Шабанов (2002-2006) Каким будет третий синтез в теории эволюции?1
Выкладывая на "Батрахосе" ретроспективу, решил выложить и эту важную для меня статью. Написана она была в 2002 году. Ее не принял "Журнал общей биологии", ее раскритиковал Махаил Александрович Шишкин. В 2006 году ее разместил на свой сайт Александр Марков (тогда на "macro...
Д. А. Шабанов
Яким буде третій синтез у теорії еволюції?
Авторське передповідомлення до мережевої публікації
Завдяки люб'язності творця «Макроеволюції» Олександра Маркова я отримав можливість опублікувати статтю, написану у 2002 році. Повинен визнати, що ця стаття зустріла критичне ставлення і була відправлена на переделку редакцією «Журналу загальної біології», куди я її запропонував. Я дуже вдячний за критику, висловлену Олексієм Меркурьевичем Гіляровим. Коротко резюмуючи її спрямованість, можу сказати, що вона стосується недостатньої «розгорнутості», обґрунтованості ряду тверджень. На жаль, моя спроба переделки цієї статті переконала мене, що для достатнього обґрунтування викладеної в ній ідеї статтю треба перетворювати на книгу. Водночас я сподіваюся, що і в нинішній формі цей текст виявиться джерелом матеріалу для обговорення з однодумцями та конструктивно налаштованими «противниками» — прихильниками інших поглядів.
З відгуків на рукопис цієї статті для мене особливо важливим був відгук Михайла Олександровича Шишкіна. Цей відгук також виявився гострокритичним. З точки зору Михайла Олександровича, пояснювальні підходи синтетичної та епігенетичної теорії принципово різні, і їхнє змішування, продемонстроване в пропонованій статті, не може привести ні до чого доброго. На мій погляд, таке поєднання необхідне. Я вважаю, що однією з нагальних задач побудови нової еволюційної теорії є побудова хорошої класифікації ознак за характером управління їх розвитком. Коли мова йде про біохімічні ознаки бактерій (особливо ознаки «є поломка ферменту»/«немає поломки ферменту») методологія синтетичної теорії еволюції виявляється цілком достатньою. Якщо нас цікавить вироблення нових адаптацій у представників швидко еволюціонуючих високоорганізованих груп організмів, ми не зможемо пояснити їх на мові відбору алелей і повинні будемо звернутися до підходу епігенетичної теорії.
Ще одне необхідне передповідомлення стосується використання терміна «епігенетичний». Це поняття може використовуватися в кількох різних сенсах, навіть якщо не торкатися знаменитої дискусії епігенетиків і преформістів в історії біології розвитку. Ці сенси можна назвати молекулярним і онтогенетичним. Обидва вони в загальних рисах відповідають визначенню К.Х. Воддінгтона, автора цього поняття в його сучасній трактуванні. За Воддінгтоном, епігенетику називають розділ біології про причинні взаємодії між генами і їх продуктами, що формують фенотип. Розрив між молекулярним і онтогенетичним підходами до епігенетики відображає розрив між рівнями дослідження, доступними для сучасної науки. Ми можемо вивчати розвиток або на рівні окремих молекул, або на рівні онтогенезу як цілого. Опис взаємовпливу всіх факторів управління розвитком навіть на рівні однієї клітини – надмірна для сучасної науки задача. Проте сам Воддінгтон використовував обговорюваний термін радше в онтогенетичному сенсі, як, наприклад, у своїй відомій метафорі епігенетичного ландшафту. В онтогенетичному сенсі це поняття використовується і в цій статті (як і в працях М.А. Шишкіна).
Вважаю сенсом цієї мережевої публікації, що стала можливою завдяки А.В. Маркову, обговорення розглянутих у ній ідей. Буду вдячний за конструктивну критику.
Дмитро Шабанов
У першій половині ХХ століття криза класичного дарвінізму (першого еволюційного синтезу) викликала появу другого синтезу — синтетичної теорії еволюції (СТЕ). Сучасна криза СТЕ має призвести до появи третього синтезу, який подолає її редукціоністську обмеженість. Можливі елементи третього синтезу розробляються сьогодні відносно незалежно один від одного. Це екологічна теорія В. А. Красілова, неосальтаціонізм, концепції видового мономорфізму, переривчастого рівноваги, «еволюції еволюції» тощо. Одним із наслідків епігенетичної теорії М. А. Шишкіна, яка може стати ядром третього синтезу, є новий підхід до вирішення проблеми виникнення біологічної цілеспрямованості. Можна припустити, що в рамках третього синтезу буде знайдено пояснення механізму набуття в ході еволюції цілісних комплексів ознак, багаторівневості еволюції та зміни її темпів.
Накануне третього еволюційного синтезу
Згідно з періодизацією розвитку еволюційного вчення, запропонованою Н. Н. Воронцовим (1999), дарвінізм став першим широким синтезом еволюційних ідей. У кінці XIX — на початку XX століття настала криза першого синтезу. Деякі з викликаних цією кризою проблем були вирішені другим синтезом — синтетичною теорією еволюції (СТЕ). Сучасний час — період кризи СТЕ, що передує третьому еволюційному синтезу. Деякі риси майбутнього синтезу почали складатися вже кілька десятиліть тому (наприклад, Шварц, 1967; Красілов, 1984; Шишкін, 1987; Назаров, 1991), проте цей процес ще далеко не завершений.
Сучасний стан еволюційної теорії дозволяє передбачити ряд особливостей третього синтезу. Криза еволюціонізму стимулювала появу ряду нових теорій. Головна думка даної роботи — твердження, що згадані в статті концепції, які розглядають процес еволюції з різних точок зору, добре узгоджуються між собою. Їх об’єднання, на думку автора, має призвести до появи третього еволюційного синтезу. Після характеристики деяких рис кризи СТЕ у цій статті розглядається епігенетична теорія еволюції, яка, на думку автора, має стати ядром нового синтезу.
