
Шабанов (2015) Эволюционная экология популяционных систем... (Автореферат)
Русский перевод автореферата диссертации, защищенной 26 марта 2015 года, а также ссылка на украинский автореферат в pdf.
Нижче подано російський переклад автореферату. Український текст автореферату у форматі pdf лежить тут.
МІНІСТЕРСТВО ОБРАЗУВАННЯ І НАУКИ УКРАЇНИ
ДНІПРОПЕТРОВСЬКИЙ НАЦІОНАЛЬНИЙ УНІВЕРСИТЕТ ІМЕНІ ОЛЕСЯ ГОНЧАРА
ШАБАНОВ ДМИТРО АНДРІЙОВИЧ
УДК (597.841::574.4):[477.5+292.485]
ЕВОЛЮЦІЙНА ЕКОЛОГІЯ ПОПУЛЯЦІЙНИХ СИСТЕМ
ГІБРИДОГЕННОГО КОМПЛЕКСУ ЗЕЛЕНИХ ЖАБ
(PELOPHYLAX ESCULENTUS COMPLEX)
ЛІВОБЕРЕЖНОЇ ЛЕСОСТЕПІ УКРАЇНИ
03.00.16 — екологія
АВТОРЕФЕРАТ
дисертації на здобуття вченого ступеня доктора біологічних наук
Дніпропетровськ — 2015
Дисертація є рукописом
Робота виконана на кафедрі зоології та екології тварин біологічного факультету Харківського національного університету імені В. Н. Каразина Міністерства освіти і науки України
Офіційні опоненти доктор біологічних наук, професор
Серебряков Валентин Валентинович,
Київський національний університет
імені Тараса Шевченка, кафедра зоології, завідувач;
доктор біологічних наук, професор
Домнич Валерій Іванович,
Запорізький національний університет,
кафедра охотоведення та їхтіології, завідувач;
доктор біологічних наук, доцент
Жуков Олександр Вікторович
Дніпропетровський національний університет
імені Олеся Гончара, кафедра зоології та екології, професор.
Захист відбудеться «_26_» березня 2015 року о _10.00_ на засіданні спеціалізованої вченої ради Д 08.051.04 для захисту дисертацій на здобуття наукового ступеня доктора біологічних наук у Дніпропетровському національному університеті імені Олеся Гончара Міністерства освіти і науки України за адресою: 49010, м. Дніпропетровськ, проспект Гагаріна, 72, корпус 17, факультет біології, екології і медицини, ауд. 611.
З дисертацією можна ознайомитися в науковій бібліотеці Дніпропетровського національного університету імені Олеся Гончара за адресою: м. Дніпропетровськ, вул. Казакова, 8.
Автореферат розіслано «_26_» _лютого_ 2015 р. Учений секретар спеціалізованої вченої ради,
Учений секретар спеціалізованої вченої ради,
кандидат біологічних наук, доцент А. А. Дубіна
ЗАГАЛЬНА ХАРАКТЕРИСТИКА РОБОТИ
Актуальність теми. П’ятдесят років тому, у 1964 році польський гідробіолог Лешек Бергер виявив, що класичний об’єкт експериментальної біології, відомі ще Карлу Ліннею їстівні жаби, Pelophylax esculentus (Linnaeus, 1758), не є звичайним видом (Berger, 1964). Це міжвидовий гибрид жаби ставкової, Pelophylax lessonae (Camerano, 1882) і жаби озерної, Pelophylax ridibundus (Pallas, 1771). Зусиллями групи європейських дослідників було показано, що розмноження Pelophylax esculentus підтримується завдяки геміклональній спадковості (клональній передачі з покоління в покоління окремих геномів батьківських видів у відносно незмінному вигляді). Наразі встановлено, що геміклональна спадковість вимагала зміни базових механізмів, що забезпечують розмноження подавляючої більшості еукаріотичних організмів. Незвичність зелених жаб проявляється на всіх рівнях їх біології: від функціонування їх популяційних систем і способу еволюції до механізмів роботи їх клітин. Вивчення такого незвичного феномена дозволяє розширити наше розуміння закономірностей, що стосуються сфери популяційної та еволюційної екології, а також цілого ряду інших біологічних наук.
Десять років тому за участі дисертанта в Харківській області, у лісостеповій частині басейну річки Північний Донетц були описані незвичні за складом популяційні системи зелених жаб, Pelophylax esculentus complex (Mass…, 2004; Массовая..., 2005; Изучение…, 2006). За час їх вивчення стало ясно, що вони належать до числа найскладніших з відомих науці. Вивчення популяційних систем гибридогенного комплексу зелених жаб у лісостеповій частині басейну Північного Донця вимагало створення на кафедрі зоології та екології тварин Харківського національного університету імені В. Н. Каразина робочої групи, науковим керівником якої є дисертант. Це дослідження покликане інтегрувати результати, отримані робочою групою за участі дисертанта, і створити теоретичні рамки, що організують подальше вивчення відкритого в ході спільної роботи природного феномена.
До початку виконання дисертаційного дослідження склад популяцій і популяційних систем представників Pelophylax esculentus complex Лівобережної лісостепі України був вивчений надзвичайно нерівномірно. Склад цих систем у частині Лівобережної Лісостепі, розташованої у водозбірному басейні Дніпра (Лівобережно-Дніпровській фізико‑географічній провінції), був встановлений у ряді робіт, виконаних з використанням електрофорезу білкових маркерів та іншими методами співробітниками Інституту зоології імені І. І. Шмальгаузена НАНУ (Морозов‑Леонов, 1998; Некрасова, Морозов‑Леонов, 2001; Некрасова, 2002). Тому в нашій роботі достатньо було вивчити склад таких систем лише для східної частини цієї провінції. Розташовану в басейні Північного Донця (приєднання Дону) Середньоруську фізико‑географічну провінцію, що належить до Лісостепі України, слід було дослідити максимально повно, у зв’язку з її недостатньою вивченістю. Крім того, популяційні системи Pelophylax esculentus complex, аналогічні тим, що реєструються в Лісостепі України, зустрічаються і в тій частині Степу, де поширені елементи лісостепних ландшафтів. Тому частину досліджень треба було виконати в Донечко‑Донській північностепній фізико‑географічній провінції.
Зв’язок роботи з науковими програмами, планами, темами. Робота виконана у відповідності до науково‑дослідних тем кафедри зоології та екології тварин біологічного факультету Харківського національного університету імені В. Н. Каразина: «Неортодоксальне видоутворення (гібридизація, немендлєвська спадковість, поліплоїдія та сітчаста еволюція): вивчення на прикладі амфібій» (№ державної реєстрації 0109U007439; 2009‑2010 рр.); «Розробка технологій отримання клонального потомства хребетних тварин шляхом схрещування напівклональних міжвидових гибридів (на прикладі зелених жаб)» (№ державної реєстрації 0112U001026; 2012‑2014 рр.).
Мета і завдання дослідження. Мета роботи — з’ясувати закономірності функціонування та еволюції популяційних систем Pelophylax esculentus complex Лівобережної лісостепі України.
Для досягнення зазначеної мети у роботі поставлені такі завдання:
— встановити склад популяційних систем Pelophylax esculentus complex, характерних для Середньоруської фізико‑географічної провінції, для східної частини Лівобережно‑Дніпровської фізико‑географічної провінції, а також для прилеглої до Лісостепі частини Донечко‑Донської північностепної фізико‑географічної провінції;
— виділити в регіоні дослідження окремі субрегіони, що відрізняються складом поширених у них популяційних систем Pelophylax esculentus complex; виявити біоекзоценотичні та географічні фактори, що визначають розподіл по їх території популяційних систем Pelophylax esculentus complex;
— визначити ті субрегіони дослідження, які представляють найбільший інтерес для вивчення і мають найвищу цінність з точки зору збереження їх біорізноманіття; встановити характерні особливості поширених у цих субрегіонах представників Pelophylax esculentus complex;
— з’ясувати специфіку біосистем, у яких відбувається геміклональне розмноження Pelophylax esculentus;
— розробити способи вивчення стійкості та закономірностей трансформацій популяційних систем Pelophylax esculentus complex з використанням імітаційного моделювання;
— побудувати динамічну типологію популяційних систем, що включають диплоїдних представників Pelophylax esculentus complex;
— визначити рівні відбору, що впливають на динаміку біосистем, у яких відбувається геміклональне розмноження Pelophylax esculentus; встановити можливі механізми підтримки відбором геміклонального розмноження гибридів у Pelophylax esculentus complex.
Об’єкт дослідження — популяційні системи Pelophylax esculentus complex Лівобережної лісостепі України.
Предмет дослідження — закономірності функціонування та підтримання стійкості популяційних систем Pelophylax esculentus complex, їх різноманіття та еволюційне значення.
Методи дослідження:
— якісне визначення типу популяційних систем зелених жаб за складом мешкаючих у них форм;
— формалізований опис місць проживання популяцій і популяційних систем Pelophylax esculentus complex,
— оцінка чисельності та складу популяційних систем з використанням маркування та повторного відлову;
— первинне визначення видової приналежності особин за ознаками зовнішньої морфології;
— експрес‑визначення плоїдності особин з використанням цитометрії;
— скелетохронологічне визначення віку особин і ретроспективна оцінка швидкості їх росту на підставі аналізу розмірів ліній склеювання на зрізах трубчастих кісток;
— порівняльні екологічні, морфологічні, біохімічні та фізіологічні дослідження представників Pelophylax esculentus complex;
— генотипування особин з використанням проточної ДНК‑цитометрії, електрофорезу білкових маркерів;
— кариоаналіз роздавлених препаратів мазкових тканин, розкритих метафазних пластинок і розкритих клітин;
— визначення геномного складу гамет з використанням проточної ДНК‑цитометрії, електрофорезу білкових маркерів, аналізу маркерів хромосом типу лампових щіток та аналізу результатів схрещувань;
— імітаційне моделювання та експерименти з імітаційними моделями; статистична обробка результатів моделювання (аналіз методом головних компонент);
— порівняння результатів моделювання з емпіричними даними, формалізація емпіричних і літературних даних.
