
Шишкин, 2010. Эволюционная теория и научное мышление
Шишкин М.А. Эволюционная теория и научное мышление. // Палеонтологический журнал, 2010. — № 6. — с. 3-17
Шишкин М.А. Еволюційна теорія та наукове мислення. // Палеонтологічний журнал, 2010. — № 6. — с. 3-17
УДК 576.12:001.8
ЕВОЛЮЦІЙНА ТЕОРІЯ ТА НАУКОВЕ МИСЛЕННЯ
© 2010 р. М. А. Шишкин
Палеонтологічний інститут імені А.А. Борисяка РАН e-mail: shishkin@paleo.ru Надійшло в редакцію 25.12.2009 р.
Прийнято до друку 29.01.2010 р.
Прогрес теоретичної думки та зростання фактичного знання в природничих науках не пов’язані прямою залежністю. Стан теорії надовго визначається особливостями колективного мислення, що склалися в даній галузі знань. Це особливо стосується концепції природного відбору, відомої у вигляді двох полярних інтерпретацій — генетичної теорії еволюції та епігенетичної теорії. Передбачуваний підсумок їх протистояння не буде пов’язаний з “вирішальними аргументами” на користь однієї з них. Його настання залежить від того, наскільки швидко еволюційна біологія звільниться від традиції мозаїчного мислення, що дозволяє об’єднувати взаємовиключні концепції. Ключовим у цьому плані буде усвідомлення несумісності ідеї корпускулярної детермінанції розвитку з принципом системної обумовленості того ж процесу.
## ВСТУП
Прогрес теоретичної думки та зростання фактичного знання в природничих науках узгоджуються один з одним лише в загальних рисах. Більше того, екстенсивне накопичення емпіричних спостережень може відкинути теоретичну думку назад. На конкретних історичних етапах вирішальними для стану теорії можуть бути зовсім інші фактори. Перш за все, це особливості та традиції колективного наукового мислення, що склалися в даній галузі знань. Усе це стосується і теорії біологічної еволюції на основі природного відбору.
У дарвінському викладі теорії її базові поняття (відбір, змінність і успадкування) слабо конкретизовані, що було природно при тодішньому рівні біологічних уявлень і відсутності у Дарвіна схильності до умозричних конструкцій. Йому багато в чому було лише намічено те, що стало усвідомленим значно пізніше. Особливо відзначимо близькість творця теорії до інтуїтивного розуміння ролі природного відбору як механізму стабілізації історичних змін: “відбір у цих випадках ще не встиг опанувати схильності до подальшої змінності…”; або: “…відбір вже встиг повідомити… органу постійний характер” (см. Дарвін 2003, с. 166). Звідси ж логічно випливало і пояснення ним змінності рудиментарних органів як результату відсутності селективного контролю (там же, с. 165). Глибинний сенс цих узагальнень не міг бути тоді зрозумілим через відсутність уявлень про системні процеси.
Далі дарвінська теорія отримала дві діаметрально полярні інтерпретації, спочатку пов’язані з іменами А. Вайсманна (неодарвінізм, або генетична теорія еволюції — ГТЕ) та І.І. Шмальгаузена—К. Воддінгтона (вчення про стабілізуючий відбір, або епігенетична теорія — ЕТЕ). Вони спираються на альтернативні концепції індивідуального розвитку, тобто, відповідно, на преформаційну (прямо або за замовчуванням) і епігенетичну моделі. Перша з них передбачає ту чи іншу форму упорядкованого відповідності між елементами зародкової клітини та частинами дорослого організму. Навпаки, у рамках другої моделі ці категорії належать до різних ієрархічних рівнів системи розвитку і не пов’язані прямими залежностями.
У уявленнях ГТЕ субстратом відбору є мозаїка незалежних успадкованих факторів (генів) у статевих клітинах. Стійкість (успадкованість) ознак дорослого фенотипу є виразом іманентних властивостей цих факторів, що не вимагає еволюційного пояснення. Еволюція організму і його онтогенезу є результатом відбору генів; тому вона протікає, як і сам онтогенез, у напрямку від зиготи до дорослого стану. Успадковані зміни (мутації) і природний відбір (у ролі ситечка для них) взаємодіють в еволюції як незалежні фактори. Успадковані та неуспадковані зміни фенотипу якісно різні.