Деякі авторитетні еволюціоністи висловлювали думку, що накопичення нових даних означає не заміну СТЕ новим синтезом, а лише видозміна та розширення самої СТЕ. Як аналогію можна згадати погляди І.І. Шмальгаузена, який вважав зайвим розмежування дарвінізму і неодарвінізму (СТЕ). На думку автора, у цьому питанні не слід змішувати науку в цілому (у даному випадку — еволюційну біологію) і ту чи іншу теорію (дарвінізм або СТЕ). Наука розвивається, переходячи від одних поглядів до інших. Теорія — продукт певного часу, що характеризується відносно цілісними поглядами. Так, для СТЕ характерне уявлення про еволюцію як про зміну частот алелей у популяціях. Відмова від цього підходу не розширює рамки СТЕ, а вимагає переходу до іншої теорії.
Криза СТЕ
І початковий успіх, і сучасна криза СТЕ пов’язані з її редукціонізмом. Прояви його різні: це і зведення видоутворення і макроеволюції до мікроеволюції, і відрив еволюції популяцій від змін біогеоценозів і біосфери, і прийняття припущення про незалежну еволюцію окремих генів. Зокрема, математична теорія відбору базується на припущенні, що всі алелі мають постійну пристосувальну цінність. При цьому не враховуються такі важливі фактори, як вплив генотипу, невизначеність результатів онтогенезу (обумовлена взаємодією зовнішніх і внутрішніх факторів), а також екологічна неоднорідність середовища.
Називаючи проблеми, які були вирішені завдяки синтезу теорії еволюції і генетики, прихильники СТЕ часто посилаються на критику дарвінізму Ф. Дженкіна (Jenkin, 1867). За цим автором, адаптивна ознака не може зберегтися в ряді поколінь через «розмивання» при схрещуванні її носіїв з особинами, позбавленими даної ознаки. «Після переоткриття законів Менделя і доказу, що фактори, що визначають розвиток успадкованих ознак, передаються нащадкам не розпадаючись, «кошмар Дженкіна» був спростований» (Сойфер, 1975, с. 309). Однак очевидно, що Дженкін мав на увазі зовсім не моногенні ознаки: «Припустимо, білий чоловік потрапив у кораблекрушення на острові, населеному неграми... Наш постраждалий герой, ймовірно, стане королем; він вб’є велику кількість чорних у боротьбі за існування; він матиме величезну кількість жінок і дітей, тоді як багато його підданих будуть жити і помруть холостяками. ... У першому поколінні буде кілька дюжин розумних молодих мулатів, у середньому перевершуючих інтелектом негрів. Ми можемо очікувати, що трон протягом кількох поколінь займатиме більш-менш жовтий король; але чи зможе хтось повірити, що весь острів поступово набуде білої або навіть жовтої популяції, або що островитяни набудуть енергії, хоробрості, винахідливості, настирливості, самоконтролю, витривалості, завдяки яким наш герой вбив так багато їх предків і породив так багато дітей, тобто ті якості, які фактично відбирає боротьба за існування, якщо вона може щось відбирати?» (Jenkin, 1867, p. 289–290).
Для оцінки роздуму Дженкіна не важливо, що расового переваги не існує, а названі ознаки успадковуються в основному культурно. Якщо адаптивність особини залежить від комплексу незалежно успадкованих ознак, «розсипання» такого комплексу має перешкоджати відбору. Припущення, що кожна з таких ознак буде відбиратись незалежно від інших, спростовано в рамках самої СТЕ. Розробляючи математичну теорію відбору, Дж. Холдейн (Haldane, 1957) показав, що відбір за кількома незалежними парами алелей неефективний через наростання генетичного навантаження і зниження репродуктивного потенціалу популяції. Припущення про тривалий відбір за багатьма окремими генами, кожен з яких вносить свій внесок на рівні фенотипу (особливо характерне для розвиненої на основі СТЕ соціобіології) не лише суперечить розрахункам Холдейна, а й спростовується даними про відносну невелику кількість генів людини та інших видів, отриманими в ході робіт з розшифрування геномів.
Створенню СТЕ сприяло відкриття С. С. Четвериковим генетичного різноманіття носіїв нормальних фенотипів із природних популяцій. Однак з точки зору СТЕ цей феномен так і не знайшов належного пояснення (Левонтин, 1978). Не менш складно встановити причини стійких у часі різниць між сусідніми популяціями і групами розмноження, виявлених у багатьох видах. Для пояснення цих явищ прихильники СТЕ вдаються до посилань на дрейф генів. Фактично це означає відмову від селекціонізму, що лежить в основі самої СТЕ.
Вивчення філогенезу багатьох таксонів показало помилковість характерного для СТЕ уявлення про переважно дивергентний характер еволюції, тісно пов’язаного з визнанням ненаправленості мутацій і дрейфу генів. Паралелізми і конвергенції є нормою у становленні більшості груп (Татаринов, 1987).
З сказаного випливає, що теорія, яка замінить СТЕ, повинна буде пояснити механізм набуття в ході еволюції узгоджених комплексів адаптивних ознак.
Деякі інші риси кризи другого синтезу розглянуті далі у порівнянні з альтернативними поглядами.
Епігенетична теорія М. А. Шишкіна
Центральну роль у третьому еволюційному синтезі (Гродницький, 2001) може зіграти концепція М. А. Шишкіна (1987, 1988), названа ним «епігенетичною теорією еволюції Шмальгаузена‑Воддінгтона». Хоча автор і виходить з ідей І. І. Шмальгаузена і К. Х. Воддінгтона, він приходить до суттєво інших висновків. Тому, ймовірно, правильніше говорити про «епігенетичну теорію М. А. Шишкіна».
Ключове поняття даної теорії — епігенетична система (далі — ЕС), сукупність взаємодій генетичних і інших факторів, що впливають на онтогенез. Відповідно до цієї теорії окремі успадковані задатки не є безпосередньою причиною тих чи інших особливостей фенотипу, а лише впливають на систему управління онтогенезом. У результаті природного відбору відбираються і відтворюються у потомстві не окремі алелі або ознаки, а цілісні фенотипи. Незалежно від того, які фактори (генетичні та негентетичні успадковані задатки, зовнішні впливи) викликають розвиток адаптивної ознаки, її власники внесуть відносно більший внесок у формування наступного покоління.