Наукова новизна отриманих результатів. Результати, що відображають наукову новизну, полягають у наступному: вперше:
— у Лівобережній лісостепі України виявлені регіони, де масово поширені поліплоїдні Pelophylax esculentus;
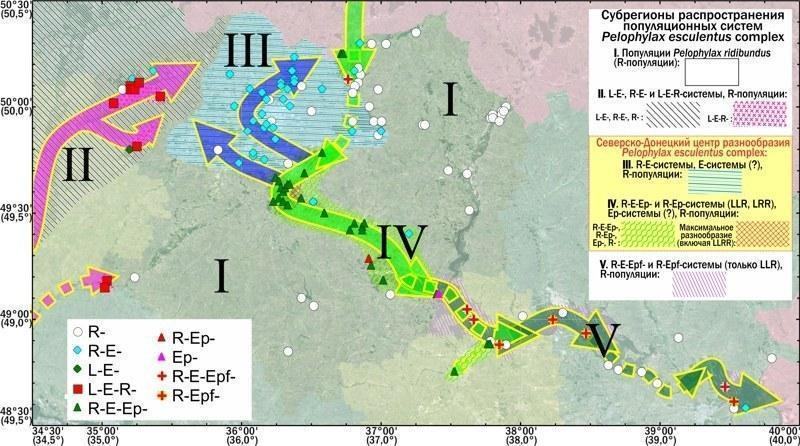
— у регіоні дослідження виділено п’ять субрегіонів, які відрізняються набором типів популяцій і популяційних систем представників Pelophylax esculentus complex, поширених на їх території;
— встановлені біоекзоценотичні зв’язки, що визначають склад локальних популяційних систем Pelophylax esculentus complex у кожному з описаних субрегіонів;
— виявлено і описано Північнодонецький центр різноманіття Pelophylax esculentus complex;
— запропоновано поняття «геміклональна популяційна система» (ГПС) і обґрунтовано, що ГПС є особливою категорією біосистем, характерною для гібридогенних комплексів видів;
— обґрунтовано, що трансформації ГПС є особливим типом динаміки біосистем; встановлені ключові фактори, що визначають ці трансформації;
— розроблено концептуальну модель трансформацій ГПС, на основі якої створені імітаційні моделі цих процесів;
— розроблено динамічну типологію ГПС Pelophylax esculentus complex, що включають диплоїдні міжвидові гибриди;
— за допомогою імітаційного моделювання ГПС Північнодонецького центру різноманіття Pelophylax esculentus complex, що складаються з диплоїдних представників, сформульовано доступні для перевірки припущення про склад передаваних у них клональних геномів;
— висунуто гіпотезу, що пояснює феномен гибридної амфіспермії конкуренцією в клітинах зароджувального шляху клональних геномів, змінених внаслідок відбору на їх здатність викликати елімінацію альтернативних геномів;
— різниці у стійкості різних типів ГПС Pelophylax esculentus complex розглянуто як прояв групового відбору;
— висунуто гіпотезу, що розглядає здатність до геміклональної гибридизації як адаптацію до гомологічної міжвидової рекомбінації, підтримуваної відбором видів;
отримали подальший розвиток:
— уявлення про різноманіття популяційних систем Pelophylax esculentus complex;
— класифікація типів популяційного розмноження і уявлення про зв’язок цих типів з особливостями біосистем індивідуального, популяційного та видового рівнів;
— способи використання імітаційних моделей для дослідження емпірично зареєстрованого різноманіття природних біосистем;
— концептуальна база для опису і моделювання популяційних систем гібридогенних комплексів видів з геміклональною спадковістю;
— уявлення про форми типології біосистем і способи побудови динамічних типологій складних систем;
— концепція стійкості біосистем, як їх здатності зберігати свій динамічний тип з плином часу;
— концепція багаторівневого відбору;
— уявлення про відбір клітинних ліній у гондах Pelophylax esculentus;
— теоретичні уявлення про закономірності еволюції рекомбінації;
— концепція відбору видів.
Практичне значення отриманих результатів. Отримані в ході роботи результати важливі для планування та організації охорони унікальних популяційних систем Pelophylax esculentus complex. Розроблені способи використання імітаційних моделей дозволяють оцінювати стійкість складних біологічних систем і корисні для тестування гіпотез, що компенсують нестачу емпіричних даних. Продемонстровано можливість використання імітаційних моделей для розробки динамічної типології біосистем. Отримані в ході роботи дані та узагальнення використовуються при викладанні загальних і спеціальних курсів «Екологія», «Моделювання в екології», «Батрахологія і герпетологія», «Біологічна статистика в зоології та екології», «Зоологія хребетних», а також при проведенні навчально‑польової практики з зоології хребетних на біологічному факультеті ХНУ імені В. Н. Каразина.
Особистий внесок здобувача. Дисертаційна робота є самостійним і оригінальним дослідженням.
Ряд публікацій, що відображають розглянуті в дисертації теми, підготовлені у складі робочої групи популяційної екології амфібій ХНУ імені В. Н. Каразина. Автор є науковим керівником цієї групи; він грав керівну роль при постановці завдання для дослідження, брав особисту участь у всіх етапах роботи, у тому числі польових, і керував інтерпретацією результатів. Деякі з результатів, представлених у даній роботі, були частково відображені в кандидатських дисертаціях А. В. Коршунова (2010) і М. А. Кравченка (2013), виконаних під керівництвом автора. Ряд публікацій підготовлений у результаті міжнародного співробітництва з науковими колективами Росії, Німеччини та інших країн. У цих публікаціях автор брав участь у постановці завдання, особисто збирав матеріал, брав участь у його обробці та інтерпретації результатів.
Апробація результатів дисертації. Матеріали дисертації опубліковані на III, IV і V Міжнародних наукових конференціях Zoocenosis-2005, Zoocenosis-2007 і Zoocenosis-2009 (Дніпропетровськ); науковій конференції «Сучасні проблеми зоології та екології» (Одеса, 2005); IX і XI Міжнародних науково‑практичних екологічних конференціях, присвячених популяційній екології (Бєлгород, 2006 і 2010); III і V з’їздах Герпетологічного товариства ім. А. М. Нікольського (Пущино, 2006; Мінськ, 2012); науковій конференції біологічного факультету ХНУ, присвяченій 100‑річчю зі дня народження Є. І. Семененка (Харків, 2006); Міжнародній конференції «Сучасні проблеми біологічної еволюції» (Москва, 2007); III, IV, V, VI і VII з’їздах Українського герпетологічного товариства (Мелітополь, 2007; Дніпропетровськ, 2008; Харків, 2010; Київ, 2011; Вілково, 2013); III Міжнародної конференції молодих вчених «Біологія: від молекули до біосфери» (Харків, 2008); Міжнародної науково‑практичної конференції «Біорізноманіття і стале розвиток» (Сімферополь, 2010); I Всеукраїнської науково‑практичної конференції «Сучасна проблематика і методологія біоекологічних досліджень: популяційний підхід» (Івано‑Франківськ, 2012); конференції‑школи «Аномалії і патології амфібій і рептилій: методологія, еволюційне значення, можливість оцінки здоров’я середовища» (Єкатеринбург, 2013); конференції, присвяченої 100‑річчю Біологічної станції ХНУ імені В. Н. Каразина (Гайдари, 2014).
Публікації. За матеріалами дисертації опубліковано 51 наукову публікацію, у тому числі 31 стаття в профільних наукових журналах і збірниках наукових трудів, з них — 9 у виданнях, включених у міжнародні бібліографічні бази, 7 статей в інших виданнях, а також 14 наукових публікацій у матеріалах і тезах конференцій.
Структура і обсяг дисертації. У склад дисертації входить перелік умовних позначень, введення, 9 глав, висновок, підсумки і список використаної літератури. Робота містить 36 таблиць і 62 рисунка. Матеріали дисертації викладені на 364 сторінках комп’ютерного набору, з яких основного тексту — 275 сторінок. Список використаної літератури містить 419 джерел, з яких 125 — латиницею.
ПЕРЕЛІК УМОВНИХ ПОЗНАЧЕНЬ ТА ВИЗНАЧЕННЯ СПЕЦИФІЧНИХ ТЕРМІНІВ
Терміни подано в логічному, а не в алфавітному порядку.