Основа цього світогляду наочно виражена в словах Вайсманна: “Природний підбір лише на вигляд має справу з якісними особливостями готового організму; насправді ж лише зі схованими в статевих клітинах зачатками цих особливостей” (Weissmann, 1883; цит. за Філіпченко, 1977, с. 148).
Навпаки, для ЕТЕ об’єктом еволюції є індивідуальний розвиток як ціле, тобто онтогенез розглядається як динамічна система, спрямована до кінцевого рівноваги (дорослої норми). Еволюція — це ланцюг актів репарації такої системи, що слідують після кожного порушення її стійкості. Відбір — це механізм самоорганізації, тобто пошуку порушеною системою нового рівноваги шляхом стабілізації одного з реалізованих відхилень розвитку. Матеріал відбору — це гетерогенні особи-ізоаберанти, а не носії “відбираної мутації”. Еволюційні зміни починаються з дорослого фенотипу і поширюються у напрямку генотипу наступних поколінь¹. Успадкованість (стійкість) — не партнер природного відбору, а його продукт, що виступає як системна властивість розвитку. Усі зміни фенотипу є виразами норми реакції і в цьому якості однорідні.
Ці принципи відображені, зокрема, у наступних узагальненнях: “Хоча зміна генотипу і є необхідною базою для еволюційного процесу, проте не вона визначає еволюцію… Навпаки, еволюція організму визначає зміни його генотипу…” (Шмальгаузен, 1940, с. 57). “Теорія природного відбору є … теорією, що пояснює виникнення та перетворення успадкованих механізмів. …Стійкість… ознаки… не властивість гена, а вираз взаємозалежності… частин … розвиваючогося організму” (Шмальгаузен, 1982, с. 110, 174) (курсив наш — МШ).
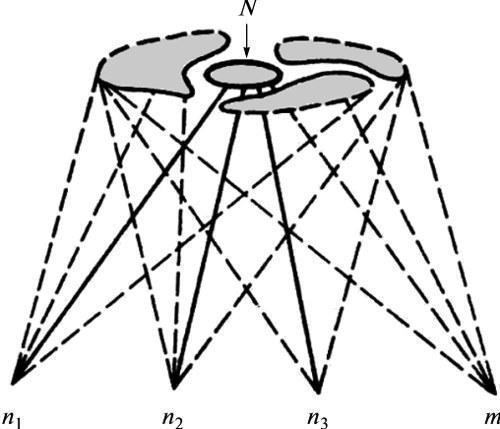
Згідно з ЕТЕ, простір можливих аберацій розвитку в даній системі становить її видоспецифічну характеристику. Будь‑яке відхилення від нормального підсумку розвитку залишається в межах цього обмеженого простору і представляє цілісний відповідь системи на порушення належної координації подій у ній (рис. 1). Він, таким чином, відображає властивості самої системи, а не специфіку того чи іншого внутрішнього або зовнішнього пошкоджувального фактора (ср. Goldschmidt, 1938, 1940). У сумі цей простір реагування описується як сукупність онтогенетичних траєкторій, здійснених у даному типі розвитку з тією чи іншою ймовірністю. Його наочна тривимірна модель — епігенетичний ландшафт К. Воддінгтона у вигляді мережі дивергентних долин, що позначають можливі варіанти розвитку (рис. 2, а). Демонстрована цією моделлю відносно проста структура “є властивістю вищого порядку, заснованою на підкладальній мережі значно складніших взаємодій” (Waddington, 1957, с. 34, рис. 5), тобто процесів, що протікають на рівнях генної експресії та елементарних морфогенетичних актів (рис. 2, б).
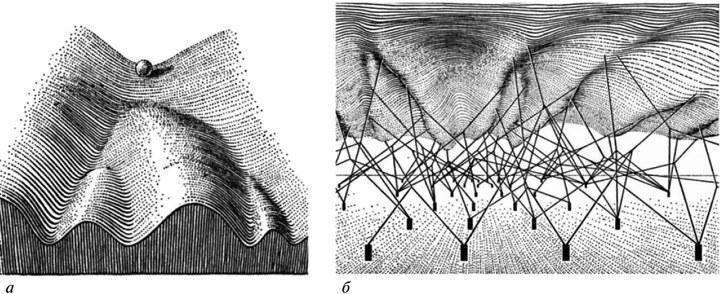
З сказаного ясно, що в уявленнях ЕТЕ не існує фенотипічних новацій, привнесених мутаціями. “Природний відбір… створює епігенетичний ландшафт, який… робить можливими фенотипічні ефекти мутацій. У цьому світлі звичне твердження, що сировина еволюції створюється випадковими мутаціями, є порожнім” (Waddington, 1957, с. 188). Іншими словами, “мутаційні” зміни фенотипу реалізують лише ті чи інші з існуючих можливостей реагування, властивих даній системі розвитку (Шишкін, 2006). Саме ж еволюційна зміна такої системи, згідно з ЕТЕ, означає перебудову її епігенетичного ландшафту, тобто створення в ньому нової каналізованої траєкторії і, відповідно, зміну загального спектру аберацій розвитку (рис. 3).