Потомки успадковують від батьків не лише гени, а й загальну організацію ЕС. Якщо характер відбору залишається прежнім, перевагу отримують ті, які відтворюють у своєму розвитку відобрані в попередньому поколінні адаптивні фенотипи. Відбір таких фенотипів має призвести до перебудови ЕС, що підвищує стійкість траєкторій розвитку, що їх викликає (креодів).
Поки популяційна норма залишається адаптивною, відбір перебудовує ЕС таким чином, щоб при максимально широкому різноманітті генотипів забезпечувався нормальний хід онтогенезу. Наслідком цього саме є виявлене С. С. Четвериковим генетичне різноманіття носіїв нормального фенотипу з природних популяцій. Відбір проти склалої фенотипічної норми дестабілізує провідні до неї епігенетичні креоди. У результаті проявляється ряд відносно дискретних аберацій, які відповідають можливим для даної ЕС альтернативним креодам. Відбір на користь одного з них збільшує стійкість його реалізації в онтогенезі.
Таким чином, згідно з розглянутими поглядами, нові ознаки з’являються внаслідок перетворення видоспецифічної ЕС шляхом відбору. При цьому стабілізуються варіанти розвитку, спочатку виникаючі як аберації.
Сказане підтверджується численними фактами. Як відомо, однозначного відповідності ген‑ознака не існує. Дія кожного гена залежить від інших успадкованих задатків і характеризується тією чи іншою експресивністю і пенетрантністю. Особливо нестабільно проявлення мутантних алелей, реалізація яких в онтогенезі не стабілізована відбором: навіть домінантні мутації не завжди не проявляються в гомозиготі. Р. Гольдшмідт (Goldschmidt, 1940) показав, що одна і та сама ознака може розвиватися або не розвиватися в силу дуже різних причин (різних впливів на ЕС): і мутацій різних генів, і впливів середовища.
Розвиток епігенетичної теорії розпочався з експериментів Воддінгтона, у яких була продемонстрована стабілізація лабільних ознак. У цих експериментах відбір на здатність формувати морфози (аномалії розвитку) dumpy або bithorax у відповідь на температурні або токсичні впливи приводив до того, що ці морфози почали розвиватися і за нормальних умов (Waddington, 1957). З точки зору концепції М. А. Шишкіна, такий результат обумовлений перебудовою ЕС, що стабілізує відбирані шляхи розвитку (а не виникненням генів dumpy або bithorax).
У експериментах Г. Х. Шапошникова (1978), в результаті відбору морфоз тлії, викликаних зміною кормової рослини, виникали нові, морфологічно та екологічно своєрідні форми експериментальних тварин, репродуктивно ізольовані від вихідних.
Епігенетична система як рівень інтеграції
Виникає закономірне питання: чи слід розглядати ЕС як окремий рівень регуляції розвитку, якщо взаємодія генів розглядається і в класичній генетиці? Чи не є поняття «епігенетична система» синонімом поняття «генотип»?
Очевидно, що виділення окремого структурного або функціонального рівня інтеграції виправдане в тому випадку, коли на ньому виникають нові (емергентні) властивості, відсутні на нижчележачих рівнях. Недооцінка одного з суттєвих рівнів інтеграції (рівня n) призведе до типічної редукціоністської помилки: особливості рівня n+1 будуть інтерпретовані як результат функціонування рівня n‑1. При цьому емегентні властивості рівня n стануть недоступними для вивчення.
Онтогенез, один із найскладніших відомих процесів, регулюється різноманітними успадкованими задатками, середовищними впливами і їх взаємодією. Еквіфінальність індивідуального розвитку і залежна від зовнішніх умов можливість його адаптивної зміни задаються на рівні не окремих генів, а ЕС в цілому. Розгляд цього рівня є необхідною умовою вивчення найважливіших властивостей живих систем.
Де локалізовані механізми каналізації розвитку? Це питання не більш коректне, ніж, наприклад, питання про те, в якому місці мозку знаходиться дана думка, чи де в біосфері розташований регулятор вмісту кисню в атмосфері. Коли мова йде про складні системи (ЕС, мозок, біосфера тощо), вказати локалізацію їх емегентних властивостей просто неможливо, оскільки вони задаються сукупністю зв’язків між усіма елементами. Проте епігенетичні механізми проявляються і на рівні окремих успадкованих задатків, мислення пов’язане з функціонуванням клітин мозку, а біосферний гомеостаз здійснюється завдяки життєдіяльності окремих організмів.
Закономірно, що ядром першого еволюційного синтезу стало дослідження змінності фенотипів і їх взаємодії зі середовищем — найявнішого рівня регуляції історичного розвитку (рис. 1). Вивчення відносно простого механізму успадкованої змінності і впливу відбору на частоти успадкованих задатків привело до другого синтезу. Припускається, що в ході третього синтезу буде зрозуміле значення найскладнішого рівня регуляції розвитку — ЕС.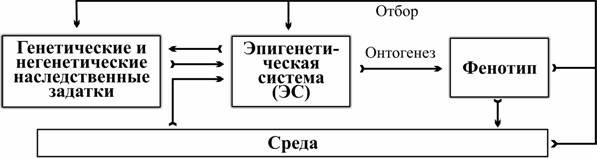
Рис. 1. Роль ЕС у управлінні індивідуальним і історичним розвитком
Подолання «геноцентризму»
Трактуючи описані вище експерименти щодо закріплення морфоз у результаті відбору, К. Х. Воддінгтон назвав виявлене ним явище генетичною (а не епігенетичною) асиміляцією. Для пояснення результатів аналогічних експериментів висловлювалося припущення, що початкові феноадаптації замінюються геноадаптаціями (Шмальгаузен, 1938; Гаузе, 1984). Воно засноване на переконанні, що кожна ознака обумовлена специфічними генами. Такий «геноцентризм» широко поширений і до сьогодні. Наприклад, твердження, що «матеріалом для еволюції ... слугують мутації і їх комбінації», Н. Н. Воронцов (1999, с. 470) відносить до числа положень, «про які не сперечаються еволюціоністи».