Pelophylax esculentus complex
Група європейських зелених жаб: ставкова жаба, Pelophylax lessonae (Camerano, 1882), озерна жаба Pelophylax ridibundus (Pallas, 1771), а також різні форми їх міжвидових гибридів: їстівна жаба, Pelophylax esculentus (Linnaeus, 1758), для якої використовується назва, аналогічна видовій
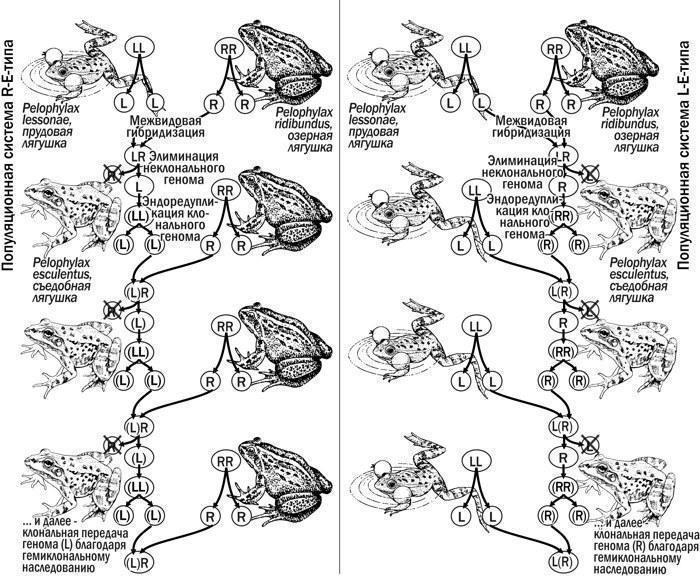
Геміклональна спадковість
Спадковість у міжвидових гибридів, при якій один з батьківських геномів (клональний геном) передається (у типічному випадку) у гамети цілком, без рекомбінації
Клональні геноми: (L), (R)
Геноми, що передаються потомству як ціле, без рекомбінації: (L) — геном P. lessonae, (R) — геном P. ridibundus
Рекомбінантні геноми: L, R
Геноми, що передаються з рекомбінацією (як у батьківських видів): L — геном P. lessonae, R — геном P. ridibundus
XL, XR, (XL), (XR)
Жіночі геноми (що несуть X‑хромосому)
YL, YR, (YL), (YR)
Чоловічі геноми (що несуть Y‑хромосому)
Кариогенетичні форми
Форми представників P. esculentus complex, що відрізняються складом геномів у їх генотипі та у продукуваних ними гаметах
R(L), L(R), (L)(R)
Диплоїдні P. esculentus з різними клональними геномами
LLR, LRR{
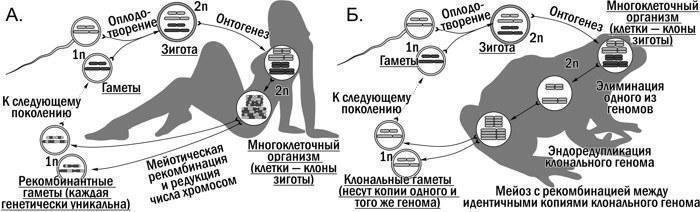
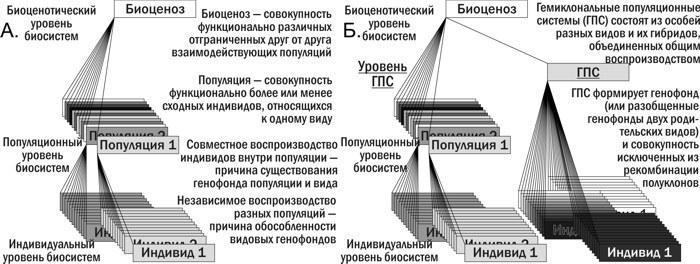
"text": "Триплоїдні P. esculentus\n\n\nГПС,\n\t\t\tгеміклональна\n\t\t\tпопуляційна\n\t\t\tсистема\n\n\nСукупність спільно мешкаючих та розмножуючихся представників різних форм гібридів та особин батьківських видів, у якій з покоління в покоління передаються як клональні, так і рекомбінантні геноми\n\n\nТрансформації ГПС\n\n\nФорма динаміки ГПС, що полягає у зміні в них чисельності та співвідношення різних каріогенетичних форм\n\n\nR-E-L-тип ГПС; R-Ep-тип ГПС\n\t\t\t(тощо)\n\n\nПозначення типів ГПС в аналітичній класифікації, де літера R позначає P. ridibundus, E — P. esculentus, а L — P. lessonae, а літерою p позначається наявність у ГПС поліплоїдних P. esculentus\n\n\nКонспецифічний геном\n\n\nКлональний геном того ж виду, що й переважаючий батьківський вид у ГПС (наприклад, (R) у ГПС R-E-типу); поняття застосовне для ГПС R-E- та L-E-типу\n\n\nГетероспецифічний геном\n\n\nКлональний геном іншого виду, ніж переважаючий батьківський вид у ГПС (наприклад, (L) у ГПС RE-типу); поняття застосовне для ГПС RE- та LE-типу\n\n\nРозмноження\n\n\nФеномен організменного рівня організації біосистем, пов’язаний з появою у організму нащадків; слугує основою популяційного відтворення\n\n\nВідтворення\n\n\nФеномен популяційного рівня організації біосистем, пов’язаний з підтриманням певного складу внаслідок розмноження організмів\n\n\nГібридоліз\n\n\nПоява особин батьківського виду в результаті схрещування гібридів, що клонально передають геноми одного виду\n\n\nГібридна амфіспермія\n\n\nПродукування міжвидовими гібридами (P. esculentus) суміші гамет, частина з яких несе геном P. lessonae, а частина — P. ridibundus. Диплоїдні гібриди з гібридною амфіспермією в даній роботі позначаються (L)(R).\n\n\nКлон\n\n\nСукупність клітин або особин з ідентичним (з точністю до помилок копіювання) генотипом. Організм людини в типовому випадку є клоном клітин; однояйцеві близнюки є організмами-клонами\n\n\nГеміклон\n\n\nСукупність особин, що мають ідентичний (з точністю до помилок копіювання) клональний геном та різні рекомбінантні геноми\n\nОСНОВНИЙ ЗМІСТ РОБОТИ\nСУЧАСНИЙ СТАН ПРОБЛЕМИ\nНаведено стислий опис історії відкриття та вивчення гібридогенезу в Pelophylax esculentus complex, а також обговорення його екологічного значення. Показано, що типовий для еукаріот життєвий цикл (рис. 1.А.) є причиною характерної для них ієрархії біосистем (рис. 2.А.) Геміклональне успадкування, характерне для P. esculentus та інших міжвидових гібридів, є глибокою видозміною цього циклу (рис. 1.Б.). Питання про те, як така видозміна відображається на різних рівнях біосистем, зокрема тих, що розглядаються в рамках екології, залишається недостатньо вивченим.\n





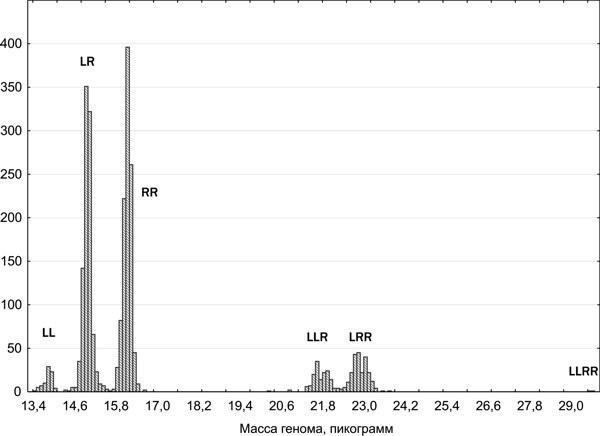
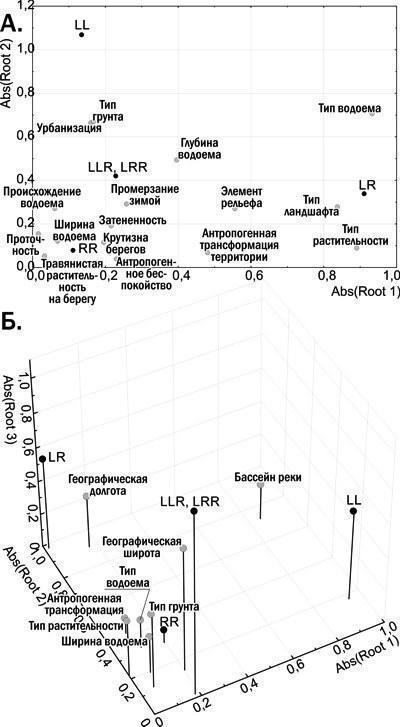
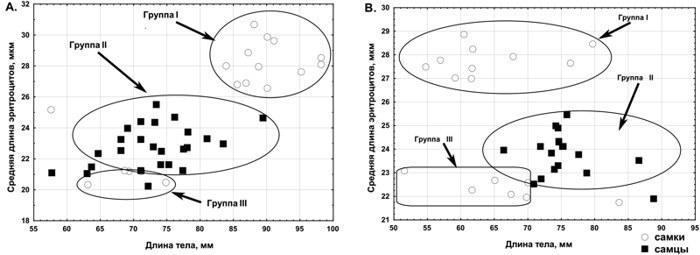
}{"translated_text":"Крім батьківських видів у регіоні дослідження зареєстровані ді-, три- і, у рідкісних випадках, тетраплоїдні гібриди. Серед точних способів визначення жаб, використаних у роботі, наймасовішою була проточна ДНК-цитометрія, яку виконували Ю. М. Розанов і С. М. Литвинчук (ЦІН РАН, Санкт‑Петербург). Цей метод базується на тому, що маса геному P. lessonae становить 6,92 пг, а геному P. ridibundus — 8,07 пг (рис. 4). Визначення ді‑ і триплоїдів зелених жаб підтверджено різними методами кариоаналізу. Склад представників Pelophylax esculentus complex, поширених на сході Лівобережно‑Дніпровської фізико‑географічної провінції, відповідає тому, який був виявлений іншими дослідниками в її західній частині (Морозов‑Леонов, 1998; Некрасова, Морозов‑Леонов, 2001; Некрасова, 2002). На території Середньоруської та Донечко‑Донської північно‑степної фізико‑географічних провінцій виявлені регіони масового поширення триплоїдних P. esculentus.\nБІОГЕОЦЕНТИЧНІ ТА ГЕОГРАФІЧНІ ФАКТОРИ, ЯКІ ВИЗНАЧАЮТЬ РОЗПРОСТРАНЕННЯ ПОПУЛЯЦІЙНИХ СИСТЕМ PELOPHYLAX ESCULENTUS COMPLEX У РЕГІОНІ ДОСЛІДЖЕННЯ\nДля визначення факторів, що впливають на склад популяційних систем зелених жаб, застосовано канонічний аналіз (пошук зв’язків між двома списками змінних: «лівим», що стосується наявності або відсутності певної форми в місці проживання, і «правим» набором ознак місця проживання). Біогеоценотичні ознаки місць проживання дозволяють пояснити близько 25 % дисперсії за складом представників зелених жаб. На рис. 5.А. показано розміщення змінних, використаних у скороченому варіанті канонічного аналізу з біогеоценотичними ознаками, на площині, заданій абсолютними (без урахування знаку) значеннями канонічних коренів. Біогеоценотичні ознаки, розташовані на цій площині близько до змінних, що вказують на наявність певних форм жаб, впливають на поширення цих форм.\n