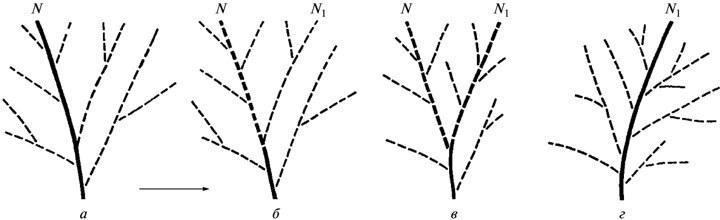
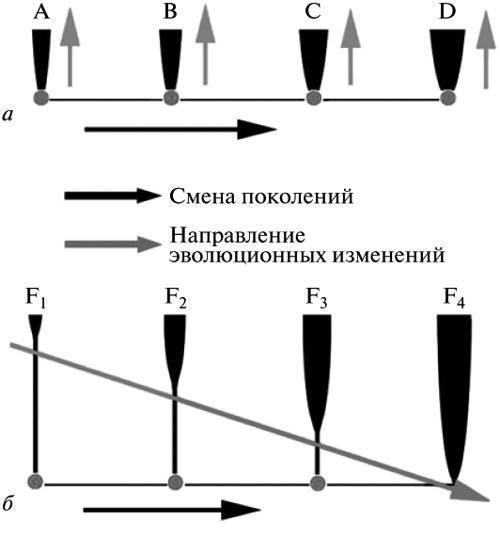
Суть різниці між двома розглянутими альтернативами дарвінізму може бути виражена наступним чином. Для ГТЕ (рис. 4, а) кожна елементарна еволюційна подія є продуктом одиничного онтогенетичного циклу, реалізованого на основі зміненої (мутантної) зародкової клітини. Таким чином це за визначенням сальтаціоністська теорія. Навпаки, для ЕТЕ (рис. 4, б) кожна еволюційна подія — це завжди процес, що охоплює чергу поколінь, де початкова фенотипічна відхилення проявляється раніше, ніж формується стабілізований (успадковано‑стійкий) механізм його здійснення.
Кінцевий підсумок протистояння цих концепцій здається цілком передбачуваним, проте основою для нього очевидно не буде подання “вирішальних аргументів”, переконливих для обох сторін. Доля теорій, як і обставини їх виникнення, зазвичай визначається інакше (см. розділ VI). Розглянемо деякі суттєві в цьому випадку фактори.
I. ТЕОРІЇ ТА ІНТЕРПРЕТАЦІЯ ФАКТІВ
Поява теорії — евристичний акт, що не зводиться до логічних операцій. Теорії не будуються безпосередньо з фактів; навпаки, останні отримують освітлення у світлі теорії (Любищев, 1925). Від вибору постулатів теорії (не обов’язково усвідомленого) залежить, які факти для неї суттєві та передбачувані, а які належать до інформаційного шуму, що затемнює належний хід подій. Наприклад, усі основні “шуми” ГТЕ — це саме область передбачуваних явищ для ЕТЕ (табл. 1).