Показано, що близькі види можуть суттєво відрізнятися організацією свого генетичного матеріалу, наприклад, співвідношенням різних типів послідовностей (Корочкін, 1985). Водночас близькість цих видів пов’язана зі схожістю їх ЕС. Ймовірно, саме схожість ЕС різних видів є причиною існування гомологічних рядів змінності, описаних Н. І. Вавиловим.Оскільки гомологічні аберації розвитку можуть мати різне генетичне підґрунтя, гомологічними є не гени, а епігенетично детерміновані креоди.
Як зазначає А. С. Раутіан (1993), у силу принципу відносності змісту інформації А. А. Ляпунова, інформація в генотипі доступна лише певному компетентному користувачеві. Таким користувачем є не фенотип у цілому, а саме ЕС, первісно успадкована від попереднього покоління (наприклад, при утворенні яйцеклітини), а потім змінювана в онтогенезі. Зміна фенотипів у ході еволюції пов’язана не стільки зі зміною генів (і інших успадкованих задатків), скільки зі зміною ЕС. Одна й та сама послідовність нуклеотидів, прочитана ЕС різних організмів, може мати для них зовсім різний сенс! Розглянута у популярному кінофільмі (і романі) можливість отримати динозавра, пересадивши його гени в яйцеклітину жаби, є необґрунтованою фантазією: таким шляхом можна отримати лише ембріон жаби з порушеною взаємодією генотипу і ЕС. Як відомо, у ході антропогенезу послідовності ДНК людини зазнали лише незначних змін. Таким чином, людина відрізняється від шимпанзе не стільки тим, що має інші гени, скільки наявністю реорганізованої ЕС.
«Геноцентризму» протирічить також зростаюча кількість прикладів негенетичного успадкування. Ймовірно, динамічне успадкування (передача специфічної конформації білкових молекул), що проявляється при прионних захворюваннях, є не аномалією, а результатом дії загального механізму (Інге-Вечтомов, 2000). Так, шаперони-білки беруть участь у синтезі багатьох поліпептидних ланцюгів, визначаючи їх конформацію. Способністю до успадкування володіє навіть цитоскелет, що передає особливості просторової організації клітинних структур (Албертс та ін., 1987, т. 3, с. 132). Культурне успадкування у людини (і, у меншій мірі, у деяких інших видів) забезпечує передачу набутих ознак і перехід до ефективніших способів вироблення адаптацій.
Як показано на рис. 1, ЕС не лише залежить від успадкованих задатків, а й впливає на них. Прикладами таких впливів на генетичну успадкованість можуть слугувати епігени (Голубовський, Чураєв, 1987), а на негенетичну — динамічно успадковані зміни конформації білків. Експериментально встановлено здатність бактерій переструктурувати свій генетичний апарат протягом життя однієї клітини, забезпечуючи зміни генів, необхідні для живлення у певному середовищі (Голубовський, 2001). При вирощуванні льону на збагаченій біогенами ґрунті відбувалася генерація успадкованих змін, що призводили до збільшення розмірів рослин (Marx, 1984). Наведені факти можуть здаватися відповідаючими поглядам Т. Д. Лисенка. Однак принциповою особливістю цих результатів є те, що вони отримані в коректних експериментах і позбавлені ідеологічної підкладки.
Як відомо, характерною особливістю зміни наукової парадигми є переосмислення відомих фактів. Епігенетична теорія дає підстави для ревізії таких понять, як ген, мутація, модифікація, рухомий відбір тощо (Шишкін, 1987). Неправомірним виявляється ототожнення менделєвських генів з локалізованими в хромосомах цитострами. Насправді менделєвський ген є перемикачем між двома епігенетично стабілізованими траєкторіями індивідуального розвитку, а не причиною їх виникнення. На противагу загальноприйнятим поглядам, можна виділити лише два основних типи відбору: стабілізуючий і дестабілізуючий. Ознаки слід ділити не на успадковані (генотипічні) і неуспадковані (фенотипічні), а на стійкі та лабільні у своєму прояві.
**Проблема цілеспрямованості вирішена?**
Проблема біологічної цілеспрямованості стала ключовою проблемою для науки про природу ще за часів Аристотеля. Уявлення, що цілеспрямовані організми виникли випадково, з’явилося в античності і в різних формах було виражено Емпедоклом, Епікуром і Лукрецієм. Дарвінізм додав до цього уявлення гіпотезу про довгі послідовності малих ненаправлених змін, випадково підвищуючих пристосованість організмів. Сучасна нелінійна термодинаміка вивчає самоорганізацію як наслідок випадкових збурень. Проте уявлення про виникнення цілеспрямованості на основі випадкових відхилень не можна вважати остаточно доведеним. Йому протирічить той факт, що мутації зазвичай несприятливі і не інтегровані в генотип. Для вчених, переконаних у досконалості природи, ідея випадкового виникнення цілеспрямованості неприйнятна. Ймовірно, саме тому Л. С. Берг постулював іманентну, недоступну науковому аналізу цілеспрямованість живого (Берг, 1977).
Епігенетична теорія відкриває можливість нового вирішення проблеми цілеспрямованості, що виходить за межі запропонованої Бергом класифікації (там же, с. 99–101). Цілеспрямованість мутацій, постулювана в СТЕ, — явище на межі дива. Навпаки, викликані дестабілізацією нормального розвитку аберації потенційно цілеспрямовані, оскільки в їх формуванні бере участь ЕС, сформована попередньою еволюцією даного виду і життя в цілому. Мутація гена не пов’язана з відповідними змінами інших елементів генотипу; епігенетична аберація може зачіпати багато ознак при збереженні кореляцій між ними. Мутації виникають випадково, а аберації — коли колишня популяційна норма перестає бути адаптивною. Нарешті, як зазначає Д. Л. Гродницький (2001), закономірність появи аберантних фенотипів зумовлює достатню ймовірність зустрічі їх носіїв.