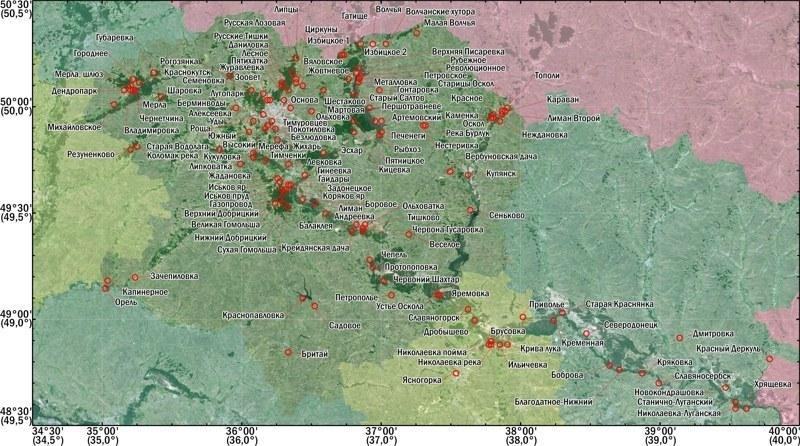
"translated_text": "У 2010-2011 рр. у описуваній ГПС з’явилися численні одно- і дворічні жаби. З 2010 року ми розпочали її дослідження методом маркування та повторного відлову (Популяційна ..., 2012). Склад цієї ГПС продовжує змінюватися. У 2012-2013 рр. кількість статево зрілих особин у складі нерестового стада в цій популяційній системі становила близько 500 особин, а у 2014 р. вона значно знизилася. Зареєстровано значний зріст частки тріплоїдних P. esculentus (p < 0,02) та зменшення частки самок (що становлять близько 20 % нерестового стада). Ймовірно, ГПС Іськова ставка не знайшла стабільного стану і знову опинилася в кризі (Meleshko et al, 2014). Спостереження за її майбутнім представляє особливий інтерес.\nЗ 2012 року розпочато маркування ще в двох ГПС: Нижній Добрицький та Жовтнєве (рис. 6). У ГПС зелених жаб, що населяють ставок у с. Жовтнєве, зареєстровано систему R‑E‑Epf‑типу, у якій тріплоїди представлені самками LRR і LLR. Результати експрес‑аналізу плоїдності за допомогою вимірювання розміру еритроцитів відповідають результатам проточної ДНК‑цитометрії. Ця ГПС розташована на межі IV та III субрегіонів і демонструє можливість часткової втрати геномів, що призводить до появи тріплоїдів.\n
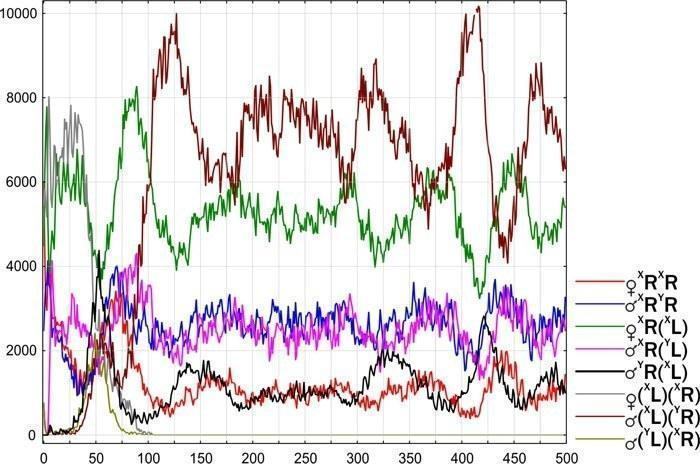
}Клітинний рівень відбору. Встановлено, що гаметогенез у P. esculentus значно менш стійкий, ніж у P. ridibundus. У сім’яниках самців P. esculentus виявлені клітини з кариотипом, відсутнім у спермі (Михайлова та ін., 2011; Кечеджи та ін., 2011). Це свідчить про наявність механізму елімінації клітинних ліній, розвиток яких не привів до формування повноцінних гамети.
Організменний рівень відбору. Порушення сперматогенезу у гібридів першого покоління відомі давно (Berger, 1971), але зареєстрована нами ступінь поширеності таких порушень виявилась неочікуваною (Нарушення .., 2014). Порушення фертильності виявлені у 85 % P. esculentus (n = 46) і 29 % P. ridibundus (n = 17) Північно‑Донецького центру різноманіття зелених жаб.
Скелетохронологічні дослідження демонструють різниці особин за швидкістю зростання, часом настання статевої зрілості та тривалістю життя. У різноманітті внутрі‑популяційних онтогенетичних стратегій зелених жаб (і ряду інших груп) можна припустити існування двох типових випадків, швидкорослості та повільнорослості (Внутріпопуляційні ..., 2014).
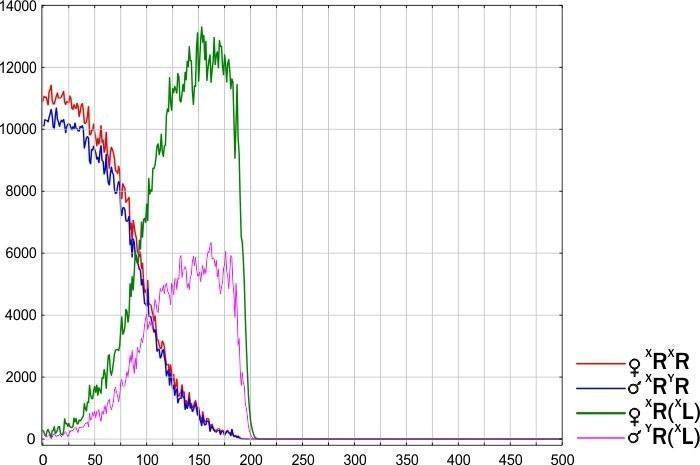
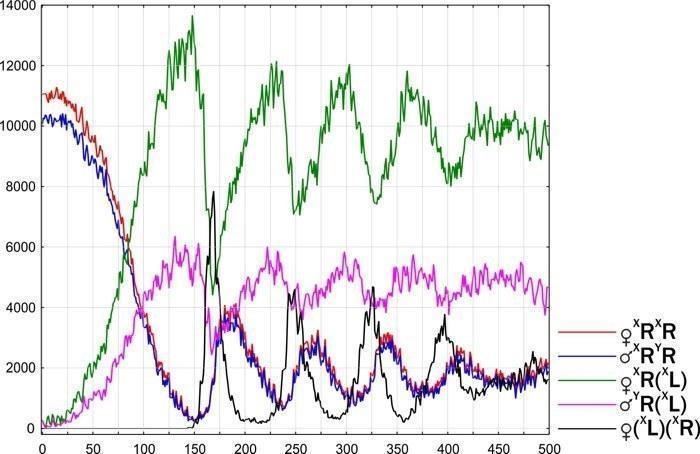
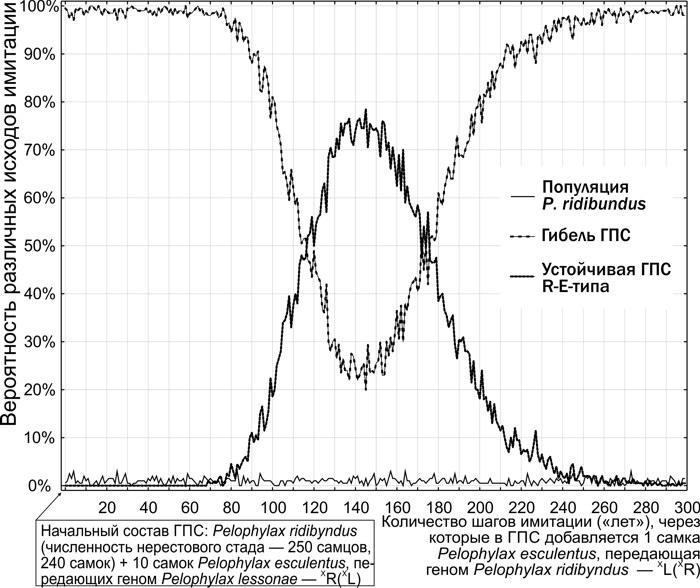
Груповий рівень відбору. Неоднакова стійкість різних типів ГПС — результат моделювання, що відповідає спостереженням. Різні типи ГПС гинуть, змінюються або перебувають у стійкому стані. Вони можуть «розмножуватись» (розселюватись), передаючи певний набір клональних геномів.
Філогенетичний рівень відбору. Відомо три шляхи подолання гібридної стерильності, що дозволяють зберегти гібридну композицію геномів (Shabanov, 2006): перехід до клонального відтворення (від вегетативного розмноження до партеногенезу); поліплоїдія; геміклональність. З них саме геміклональне успадкування вимагає дії складного кариогенетичного механізму, що забезпечує елімінацію та ендоредуплікацію геномів. Оскільки причиною доцільності біосистем є відбір, необхідно встановити, відбір якого рівня підтримує геміклональне успадкування.
У ряді робіт показано, що внаслідок порушень геміклональності при відтворенні гібридів виникають часткові рекомбіна́ти, які при схрещуванні з представниками батьківських видів можуть переносити у їх генофонди фрагменти генетичної інформації іншого батьківського виду (Наследование ... 2009; The ploidy ... 2010 та ін. роботи). Це прояви гомологічної рекомбінації (табл. 5), тобто рекомбінації між відповідними хромосомами, що належать різним видам. Ми висловили припущення, що обмежена (рівнем порушень при передачі клонального геному) міжвидова гомологічна рекомбінація дає еволюційну перевагу тим видам, схрещування яких призводить до появи геміклональних гібридів. Це пояснення відповідає концепції відбору видів (Eldredge, Gould, 1972), яка була доведена у ряді випадків (Goldberg et al., 2010). Ми припускаємо, що здатність до схрещування з утворенням геміклональних гібридів у роді Pelophylax підтримується відбором видів. Цим пояснюється швидка еволюція роду та наявність у ньому кількох гібридогенних комплексів (Plötner, 2005).
**ЗАКЛЮЧЕННЯ**
Обговорюються перспективи подальших досліджень ГПС зелених жаб Північно‑Донецького центру різноманіття Pelophylax esculentus complex.
