Adamisauridae, Carusiidae, Paramacellodidae, Glyptosauridae, Necrosauridae, a possible dorsetosaurid, and a possible paravaranid have been recorded. Among modern families in the upper Cretaceous of Mongolia, agamids of the toad-headed agama group have been recorded, sometimes separated into their own family, agamids of the extinct subfamily Priscagaminae, isodontosaurine agamids, various teiids, a probable gymnodactylid, lacertid, gekkonid, xenosaurid, and varanid.
One of the most remarkable finds of fossil Cretaceous lizards deserves special attention - it is Sineoamphisbaena hexataburis from the Djadokhta Formation of Inner Mongolia (China). Sineoamphisbaena is a small lizard (skull length about 4 cm) with a short and very wide skull, a small upper temporal fenestra separated below by a postorbital bone, a tall maxilla in contact with the nasal bone, and pleurodont teeth.
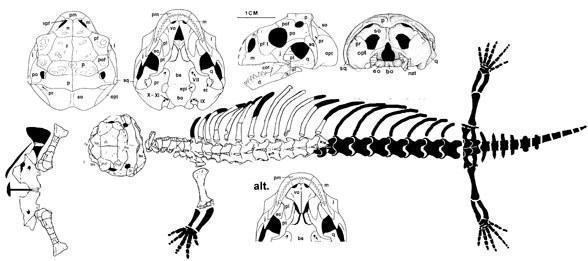
Sineoamphisbaena hextabularis (Wu et al. 1993)
In the Cenozoic, modern genera of lizards appear rapidly, and the last extinct families - Arretosauridae and Necrosauridae - disappear by the end of the Eocene-beginning of the Oligocene.