Якщо якась з виникаючих аберацій виявляється адаптивною, у популяції відбувається процес, змодельований описаними експериментами Воддінгтона: відбір на стійкість її відтворення в ряді поколінь. Кінцева стадія — стабілізація нової норми.
Отже, нова ознака проходить такі етапи свого становлення: більш‑менш адаптивна «модифікація», що розвивається за відповідного впливу середовища; один з альтернативних варіантів розвитку, що все більш стійко реалізується в онтогенезі; норма, для розвитку якої не потрібні специфічні зовнішні впливи.
Щоб пояснити появу (часто паралельну у різних груп організмів) комплексів цілеспрямовано взаємопов’язаних ознак, слід розглядати вплив регуляторного механізму, що лежить вище рівня окремих успадкованих задатків — ЕС. Саме таков шлях подолання «кошмара Дженкіна».
П. Тейяр де Шарден (1987) стверджував, що життя рухається вперед «на дотик». Ці слова зазвичай розуміли як констатацію неминучості накопичення випадкових помилок. Ймовірно, аналогія Тейяра значно глибша. Хоча в еволюційному процесі немає заздалегідь визначеного кінцевого стану, у ньому немає і випадкового метання з боку в бік. Напрямок кожного кроку еволюції обумовлений реакцією цілісного організму на зміну середовища, у якому він розвивається.
**Екологічна теорія еволюції**
Характерний для СТЕ редукціонізм проявляється не лише у уявленні системи управління розвитком у вигляді мозаїки генів, а й у трактуванні популяції як основного театру еволюційних подій.
Кожен біоценоз володіє певною здатністю підтримувати гомеостаз. Зміни зовнішніх (наприклад, кліматичних) факторів можуть компенсуватися внутрішніми регуляторними механізмами даного біоценозу. Саме вони підтримують характерну структуру екологічних ніщ у певному діапазоні значень зовнішніх умов. При цьому на популяції діє стабілізуючий відбір нормального розвитку, внаслідок чого вони не змінюються або еволюціонують узгоджено і повільно. Еволюцію в межах стабільного біоценозу В. А. Красилов (1969) назвав когерентною еволюцією. У випадку розпаду біоценозу спостерігається швидка некогерентна еволюція окремих його елементів. Популяції найпластичніших і найменш інтегрованих у зруйноване співтовариство видів здатні існувати і в змінених умовах. Припинення стабілізуючого відбору та різноманітність умов розвитку призводять до дестабілізації онтогенезу і прояву резерву змінності, характерного для ЕС даних видів. У нестабільних спільнотах перевагу отримують r‑стратегії. Результатом r‑відбору стає прискорення індивідуального розвитку, що може призводити до часткової втрати спеціалізації, досягнутої на останніх етапах онтогенезу. Мозаїчність ділянок території, що перебувають на різних стадіях сукцесії та формування нових біоценозів, сприяє квантовій еволюції.
Так, у антропоцентрично змінених біоценозах у ряду видів зафіксовано швидке виникнення адаптивних ознак (наприклад, нових форм гніздової поведінки птахів або спрощення життєвого циклу проходних риб).
В епігенетичній теорії не розглядаються причини того, чому відбір починає підтримувати одну з аберацій, а не попередню норму. Екологічна теорія пояснює стрибкоподібну зміну характеру відбору. Обидві концепції акцентують увагу на цілісних властивостях досліджуваних систем і успішно доповнюють одна одну.
**Концепція видового мономорфізму**
Цікаво порівняти висновки епігенетичної теорії еволюції з даними про існування мономорфного комплексу генетичних і фізіологічних ознак у видів. «Одне з найважливіших властивостей еукаріотичного геному — двоїстість його структурно‑функціональної організації, що знаходить пряме відображення у співіснуванні двох реальних явищ: поліморфізму і мономорфізму» (Алтухов, 1989, с. 219). Внутрівидовий мономорфізм характерний для генетичних систем з великою кількістю структурних компонентів і високою надмірністю (там же, с. 203), тобто для регуляторної частини геному.
Аналогічно, Х. Карсон розрізняє дві системи генетичної змінливості: «відкриту», представлена локусами, що забезпечують внутрішньовидовий поліморфізм, і «закриту», пов’язану з коадаптованими блоками генів (Carson, 1982). Видоутворення, з цієї точки зору, є наслідком реорганізації «закритої» частини геному.
Д. К. Беляєв (1974) встановив, що дестабілізуючий відбір за одними ознаками призводить до прояву широкої змінливості інших, раніше стабільних, ознак. Можливо, це явище пов’язане з реорганізацією «закритої» частини геному та ЕС.
Вивчення змінливості геному приводить до цілком узгодженого з епігенетичною теорією висновку: «на надвидовому рівні еволюція — це не лише, або точніше, не стільки процес появи нових генів з новими функціями, скільки швидка реорганізація генетичного матеріалу з подальшим розвитком нових систем взаємодії (і регуляції) генів на посттранскрипційному та посттрансляційному рівнях» (Алтухов, 1989, с. 216).
С. С. Шварц показав, що «адаптації спеціалізованих видів і адаптації окремих популяцій широко розповсюджених видів йдуть принципово різними шляхами» (1980, с. 45). Адаптації внутрішньовидових форм до змінних умов середовища виражаються у морфофункціональних зрушеннях (зміні розмірів серця чи нирок, здатності крові поглинати кисень тощо). Видові адаптації відбуваються на тканинному і біохімічному рівнях, що робить морфофункціональні зрушення зайвими. Цілісність виду, поряд з генетичними механізмами, підтримується видоспецифічною реакцією на зміни умов середовища (там же, с. 132), яку можна розглядати і як відображення спільності ЕС.