**ВИСНОВКИ**
1. На території Лівобережної лісостепі України представники гібридогенного комплексу Pelophylax esculentus існують не лише у складі популяцій батьківських видів Pelophylax lessonae (Camerano, 1882) і Pelophylax ridibundus (Pallas, 1771), а й у складі популяційних систем, що включають диплоїдних і, у ряді випадків, поліплоїдних міжвидових гібридів Pelophylax esculentus (Linnaeus, 1758). На склад і динаміку таких систем впливають біоекзеноценотичні особливості місць існування, специфіка геміклонального відтворення Pelophylax esculentus та обмежене поширення окремих форм Pelophylax esculentus у річкових басейнах регіону дослідження.
2. У регіоні дослідження виділено п’ять субрегіонів, що відрізняються складом поширених у них популяційних систем і популяцій представників Pelophylax esculentus complex. Популяційні системи, що включають Pelophylax esculentus, зустрічаються у чотирьох з цих субрегіонів, триплоїди Pelophylax esculentus зареєстровані у двох з них.
3. Усередині виділених субрегіонів склад популяційних систем Pelophylax esculentus complex визначається біоекзеноценотичними умовами, передусім — характером рослинності навколо нерестового водоймища та рівнем антропогенної трансформації території. Різниці субрегіонів частково пов’язані з різницею характерних для них біоекзеноценотичних умов, а частково — з ізоляцією басейнів різних річок.
4. Для двох субрегіонів, розташованих у водозбірному басейні річки Північний Донець, зафіксовано унікально високе різноманіття представників Pelophylax esculentus complex, а також незвичний, у порівнянні з навколишніми територіями, склад їх популяційних систем. Цей регіон названо Північно‑Донецьким центром різноманіття Pelophylax esculentus complex.
5. Унікальність Північно‑Донецького центру різноманіття Pelophylax esculentus complex пов’язана з наступними його особливостями:
— на частині його території широко поширені триплоїдні Pelophylax esculentus обох статей з генотипами обох можливих геномних композицій (LLR і LRR);
— у ньому переважають ГПС R‑Ep‑ та R‑E‑типу;
— у ньому відсутні статево зрілі Pelophylax lessonae; особи Pelophylax lessonae, що виникають у результаті гибридолізу (схрещування гібридів), гинуть на ранніх етапах онтогенезу;
— його характеризує висока частка особин Pelophylax esculentus з гібридною амфіспермією (виробництвом статевих клітин обох батьківських видів);
— значна частина особин Pelophylax esculentus демонструє порушення фертильності.
6. Характерні властивості біосистем різних рівнів організації тісно пов’язані зі способом рекомбінації (на генному рівні), гаметогенезу або подібних процесів (на клітинному рівні), розмноження (на організменному рівні) і популяційного відтворення. Геміклональне успадкування у Pelophylax esculentus пов’язане з глибокою зміною всієї ланцюжка цих процесів і породжує специфічні біосистеми різних рівнів організації, що відрізняються від такових у організмів із типічним статевим розмноженням.
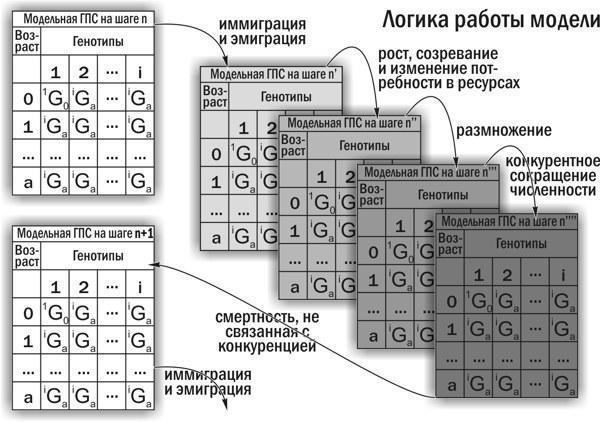
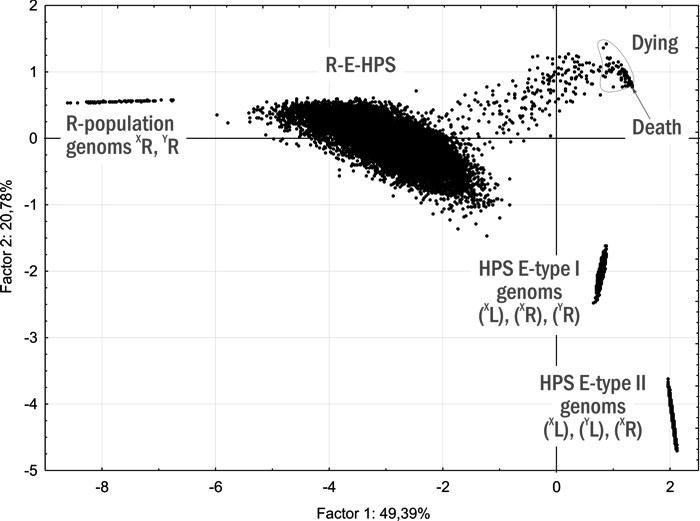
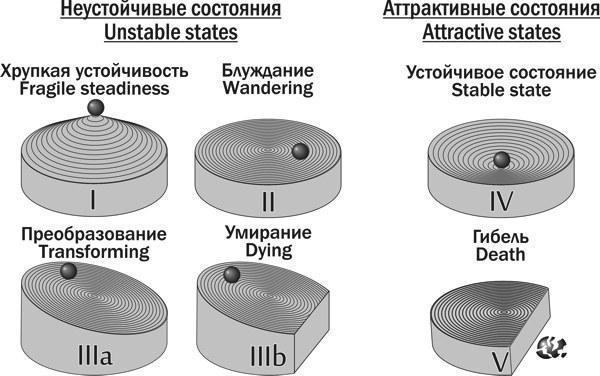
7. Існування міжвидових гібридів Pelophylax esculentus, у зв’язку з особливостями їх відтворення, можливо не в складі популяцій, а в складі геміклональних популяційних систем (ГПС). ГПС гібридогенних комплексів видів є особливою категорією біосистем, принципово відмінною від популяцій (тими, що складаються з особин різних видів і міжвидових гібридів, а також тим, що у них передаються не лише рекомбіантні, а й клональні геноми), а також від гільдій і спільнот (тими, що об’єднані спільним відтворенням). ГПС характеризуються не лише статевою та віковою структурою, а й складом кариогенетичних форм (груп особин, які відрізняються за геномною композицією та складом геномів, що передаються в гаметах), що визначає склад клонально передаваних геномів. ГПС здатні до трансформацій, тобто зміни складу і співвідношення різних кариогенетичних форм у їхньому складі.
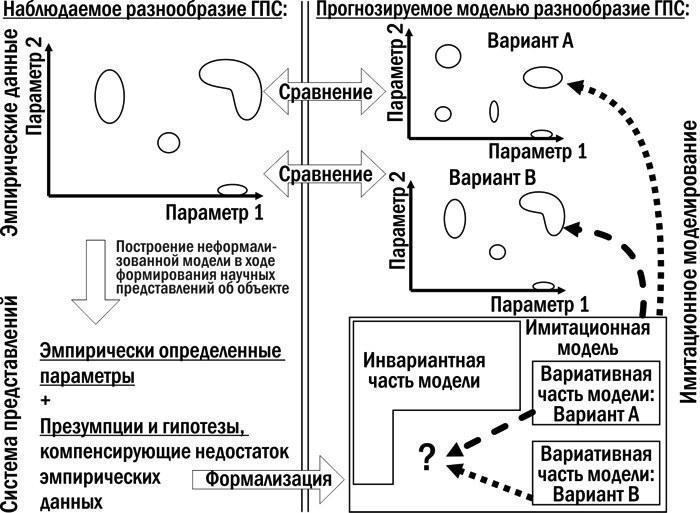
8. Для дослідження механізму трансформацій ГПС Pelophylax esculentus complex розроблено та створено імітаційну модель. Ця модель дає можливість отримувати розподіли ймовірностей результатів трансформацій ГПС залежно від вихідного складу ГПС, а також прийнятих припущень щодо характеру успадкування різних геномів і відносної життєздатності різних кариогенетичних форм Pelophylax esculentus complex в умовах певного біоекзеноценозу.
9. Використання імітаційної моделі, створеної в роботі, дозволяє тестувати гіпотези, що стосуються невивчених аспектів функціонування ГПС, на відповідність комплексу накопичених емпіричних даних.
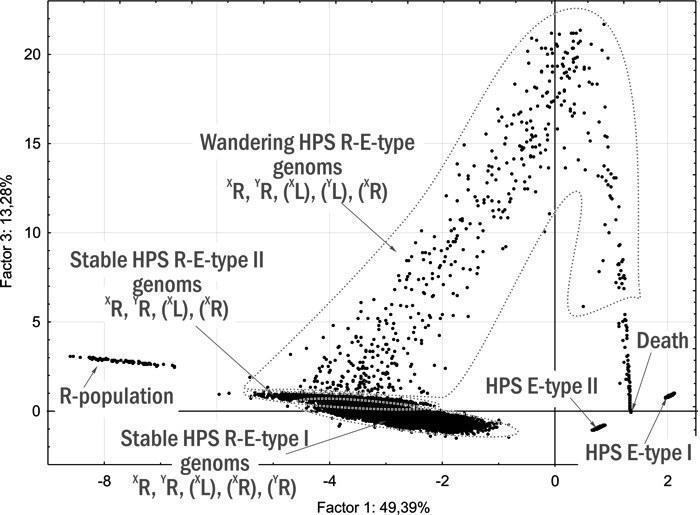
10. Розроблено динамічну (об’єднуючу розглянуті об’єкти в групи на основі прогнозу їх майбутньої динаміки) типологію ГПС, що включають диплоїдних представників Pelophylax esculentus complex. Показано, що такі ГПС можуть існувати лише у чотирьох стійких станах, у яких у їх склад входять представники Pelophylax esculentus. У всіх цих чотирьох стійких станах у ГПС передаються геноми обох батьківських видів.
11. Обґрунтовано теоретичну можливість існування геміклональних популяційних систем, що складаються виключно з диплоїдних Pelophylax esculentus. Показано, що ці ГПС будуть стійкими у випадку, якщо вони складатимуться з особин з гібридною амфіспермією, що належать до двох різних клонів (сукупностей генетично ідентичних особин).