Деякі ознаки, демонструючи різницю між різними таксонами, виявляються мономорфними всередині більшості з них (і цим нагадують видові ознаки, пов’язані з «закритою» частиною геному та ЕС). Так, у всіх ссавців однаковий розмір еритроцитів, інший, ніж у птахів, рептилій, риб і безхвостих амфібій. Єдина група хребетних, у якій зафіксовано суттєве різноманіття розмірів еритроцитів — хвостові амфібії (Шмідт‑Нільсен, 1987, с. 128).
Ймовірно, у рамках третього синтезу доведеться відмовитися від характерного для СТЕ градаціонізму (виходячи з уявлення про те, що пристосованість виникає внаслідок випадково корисних мутацій). Можна припустити, що у ньому будуть розмежовані періоди стабілізації і дестабілізації системи управління розвитком. Обґрунтованим є припущення, що мономорфна частина видового геному є основою видоспецифічної ЕС. Засвоєння нової адаптивної зони, виникнення таксонів того чи іншого рівня, швидше за все, буде відбуватися внаслідок перестройки основних характеристик ЕС.
У цьому контексті представляє інтерес епігенетична концепція виду, запропонована К. Є. Михайловим у роботі, опублікованій в Internet (Михайлов, url). За цією концепцією, основна характеристика виду — стабілізований онтогенез. При такому підході популяції і особини, що перебувають на переходовому етапі видоутворення в період дестабілізації онтогенезу, слід розглядати як девіантні форми, близькі до одного з видів, але не належні до нього. Розвиваючи погляди Михайлова, можна включити наявність стабілізованих епігенетичних креодів і в характеристику надвидових таксонів. Неочікуваним наслідком цього підходу може стати обґрунтування виділення перехідних груп, що не належать до таксонів деяких вищих рангів, тобто форм incertae sedis (наприклад, сімей, що не належать ні до одного з отрядів свого класу).
**Неосальтаціонізм і переривчаста рівновага**
Одним із можливих шляхів реорганізації геному є хромосомні перестройки (Алтухов, 1989; Воронцов, 1999). Ймовірна тісна зв’язок між реорганізацією геному і гибридизацією (Боркін, Даревський, 1980).
У останні десятиліття отримані численні підтвердження значення стрибкоподібних подій для еволюції, що змусило згадати про сальтаціонізм, здавалося б, спростованим другим еволюційним синтезом. Вивчення кариотипів близьких видів показало важливу роль хромосомних перестроек і поліплоїдії у видоутворенні не лише у рослин, а й у тварин (в тому числі у досить прогресивних групах, наприклад у жаб і гризунів). Це дозволяє припускати, що у багатьох випадках виникнення часткової чи повної ізоляції передує екологічній або морфофізіологічній диференціації (Воронцов, 1999). У такому випадку конкурентне виключення кариологічно ізольованих форм має сприяти їх дивергенції.
Можна навести приклади достатньо вивчених сальтаційних подій. Штам **Clostridium botulinum**, що викликає ботулізм, при зараженні певним фагом перетворюється у **Clostridium novyi**, збудника газової гангрени (Eklund et al., 1974). При виникненні кількох родів ссавців відбувалося закріплення аберацій hairless, мопсовидності, папіломатозу (Воронцов, 1999, с. 507–507). Один з ферментів (Cu‑Zn‑супероксидисмутаза) біолюмінесцентної бактерії **Photobacter leiognathi** схожий не з ферментами прокаріот, а з ферментами срібнобрюхових риб **Leiognathidae**, симбіонтами яких є ці бактерії. Це свідчить про «горизонтальний перенос» успадкованої інформації (там же, с. 518). Зміна гормонального балансу додаванням тироксину викликає у ілистого стрибака (**Periophthalmus**) перестройку онтогенезу, включаючи розвиток «кінцівок» замість грудних плавців (Harms, 1934). За даними молекулярної систематики, основні групи організмів виникли внаслідок послідовних ендосимбіозів (Кусакин, Дроздов, 1994) — стрибкоподібних еволюційних подій. Нарешті, деякі еволюційні події (наприклад, походження вторинноротових тварин від первинноротових, з’єднання хоанами нюхового каналу з ротовою порожниною у риб) не могли відбуватися градуально.
Наведені та інші факти демонструють недопустимість характерного для СТЕ зведення макроеволюції і видоутворення до мікроеволюції. Неосальтаціонізм, як і епігенетична, і екологічна теорії, пояснює нерівномірність темпів еволюції, на якій базується концепція переривчастої рівноваги. Можливість освоєння нової адаптивної зони може з’явитися внаслідок тих чи інших сальтаційних подій. До них належать суттєва зміна ходу розвитку в результаті макромутації, набуття адаптивної ознаки при горизонтальному переносі або цілісного комплексу ознак при гибридизації (Боркін, Даревський, 1980), інтеграції або симбіогенезу. Подальша еволюція системи управління розвитком буде відбуватися під впливом відбору, що стабілізує деякі з нових варіантів розвитку.
**Зростання ефективності еволюційного процесу**
З точки зору СТЕ швидкість еволюції має зменшуватися зі складністю генетичної системи організмів і уповільненням їх поколінь. Це суперечить факту, що темпи еволюції прискорюються зі зростанням морфофізіологічної складності організмів. Дивовижним моментом в історії Землі є так званий «кембрійський вибух»: виникнення за відносно короткий час переважаючої більшості коли‑небудь існуючих типів тварин, багато з яких незабаром вимерли. Навряд чи причини цього події кореняться в малих ненаправлених популяційно‑генетичних змінах. Швидше, воно викликане появою механізмів, що суттєво підвищують ефективність макроеволюції. Наприклад, це могло бути набуття організмами білатеральної симетрії тіла, пов’язане не стільки з набуттям відповідних генів, скільки з розвитком епігенетично регульованих варіантів розвитку клітин, тканин і органів, що відповідають за диференціацію частин тіла. Стабілізації відбором найуспішніших планів будови тіла мало передувати широке різноманіття їх варіантів. Сьогодні за план будови відповідає добре регульований епігенетичний комплекс, і значних новацій у сучасних організмах не спостерігається.
Як зазначав І. І. Шмальгаузен, наслідком міжгрупового відбору є витіснення форм, що відстають у темпах і якості пристосування до змінного середовища. Такий процес має призводити до удосконалення самого механізму еволюції (Шмальгаузен, 1968; Завадський, Колчинський, 1977).{
"translated_text": "Прискорення еволюції у високоорганізованих формах (Расницин, 1978) — результат дії такого міжгрупового відбору.\nЗа всією видимістю, у міру ускладнення організмів все більшу роль у формуванні їхніх ознак відіграє епігенетичний компонент. Біохімічні ознаки прокаріот визначаються в основному на генному рівні (це широко використовується в генній інженерії). Що стосується морфологічних ознак високо розвинених організмів, то вони залежать від складної системи епігенетичних взаємодій. Існування механізмів, що забезпечують цілісність виду і проявляються у видовому мономорфізмі, також може бути наслідком «еволюції еволюції».\nПідвищенню ефективності еволюціонування багатьох видів сприяє підроздільність їхньої популяційної структури (Коновалов, 1974; Алтухов, 1989). Так, навіть на відносно однорідній території зелені (Bufo viridis) і сірі (B. bufo) жаби формують локальні групи, які за багатьма ознаками відрізняються сильніше, ніж підвиди або близькі види (Шабанов, 2001, 2002). Значна кількість рівнів різноманіття, на більшості з яких може діяти груповий відбір (Шабанов, Шабанова, 2001), підвищує його ефективність.\nТаким чином, є підстави припускати, що магістральним шляхом еволюції є удосконалення епігенетичних механізмів регуляції індивідуального і історичного розвитку. Отже, ефективність перебудови ЕС під дією відбору може бути мірою еволюційного прогресу тих чи інших груп. Найважливіші віхи цього шляху — розвиток регуляції експресії бактеріальних цистронів; виникнення статевого розмноження; утворення багатотканинних організмів; появлення механізму культурної спадковості.\nДеякі підсумки\nНа підставі викладеного можна зробити ряд висновків.\n1. Наш час — період кризи другого еволюційного синтезу (СТЕ) і формування третього синтезу. Такий синтез призначений описати шляхи виникнення адаптивних комплексів ознак, дію спрямовуючих еволюцію і регулюючих її темпи факторів, історичне удосконалення механізмів еволюції.\n2. Відмінною особливістю третього синтезу має стати системний підхід до еволюції, її вивчення на різних рівнях з визначенням інтегруючих механізмів для кожного з них.\n3. Третій синтез має об’єднати досягнення різних відносно незалежних концепцій (класичного дарвінізму, біології індивідуального розвитку, сучасної генетики, епігенетичної теорії М. А. Шишкіна, екологічної теорії В. А. Красілова, неосальтаціонізму, концепцій видового мономорфізму, «еволюції еволюції» та перервистого рівноваги і інших).\n4. Уявлення про стрибкоподібний характер видоутворення узгоджується з даними про реорганізацію геному при виникненні виду і видовому мономорфізмі. Екологічна і епігенетична теорії, а також неосальтаціонізм пояснюють причини нерівномірності темпів еволюції, а концепція перервистого рівноваги зосереджується на його результатах.\n5. Епігенетична теорія М. А. Шишкіна акцентує увагу на ключовому рівні регуляції онтогенезу. Одним із її наслідків є новий підхід до вирішення проблеми виникнення біологічної доцільності.\nЯкі дані можуть слугувати підтвердженням викладених тут поглядів? Піддається перевірці припущення, що при розвитку в контрольованих умовах онтогенез особин, що походять з порушених (в тому числі антропогенних) екосистем, мав би бути менш стабільним, ніж у тих, що походять з еталонних місцевих середовищ.\nДля підтвердження епігенетичної теорії представиться корисною перевірка припущення, що алелі менделєвського гена є не послідовностями нуклеотидів, а заданими всією епігенетичною системою перемикачами між стабілізованими варіантами норми. Для цього бажано провести їх експериментальне отримання і руйнування. Припустимо, при відборі в дестабілізованій популяції зберігаються носії як однієї, так і іншої з двох альтернативних аберацій. Чи з’являться в результаті такого відбору менделювальні епігенетичні перемикачі онтогенезу? Чи буде цей процес ефективніший за умов культивації, суттєво відмінних від нормальних (наприклад, при вирощуванні drosophila на нестандартному середовищі)? Чи призведе відбір на стійке відтворення гетерозиготного фенотипу (що виникає при схрещуванні ліній, у яких стабілізовані різні варіанти норми) до припинення його менделєвського розщеплення? Один із варіантів такого відбору — збереження власників гетерозиготного фенотипу з тих виводків, де їхня частка була максимальною.\nПеревірити деякі положення даної статті дозволить вивчення характеру змінливості мономорфної частини геному у експериментальних популяціях, підданих гострому відбору на зміну адаптивної норми.\nГоловна мета, яку ставив перед собою автор — стимулювати обговорення і експериментальне вивчення порушених проблем.\nСписок цитованої літератури\nАлбертс Б., Брей Д., Льюїс Дж., Рефф М., Робертс К., Ватсон Дж. Молекулярна біологія клітини: У 5‑ти т. Т. 3. М.: Мир, 1987. 296 с.