12. Pelophylax esculentus complex надає виняткові можливості для вивчення багаторівневого відбору. У ньому зафіксовано відбір на кількох рівнях:
— генний, що відповідає моделі «егоїстичного гена» і проявляється у різній успішності поширення окремих алелей в популяціях або ГПС;
— геномний (пределний випадок генного, коли всі гени одного геному успадковуються як єдине ціле), що відповідає моделі «егоїстичного геному» і проявляється у конкуренції окремих геміклонів;
— клітинний, що відповідає моделі «егоїстичної лінії зародкових клітин» і проявляється у конкуренції різних ліній зародкових клітин у статевих залозах Pelophylax esculentus; формування життєздатних гамети завершує розвиток лише тих ліній, у яких успішно відбувається елімінація рекомбіантного геному та ендоредуплікація клонального (у випадку диплоїдних гібридів);
— організменний, що відповідає моделі «егоїстичного організму» і проявляється у чистому вигляді у конкуренції гібридних особин всередині клону;
— груповий, що відповідає моделі «стабільної ГПС» і проявляється у різниці ймовірності виживання та розселення ГПС різного складу.
Динаміка змін кожної окремої ГПС визначається взаємодією ефектів відбору на різних рівнях.
13. Висунуто гіпотезу, що пояснює феномен гібридної амфіспермії конкуренцією в клітинах зародкового шляху клональних геномів, змінених внаслідок відбору на здатність викликати елімінацію альтернативних геномів.
14. Висунуто гіпотезу, згідно з якою здатність двох відносно близьких видів до геміклональної гібридизації є адаптацією до гомологічної міжвидової рекомбінації, яка підтримується на рівні відбору видів.
**БЛАГОДАРНОСТІ**
Автор виконував дану роботу у співпраці з багатьма колегами та отримав цінну консультаційну і організаційну допомогу від численних фахівців. На різних етапах наукового шляху його наставляли V. P. Kudokotsev та V. M. Epstein, вдячну пам’ять про яких зберігає дисертант. Автор виражає вдячність V. L. Bulahov, який був керівником його кандидатської дисертації, та A. P. Rasnycynу, що надали цінні консультації під час підготовки докторської. Важливу роль у виконанні цієї роботи зіграла співпраця з S. N. Litvinchuk, Yu. M. Rosanov, L. Ya. Borkin, G. A. Lada та S. Yu. Morozov‑Leonov. Особливо важливою для автора була робота A. V. Korshunov та M. A. Kravchenko, які підготували свої кандидатські дисертації під його керівництвом. Всім названим і багатьом неназваним колегам автор висловлює щиру і глибоку подяку.
**ПУБЛІКАЦІЇ З ТЕМИ ДИССЕРТАЦІЇ**
Статті у виданнях, що входять до наукометричних баз даних
Наслідування батьківських геномів гібридною формою Rana “esculenta” (Amphibia, Ranidae) / S. Yu. Morozov‑Leonov, S. V. Mezhzherin, O. D. Nekrasova, D. A. Shabanov, A. V. Korshunov, F. F. Kurtyak // Генетика. – 2009. – Т. 45, № 4. – С. 488–495. (Дисертант брав участь у зборі матеріалу та інтерпретації результатів).
The ploidy and genetic structure of hybrid population of water frogs Pelophylax esculentus complex (Amphibia, Ranidae) of Ukraine fauna / S. V. Mezhzherin, S. Yu. Morozov‑Leonov, O. V. Rostovskaya, D. A. Shabanov, L. Yu. Sobolenko // Cytology and Genetics. – 2010. – Vol. 44, № 4. – P. 212–216. (Дисертант брав участь у плануванні зборів у Лівобережній лісостепі України, збирав матеріал і брав участь у інтерпретації результатів).
Influence of environmental conditions on the distribution of Central Asian green toads with three ploidy levels / S. N. Litvinchuk, G. O. Mazepa, R. A. Pasynkova, A. Saidov, T. Satorov, Yu. A. Chikin, D. A. Shabanov, A. Crottini, L. J. Borkin, J. M. Rosanov, M. Stöck // Journal of Zoological Systematics and Evolutionary Research. – 2011. – Vol. 49, № 3. – P. 233–239. (Дисертант брав участь у зборі матеріалу та інтерпретації результатів).
Кечеджи А. Е. Сперматогенез у триплоїдних Pelophylax esculentus (Amphibia, Anura) з Північно‑Донецького центру різноманіття зелених жаб (Україна, Харківська область) / A. E. Kechedzhi, O. V. Mikhailova, D. A. Shabanov // Вісник Харківського національного університету імені V. N. Kаразіна. Серія: біологія. – 2011. – Вип. 14, № 971. – С. 112–116. (Дисертант брав участь у постановці завдання для дослідження, виборі та опрацюванні методики, проведенні польових робіт, а також у інтерпретації результатів).
Бондарева A. A. Цитогенетичні особливості еритроцитів зелених жаб з Північно‑Донецького центру різноманіття Pelophylax esculentus complex / A. A. Bondareva, Yu. S. Bibik, S. M. Samilo, D. A. Shabanov // Вісник Харківського національного університету імені V. N. Kаразіна. Серія: біологія. – 2012. – Вип. 15, № 1008. – С. 116–123. (Дисертант брав участь у постановці завдання для дослідження, виборі та опрацюванні методики, проведенні польових робіт і в інтерпретації результатів).
Cytological maps of lampbrush chromosomes of European water frogs (Pelophylax esculentus complex) from the Eastern Ukraine / D. Dedukh, G. Mazepa, D. Shabanov, Ju. Rosanov, S. Litvinchuk, L. Borkin, A. Saifitdinova, A. Krasikova // BMC Genetics. – 2013. – Vol. 14. – P. 26–46. (Дисертант брав участь у зборі матеріалу і в інтерпретації результатів).
Внутріпопуляційні онтогенетичні стратегії швидкорослості та повільнорослості: визначення на прикладі безхвостих амфібій / D. A. Shabanov, A. V. Korshunov, E. V. Melesko, A. V. Shabanova, E. E. Usova // Вісник Харківського національного університету імені V. N. Kаразіна, серія «Біологія». – 2014. – Вип. 22, № 1126. – С. 115–124. (Дисертант брав участь у постановці завдання для дослідження, аналізі накопиченого матеріалу і в інтерпретації результатів).
Meleshko O. V. The study of three hemiclonal population systems Pelophylax esculentus complex from the Seversko‑Donetskiy center of green frogs diversity / O. V. Meleshko, O. V. Korshunov, D. A. Shabanov // Вісник Харківського національного університету імені V. N. Kаразіна. Серія «Біологія». – 2014. – Вип. 20, № 1100. – С. 153–158. (Дисертант брав участь у постановці завдання для дослідження, виборі та опрацюванні методики, проведенні польових робіт і в інтерпретації результатів).
Статті у профільних виданнях України та інших країн
Mass occurrence of polyploid green frogs (Rana esculenta complex) in Eastern Ukraine / L. J. Borkin, A. V. Korshunov, G. A. Lada, S. N. Litvinchuk, J. M. Rosanov, D. A. Shabanov, A. I. Zinenko // Russian Journal of Herpetology. – 2004. – Vol. 11, № 3. – P. 194–213. (Дисертант брав участь у зборі матеріалу і в інтерпретації результатів).
Різноманіття зелених жаб (Rana esculenta complex) у Харківській області: морфологічний аспект вивчення / O. V. Korshunov, T. V. Babinich, O. I. Zinenko, D. A. Shabanov // Біологія та валеологія. – 2004. – Вип. 6. – С. 24–30. (Дисертант поставив завдання для дослідження, брав участь у обробці літературних даних і в інтерпретації результатів).
Kravchenko M. A. Оцінка унікальності популяцій Bufo viridis Laurenti, 1768 (Amphibia, Anura) з точки зору раціональної екологічної етики / M. A. Kravchenko, D. A. Shabanov // Вісник Дніпропетровського університету. – 2005. – № 3/2. – С. 91–95. (Дисертант поставив завдання для дослідження, надав емпіричний матеріал для його виконання і брав участь в інтерпретації результатів).
Mazepa G. O. Формалізований опис рострального рисунку зелених ропух (Bufo viridis) / G. O. Mazepa, D. A. Shabanov // Біологія та валеологія. – 2005. – Вип. 7. – С. 77–83. (Дисертант поставив завдання для дослідження, брав участь у виборі та опрацюванні методики, зборі матеріалу, а також в інтерпретації результатів).
Вивчення популяційних систем зелених жаб (Rana esculenta complex) у Харківській області: історія, сучасний стан і перспективи / D. A. Shabanov, A. I. Zinenko, A. V. Korshunov, M. A. Kravchenko, G. A. Mazepa // Вісник Харківського національного університету імені V. N. Kаразіна. Серія: біологія. – 2006. – Вип. 3, № 729. – С. 208–220. (Дисертант поставив завдання для дослідження, брав участь у обробці літературних і емпіричних даних польових досліджень і в інтерпретації результатів).
До реорганізації зонування Національного природного парку «Гомільшанські ліси» / O. Yu. Akulov, T. A. Atemasova, O. F. Bartenyev, S. G. Viter, A. S. Vlaschenko, O. I. Zinenko, O. V. Korshunov, N. B. Saidakmedova, Ye. V. Skorobogotov, D. A. Shabanov, A. Yu. Utyevskyi // Заповідна справа в Україні. – 2006. – Т. 12, Вип. 2. – С. 73–79. (Дисертант брав участь у зборі матеріалу і формулюванні висновків роботи в частині, що стосується зелених жаб).
Еволюційно‑генетичні аспекти півклонального відтворення гібридної форми Rana kl. esculenta (Amphibia, Ranidae) / S. V. Mezhzherin, S. Yu. Morozov‑Leonov, O. D. Nekrasova, F. F. Kurtyak, D. A. Shabanov, A. V. Korshunov // Науковий вісник Ужгородського університету: Серія: біологія. – 2007. – Вип. 21. – С. 79–84. (Дисертант брав участь у зборі матеріалу і в інтерпретації результатів).