\nАлтухов Ю. П. Генетичні процеси в популяціях. М.: Наука, 1989. 328 с.\nБеляєв Д. К. Про деякі питання стабілізуючого і дестабілізуючого відбору // Історія і теорія еволюційного вчення. Л.: 1974. С. 76–84.\nБерґ Л. С. Труди з теорії еволюції. Л.: Наука, 1977. 387 с.\nБоркін Л. Я., Даревський І. С. Сітчасте (гібридогенне) видовотворення у хребетних // Журн. общ. біол. 1980. Т. 41. № 4. С. 485–506.\nВоронцов Н. Н. Розвиток еволюційних ідей у біології. М.: Видав. відд. УНЦ ДО МГУ, Прогрес‑Традиція, АБФ, 1999. 600 с.\nГаузе Г. Ф. Екологія і деякі проблеми походження видів // Екологія і еволюційна теорія. Л.: Наука, 1984. С. 5–105.\nГолубовський М. Д. Неканонічні спадкові зміни // Природа. 2001. № 8. С. 3–9.\nГолубовський М. Д., Чураєв Р. Н. Динамічна спадковість і епігени // Природа. 1997. № 4. С.16–25.\nГрант В. Еволюційний процес. Критичний огляд еволюційної теорії. М.: Мир, 1991. 488 с.\nГродницький Д. Л. Епігенетична теорія еволюції як можлива основа нового еволюційного синтезу // Журн. общ. біол. 2001. Т. 62. № 2. С. 99–109.\nЗавадський К. М., Колчинський Е. І. Еволюція еволюції (історико‑критичні нариси проблеми). Л., «Наука», 1977. 236 с.\nІнге‑Вечтомов С. Г. Приони дріжджів і центральна догма молекулярної біології // Вісн. РАН. 2000, Т. 70. №4. С.299–306.\nКоновалов С. М. Субізолят як відносно жорстка система. Структура субізолята // Журн. общ. біол. 1974. Т. 35. № 6. С. 819–838.\nКорочкін Л. І. Паралелізми в молекулярній організації геному і проблеми еволюції // Молекулярні механізми генетичних процесів. М.: Наука, 1985. С. 132–146.\nКрасілов В. А. Філогенія і систематика. Проблеми філогенії і систематики. Владивосток, 1969. С. 12–30.\nКрасілов В. А. Теорія еволюції: необхідність нового синтезу // Еволюційні дослідження. Макроеволюція. Владивосток: ДВНЦ АН СРСР, 1984. С. 4–17.\nКусакин О. Г., Дроздов А. Л. Філема органічного світу. Ч. 1. Прологеми до побудови філеми. СПб.: Наука, 1994.\nЛевонтин Р. Генетичні основи еволюції. М.: Мир, 1978. 352 с.\nМихайлов К. Є. Типологічне осмислення «біологічного виду» і шляхи стабілізації околосвітової таксономії у птахів // https://rav.sio.rssi.ru/~anna/spbird.html\nНазаров В. І. Вчення про макроеволюцію. На шляхах до нового синтезу. М.: Наука, 1991. 288 с.\nРасницин А. П. Темпи еволюції і еволюційна теорія (гіпотеза адаптивного компромісу) // Еволюція і біоценотичні кризи. М.: Наука, 1987. С. 46–64.\nРаутіан А. С. Про природу генотипу і спадковості // Журн. общ. біол. 1993. Т. 54. № 2. С. 131–148.\nСойфер В. Н. Генетика. Зародження хромосомної теорії спадковості // Історія біології з початку ХХ століття до наших днів. М.: Наука, 1975. С. 295–313.\nТатаринов Л. П. Паралелізм і спрямованість еволюції // Еволюція і біоценотичні кризи. М.: Наука, 1987. С. 124–144.\nТейяр де Шарден П. Феномен людини. М.: Наука, 1987. 240 с.\nШабанов Д. А. Мозаїчне різноманіття зелених жаб (Bufo viridis) у межах однорідного фізико‑географічного регіону // Питання герпетології. М.: МГУ, 2001. С. 334–336.\nШабанов Д. А. Популяційне різноманіття видів роду Bufo в Харківській області як предмет вивчення і охорони // Вісн. Харківськ. ун‑ту. 2002. № 551. Ч. 2. С. 232–236.\nШабанов Д. А., Шабанова А. В. Рівні різноманіття зелених жаб (Bufo viridis) // Фальцфейнівські читання. Херсон: Терра, 2001. С. 212–213.\nШапошников Г. Х. Динаміка клонів, популяцій і видів і еволюція // Журн. общ. біол. 1978. Т. 39. № 1. С. 15–33.\nШварц С. С. Сучасні проблеми еволюційної теорії // Вопр. філос. 1967. № 10. С. 143–153.\nШварц С. С. Екологічні закономірності еволюції. М.: Наука, 1980. 280 с.\nШишкін М. А. Індивідуальний розвиток і еволюційна теорія // Еволюція і біоценотичні кризи. М.: Наука, 1987. С. 76–124.\nШишкін М. А. Еволюція як епігенетичний процес // Сучасна палеонтологія. Методи, напрямки, проблеми, практичне застосування. М.: Недра, 1988. Т. 2. С. 142–169.\nШмальгаузен І. І. Шляхи і закономірності еволюційного процесу. М., 1938. 231 с.\nШмальгаузен І. І. Кібернетичні проблеми біології. Новосибірськ, 1968. 224 с.\nШмідт‑Нільсен К. Розміри тварин: чому вони такі важливі? М.: Мир, 1987. 259 с.\nCarson H. L. Speciation as a major reorganization of polygenic balances// Mechanisms of speciation. NewYork: Liss., 1982. P. 411–433.\nEklund M. W., Poysky F. T., Meyers J. A., Perloy G. A. Interspecies conversion of Clostridium botulinum type C to Clostridium novyi type A by bacteriophage // Science. 1974. Vol. 186. 456–458. (цит. по Грант, 1991, с. 189)\nGoldschmidt R. The material basis of evolution. New Haven, London, Oxford, 1940. 436 p.\nHaldane J. B. S. The cost of natural selection // J. Genet. 1957. Vol. 55. P. 511–524.\nHarms, J. W. Wandlungen des Artgefüges. Heine, Tübingen. 1934. 212 p. (цит. по Goldschmidt, 1940)\nJenkin F. The origin of species Art. I.// North Brit. Rev. 1867, June. Vol. 46. P. 277–318.\nMarx J. L. Instability in plants and the ghost of Lamarck // Science. 1984. Vol. 224. P. 1415–1416.\nWaddington C. H. The Strategy of the Genes: a Discussion on Some Aspects of Theoretical Biology. London: Allen and Unwin, 1957. 262 p."
}