A case of natural triploidy in European diploid green toad (Bufo viridis), with some distributional records of diploid and tetraploid toads / L. J. Borkin, D. A. Shabanov, O. V. Brandler, O. V. Kukushkin, S. N. Litvinchuk, G. A. Mazepa, J. M. Rosanov // Russian Journal of Herpetology. – 2007. – Vol. 14, № 2. – P. 121–132.(Диссертант брав участь у зборі матеріалу, його обробці та інтерпретації результатів).
Про південну межу поширення гібридної Rana esculenta (Ranidae, Anura, Amphibia) на території України та Молдови: дані проточної ДНК-цитометрії / Л. Я. Боркін, О. С. Безман‑Мосейко, Г. А. Мазепа, А. І. Зіненко, А. В. Коршунов, Г. А. Лада, Д. А. Шабанов, С. Н. Литвинчук, Ю. М. Розанов // Праці Українського герпетологічного товариства. – 2008. – № 1. – С. 3–8. (Диссертант брав участь у зборі матеріалу та інтерпретації результатів).
Маро А. Н. Чи можуть умови розвитку головастиків Bufo bufo визначати темпи постметаморфічного росту та дозрівання жаб? / А. Н. Маро, А. В. Шабанова, Д. А. Шабанов // Питання герпетології. Мат‑ли III з’їзду Герпетологічного товариства ім. А. М. Нікольського. – СПб: 2008. – С. 274–280. (Диссертант поставив завдання для дослідження, брав участь у обробці даних та інтерпретації результатів).
Кравченко М. А. Можливі шляхи трансформації популяційних систем Pelophylax esculentus complex (Ranidae, Anura, Amphibia) / М. А. Кравченко, Д. А. Шабанов // Праці Українського герпетологічного товариства. – № 1, 2008. – С. 15–20. (Диссертант брав участь у постановці завдання для дослідження, обробці емпіричних даних та інтерпретації результатів).
Шабанов Д. А. Які ж зелені жаби населяють Харківську область? Термінологічний і номенклатурний аспекти проблеми / Д. А. Шабанов, О. В. Коршунов, М. О. Кравченко // Біологія та валеологія. – 2009. – Вип. 11. – С. 116–125. (Диссертант поставив завдання для дослідження, брав участь у обробці літературних даних та інтерпретації результатів).
Коршунов А. В. Екологічні фактори, що впливають на поширення представників Pelophylax esculentus complex у Харківській області / А. В. Коршунов, Д. А. Шабанов // Біологічний вісник. – 2009. – Т. 13, № 1–2. – С. 76–83. (Диссертант брав участь у постановці завдання для дослідження, виборі та відпрацюванні методики, зборі матеріалу, а також інтерпретації результатів).
Кравченко М. А. Моделювання трансформацій геміклональних популяційних систем зелених жаб (Pelophylax esculentus complex; Amphibia, Ranidae) за допомогою рекурентних різницевих рівнянь / М. А. Кравченко, Д. А. Шабанов // Вісник Харківського національного університету імені В. Н. Каразіна. Серія: біологія. – 2010. – Вип. 12, № 920. – С. 70–82. (Диссертант брав участь у постановці завдання для дослідження, виборі методів його виконання та інтерпретації результатів).
Бондарєва А. А. Чи впливають розмірні ефекти клітин диплоїдних та триплоїдних їстівних жаб на їх стійкість до гіпоксії? / А. А. Бондарєва, Т. І. Махній, Д. А. Шабанов // Біологія та валеологія. – 2010. – Вип. 12. – С. 10–15. (Диссертант поставив завдання для дослідження, брав участь у виборі та відпрацюванні методики, а також інтерпретації результатів).
Михайлова О. В. Як оцінити популяційне навантаження, пов’язане з геміклональною гібридизацією в популяційних системах Pelophylax esculentus complex? / О. В. Михайлова, О. Є. Усова, Д. А. Шабанов // Біологія та валеологія. – 2011. – Вип. 13. – С. 44–50. (Диссертант поставив завдання для дослідження, брав участь у відпрацюванні методики та інтерпретації результатів).
Дослідження стійкості геміклональних популяційних систем гібридного комплексу зелених жаб за допомогою імітаційного моделювання / М. А. Кравченко, Д. А. Шабанов, М. В. Владимирова, Г. Н. Жолткевич // Вісник Дніпропетровського університету. Біологія. Екологія. – 2011. – Т. 1, Вип. 19. – С. 51–64. (Диссертант брав участь у постановці завдання для дослідження, виборі та відпрацюванні методики, а також інтерпретації результатів).
Михайлова О. В. Вивчення сперматогенезу у диплоїдних Pelophylax esculentus (Amphibia, Anura) за допомогою кариоаналізу в розчавлених препаратах / О. В. Михайлова, А. Є. Кечеджи, Д. А. Шабанов // Праці Українського герпетологічного товариства. – 2011. – № 3. – С. 120–127. (Диссертант брав участь у постановці завдання для дослідження, виборі та відпрацюванні методики, а також інтерпретації результатів).
Шабанов Д. А. Гібридогенний комплекс зелених жаб як модель для вивчення багаторівневого добору / Д. А. Шабанов // Вісник Прикарпатського національного університету імені Василя Стефаника. Серія – Біологія. – 2012. – Вип. XVII. – С. 90–94.
Популяційна система зелених жаб (Pelophylax esculentus complex) Іськова ставка в Зміївському районі Харківської області: історія вивчення та оцінка чисельності / Е. В. Мелешко, А. Д. Суворова, М. А. Кравченко, Д. А. Шабанов // Біологічний вісник. – 2012. – Т. 14, № 1. – С. 63–68. (Диссертант поставив завдання для дослідження, брав участь у виборі та відпрацюванні методики, а також інтерпретації результатів).
Вивчення флуктуючої асиметрії річкового окуня (Perca fluviatilis L., 1758) / К. П. Виноградова, Ю. В. Сакун, К. М. Белоусова, Г. Л. Гончаров, Д. А. Шабанов // Біологія та валеологія. – 2012. – Вип. 14. – С. 9–17. (Диссертант поставив завдання для дослідження, брав участь у відпрацюванні методики, а також інтерпретації результатів).
Бондарєва А. А. Порівняння деяких гематологічних показників диплоїдних та триплоїдних Pelophylax esculentus / А. А. Бондарєва, К. В. Сєдова, Д. А. Шабанов // Праці українського герпетологічного товариства. – 2013. – № 4. – С. 22–26. (Дисертант поставив завдання для дослідження, збирав матеріал та брав участь в інтерпретації результатів) (Диссертант поставив завдання для дослідження, збирав матеріал та брав участь в інтерпретації результатів).
Порушення фертильності у міжвидових гібридів зелених жаб із Сіверсько‑Донецького центру різноманіття Pelophylax esculentus complex / А. А. Боброва, Р. М. Макарян, В. П. Шейко, Д. А. Шабанов // Біологія та валеологія. – 2014. – Вип. 16. – С. 7–15. (Диссертант поставив завдання для дослідження, брав участь у відпрацюванні методики, зборі матеріалу, а також інтерпретації результатів).
Статті в інших виданнях
Масова поліплоїдія в гібридному комплексі Rana esculenta (Ranidae, Anura, Amphibia) на Сході України / Л. Я. Боркін, А. І. Зіненко, А. В. Коршунов, Г. А. Лада, С. Н. Литвинчук, Ю. М. Розанов, Д. А. Шабанов // Матеріали Першої конференції Українського Герпетологічного Товариства. – К.: Зоомузей ННПМ НАНУ, 2005. – С. 23–26.
Маро А. Н. Механізми розселення сірої жаби (Bufo bufo (L. 1758); Amphibia, Anura) та особливості її популяцій у нещодавно заселених місцях / А. Н. Маро, Д. А. Шабанов // Матеріали Першої конференції Українського Герпетологічного Товариства. – К.: Зоомузей ННПМ НАНУ, 2005. – С. 107–110.
Дослідження цілісних властивостей біосистем на прикладі імітаційного моделювання гібридних популяційних систем зелених жаб / М. В. Владимирова, Г. Н. Жолткевич, А. А. Луцик, Д. А. Шабанов // Вісник Харківського національного університету. Серія «Математичне моделювання. Інформаційні технології. Автоматизовані системи управління» – 2007. – № 780. – С. 61–70.
Шабанов Д. А. Регiональна звiтна, підручник, довідник i наукова монографія. Рецензія на книгу В. Л. Булакова, В. Я. Гассо, А. Е. Пахомова «Біологічне різноманіття України. Дніпропетровська область. Земноводні та рептилії (Amphibia et Reptilia)» / Д. А. Шабанов, А. І. Зіненко // Вісник Дніпропетровського університету. Біологія. Екологія. – 2008. – Вип. 16, т. 2. – С. 209–211.
Шабанов Д. А. Чому роздвоєні статеві організми витісняють крос‑заплідних гермафродитів: роздвоєність як рівновага за Нешем / Д. А. Шабанов // Мат. Міжнар. науков. конф. присв. 80‑річчю з дня народження проф. А. П. Крапивного. – Х.: ХНУ ім. В. Н. Каразіна, 2009. – С. 38–49.
Шабанов Д. А. Зелені жаби: життя без правил чи особливий спосіб еволюції / Д. А. Шабанов, С. Н. Литвинчук // Природа. – 2010. – № 3 (1135). – С. 29–36.
Vlasova T. Simulation of the Expanded Iterated Version of the Prisoner's Dilemma Game Recognisers / T. Vlasova, M. Vladymyrova, D. Shabanov // 7‑th Int. Conf. ICTERI, 2011. –Vol. 716. – P. 82–86.
Тези в матеріалах конференцій
Шабанов Д. А. Унікальність популяцій безхвостих амфібій як підстава для їх охорони / Д. А. Шабанов, А. В. Коршунов, М. А. Кравченко // Сучасні проблеми зоології та екології. – Одеса: Фенікс, 2005. – С. 328–329.
Кравченко М. А. До розробки методів оцінки цінності популяцій з точки зору раціональної екологічної етики / М. А. Кравченко, Д. А. Шабанов // Біорізноманіття і роль зооценозу в природних та антропогенних екосистемах: Мат. III Міжнар. науков. конф. – Дніпропетровськ: Видавництво ДНУ, 2005. – С. 78–80.
Коршунов А. В. Перетворення популяційних систем зелених жаб як особлива категорія процесів природного розвитку / А. В. Коршунов, М. А. Кравченко, Д. А. Шабанов // Сучасні проблеми популяційної екології. Мат. IX Міжнар. науково‑практ. еколог. конф. – Белгород: Видавництво ПОЛІТЕРРА, 2006. – С. 101–102.
Шабанов Д. А. Гібридні жаби Rana esculenta, що виробляють гамети обох батьківських форм, – результат незалежної клональної еволюції геномів? / Д. А. Шабанов // Матеріали конференції «Сучасні проблеми біологічної еволюції. До 100‑річчя Державного Дарвінського музею». – Москва, Видавництво ГДМ, 2007. – С. 199–201.
Кравченко М. А. Ієрархія рівнів біорізноманіття на прикладі гібридного Rana esculenta complex / М. А. Кравченко, Д. А. Шабанов // Zoocenosis‑2007. Біорізноманіття та роль тварин в екосистемах. – Дніпропетровськ, ДНУ, 2007. – С. 381–383.
Кравченко М. А. Що нового можна дізнатися про зелених жаб за допомогою імітаційного моделювання? (Пленарний доклад на відкритті конференції) / М. А. Кравченко, А. А. Луцик, Д. А. Шабанов // «Біологія: від молекули до біосфери». Матеріали III Міжнародної конференції молодих науковців. – Харків: СПД ФО Михайлова Г. Г., 2008. – С. 9–11.
Усова Е. Е. Про оптимізацію методики ретроспективної оцінки динаміки розмірів тіла представників Pelophylax esculentus complex (Amphibia, Ranidae) за допомогою скелетохронології / Е. Е. Усова, Д. А. Шабанов // Zoocenosis‑2009. Біорізноманіття та роль тварин в екосистемах. – Дніпропетровськ, ДНУ, 2009. – С. 278–280.
Матеріали про гельмінтологічну фауну зелених жаб (Rana esculenta complex) в Харківській області / М. В. Резванцева, Г. А. Лада, Д. С. Аксенов, Д. А. Шабанов, А. В. Коршунов, І. В. Чихляєв, Л. Я. Боркін, С. Н. Литвинчук, Ю. М. Розанов // Теоретичні та практичні проблеми паразитології. – Матеріали Міжнародної наукової конференції. – М., 2010. – С. 308–312.
Шабанов Д. А. До розробки класифікації типів популяційного відтворення / Д. А. Шабанов // Видові популяції та спільноти в антропогенних трансформованих ландшафтах: стан і методи діагностики. — Мат. XI Міжнар. науково‑практич. екологіч. конф. – Белгород, 2010. – С. 228–229.
Кравченко М. А. Рівні біорізноманіття Pelophylax esculentus complex / М. А. Кравченко, Д. А. Шабанов // Біорізноманіття і стале розвиток. Тези Міжнар. науково‑практ. конф. – Сімферополь: КНЦ, 2010. – С. 68–71.
Власова Т. Імітаційне моделювання розширеного ітерованого варіанту гри «дилема в’язня» / Т. Власова, М. Владимирова, Д. Шабанов // ICTERI 2011. ІКТ в освіті, дослідженнях та індустріальних додатках: інтеграція, гармонізація та трансфер знань. – Херсон: ХДУ, 2011. – С. 57–58.
Бондарева А. А. Порівняння розмірів і форми еритроцитів у диплоїдних і триплоїдних гібридів зелених жаб (Pelophylax esculentus) / А. А. Бондарева, Д. А. Шабанов // Всеукраїнський конкурс студентських наукових робіт з біологічних наук: матеріали підсумкової науково‑практичної конференції (Мелітополь, 29‑31 березн. 2011 р.). – Мелітополь: «Люкс», 2011. – С. 11–14.
Cytological maps of lampbrush chromosomes of European water frogs / D. Dedukh, G. Mazepa, D. Shabanov, S. Litvinchuk, L. Borkin, A. Saifitdinova, A. Krasikova // 18th International Chromosome Conference (Manchester, 29 August – 2 September 2011). Speaker abstracts. – P. 48–49.
АНОТАЦІЯ
Шабанов Д. А. Еволюційна екологія популяційних систем гібридогенного комплексу зелених жаб (Pelophylax esculentus complex) Лівобережного лісостепу України. — На правах рукопису.
Дисертація на здобуття наукового ступеня доктора біологічних наук за спеціальністю 03.00.16 — екологія. — Дніпропетровський національний університет імені Олеся Гончара. — Дніпропетровськ, 2015.
Наведено результати багаторічних досліджень Pelophylax esculentus complex у Лівобережному лісостепу України. У регіоні дослідження виділено п’ять субрегіонів, що відрізняються складом популяційних систем і популяцій представників Pelophylax esculentus complex, що поширені в них. Популяційні системи, що включають Pelophylax esculentus, зустрічаються в чотирьох із цих субрегіонів, триплоїди Pelophylax esculentus зареєстровані у двох із них. Описано Сіверсько‑Донецький центр різноманіття Pelophylax esculentus complex.
Геміклональне успадкування у Pelophylax esculentus є причиною появи геміклональних популяційних систем (ГПС). Для дослідження трансформацій ГПС Pelophylax esculentus complex створено імітаційну модель. За її допомогою розроблено динамічну типологію ГПС, що включають диплоїдних представників Pelophylax esculentus complex. Показано, що Pelophylax esculentus complex надає виняткові можливості для вивчення багаторівневого відбору. Запропоновано гіпотези, що пояснює феномен гібридної амфіспермії та пояснює добір на здатність до геміклональної гібридизації.
Ключові слова: екологія, еволюція, Pelophylax esculentus complex, P. ridibundus, P. lessonae, P. esculentus, геміклональні популяційні системи, імітаційне моделювання, багаторівневий добір, стійкість.
АННОТАЦІЯ
Шабанов Д. А. Еволюційна екологія популяційних систем гібридогенного комплексу зелених жаб (Pelophylax esculentus complex) Лівобережної лісостепи України. — На правах рукопису.
Дисертація на здобуття наукового ступеня доктора біологічних наук за спеціальністю 03.00.16 — екологія. — Дніпропетровський національний університет імені Олеся Гончара. — Дніпропетровськ, 2015.
Приведено результати багаторічних досліджень Pelophylax esculentus complex у Лівобережній лісостепі України. В регіоні дослідження виділено п’ять субрегіонів, що відрізняються за складом поширених у них популяційних систем і популяцій представників Pelophylax esculentus complex. Популяційні системи, що включають Pelophylax esculentus, зустрічаються в чотирьох з цих субрегіонів, триплоїди Pelophylax esculentus зареєстровані в двох з них. Різниця субрегіонів частково пов’язана з різницею характерних для них біогеоценотичних умов, а частково — з ізоляцією басейнів різних річок. Два субрегіони, розташовані в басейні річки Північний Донец, утворюють Північно‑Донецький центр різноманіття Pelophylax esculentus complex.
Показано, що характерні властивості біосистем різних рівнів організації тісно пов’язані зі способом рекомбінації, гаметогенезу, розмноження та популяційного відтворення. Геміклональне успадкування у Pelophylax esculentus пов’язане з глибокою зміною цієї ланцюгової послідовності процесів. Відтворення Pelophylax esculentus можливе лише у складі геміклональних популяційних систем (ГПС), що є особливою категорією біосистем.
Для дослідження механізму трансформацій ГПС Pelophylax esculentus complex створено імітаційну модель, що дозволяє тестувати гіпотези, що стосуються невивчених аспектів функціонування ГПС, на відповідність комплексу накопичених емпіричних даних. За її використанням розроблено динамічну (об’єднуючу розглянуті об’єкти в групи на основі прогнозу їх майбутньої динаміки) типологію ГПС, що включають диплоїдних представників Pelophylax esculentus complex. Обґрунтовано теоретичну можливість існування геміклональних популяційних систем, що складаються виключно з диплоїдних Pelophylax esculentus.
Показано, що Pelophylax esculentus complex надає унікальні можливості для вивчення багаторівневого відбору. У ньому зафіксовано прояви відбору на генному, геномному, клітинному, організмовому та груповому рівнях. Запропоновано гіпотезу, що пояснює феномен гібридної амфіспермії конкуренцією в клітинах зародкового шляху клональних геномів, змінених внаслідок відбору на здатність викликати елімінацію альтернативних геномів. Запропоновано гіпотезу, згідно з якою здатність двох відносно близьких видів до геміклональної гібридизації є адаптацією до гомологічної міжвидової рекомбінації, підтримуваною на рівні відбору видів.
Ключові слова: екологія, еволюція, Pelophylax esculentus complex, P. ridibundus, P. lessonae, P. esculentus, геміклональні популяційні системи, імітаційне моделювання, багаторівневий відбір, стійкість.
Annotation
Shabanov D. A. Evolutionary ecology of population systems of green frogs’ hybridogenetic complex (Pelophylax esculentus complex) from Left Bank forest‑steppes of Ukraine. — The Rights of the manuscript.
Dissertation for a degree of Doctor of Biological Sciences by speciality 03.00.16. – Ecology. – Oles’ Gonchar Dnipropetrovsk National University. – Dnipropetrovsk, 2015.
The results of long‑term research of Pelophylax esculentus complex in Left Bank forest‑steppes of Ukraine are represented. The 5 sub‑regions were distinguished in the study region. They differ in composition of widespread population systems and populations of Pelophylax esculentus complex representatives. Population systems, which include Pelophylax esculentus, were found in four of these regions, Pelophylax esculentus triploids were registered in two of them. The Seversko‑Donetskiy centre of green frogs’ diversity was described.{"translated_text":"Гіпотеза, що пояснює гібридну амфісемію та відбір за здатність до геміклональної гібридизації, була запропонована.\nКлючові слова: екологія, еволюція, комплекс Pelophylax esculentus, P. ridibundus, P. lessonae, P. esculentus, геміклональні популяційні системи, моделювання симуляції, багаторівневий відбір, стабільність."}