Таблиця 1. Феномени, передбачувані для ЕТЕ, та можливості їх інтерпретації в рамках ГТЕ
| Феномени, передбачувані для ЕТЕ | Можливості їх інтерпретації в рамках ГТЕ |
|---|---|
| Стійкість нормального (“дикого”) фенотипу | “генетичний гомеостаз”? |
| Гетерогенність природних популяцій | “частотозалежний відбір”? |
| Переважає не‑менделівське успадкування аномалій у популяціях | вплив “генотипічного середовища”? |
| Можливість зміни “генетичної детермінації” ознаки | ? |
| Паралелізм мутаційної та модифікаційної змінності | ?Той самий вибір постулатів передбачає і робочі поняття (мова) теорії. Поза нею вони або позбавлені змісту, або можуть мати зовсім інше значення. Наприклад, поняття відбору мутацій або генетичного дрейфу позбавлені сенсу поза межами ГТЕ; і, навпаки, уявлення про стабілізацію розвитку для цієї теорії є безпредметним. Подібним чином природний відбір означає для двох версій дарвінізму діаметрально різні речі: для ГТЕ це сито для мутацій, а для ЕТЕ — механізм перетворення цілісних властивостей живої організації. Наслідуваність (див. вище) — у першому випадку іманентна властивість матеріальних часток, у другому — епіфеномен внутрішньосистемної регуляції, створеної відбором.
З цієї ж причини одне з ключових понять неодарвінізму — «наследственна змінливість», що позначає варіації, внесені мутаціями, є в рамках ЕТЕ беззмістовним, оскільки для неї наслідуваність — це стійкість, а змінливість — її відсутність. Варіації тут наслідкові лише виключно як ціле, тобто як специфічний спектр змінливості, що характеризує дану норму реакції і тому передається у поколіннях з невизначеною ймовірністю індивідуального успадкування. Але в цьому якості всі варіації (онтогенетичні траєкторії) для ЕТЕ рівнозначні і різняться лише частотою їх здійснення.
Нарешті, з базових постулатів теорії випливають і характерні для неї уявлення про «простір можливостей», мислимий для альтернативних (неприйнятих нею) теоретичних позицій. Так, для ГТЕ принциповий водоразділ проходить між її уявленням, що еволюційні зміни починаються з зародкової клітини, і альтернативною думкою, що вони починаються зі зрілої організації. Друга позиція автоматично ототожнює ГТЕ з ламаркізмом, до різновидів якого, природно, у цьому випадку причисляють і теорію Шмальгаузена. Навпаки, для останньої (ЕТЕ) головний водоразділ лежить між поясненням органічної цілеспрямованості (наследственної стійкості, ср. Шишкін, 2006) як продукту еволюційного процесу і фактичним відмовою від такого підходу. З цієї точки зору ламаркізм і генетична теорія однаково потрапляють для ЕТЕ у другу категорію. Бо якщо в очах ламаркізму наслідуваність «створюється» самою корисністю нового набуття, то в рамках ГТЕ вона подібним же чином тавтологічно пояснюється дією відповідного фактора наслідуваності.
Усі ці властивості теорії демонструють частковий приклад загального правила, що характер і оцінка зчитуваної інформації завжди визначаються особливостями сприйняття реципієнта (у даному випадку специфікою його світогляду). У цьому відношенні наочний приклад дають погляди самого Ч. Дарвіна. Якщо врахувати, що він заслужено вважається зразком наукової безсторонності, то сприйняття ним деяких біологічних узагальнень має для нас цінність своєрідного «контрольного досвіду». Річ йде про оцінку Дарвіном двох явищ, відомих вже за його життя: (1) появу предкових рис при гібридизації і (2) повернення культурних форм до вихідного типу при одичанні в природі.
З точки зору сьогоднішніх уявлень про систему розвитку, причина цих явищ в кінцевому підсумку одна і та сама. Вона пов’язана зі збереженням в епігенетичному просторі досліджуваних форм не повністю стертих предкових траєкторій розвитку. Їх реалізація все ще може бути спровокована — або одразу, завдяки нестабільності розвитку у гибрида (1), або шляхом повторної стабілізації такої траєкторії як результат відновлення відбору на користь колишньої природної норми (2).
Розглянуті з цих позицій, обидва явища однаково добре узгоджуються з дарвінською теорією. Але для самого Дарвіна, при існуючому тоді рівні природничих уявлень, це далеко не здавалося очевидним. Він підтримував еволюційне тлумачення першого феномена, бачачи в ньому прояв рис далекого предка, але виражав стримані сумніви у достовірності другого, тобто прикладів одичання (Дарвін, 2003, с. 31, 32, 160—164). Однією з причин цього скептицизму була те, що такі факти тоді використовувалися проти дарвінської теорії (там же, с. 31) — як свідчення, що процеси змін організмів у природі не описуються механізмом штучного відбору (що саме по собі, безумовно, вірно). Цей приклад ілюструє сказане вище: оцінка фактів дослідником об’єктивно визначається тим, наскільки вони здаються йому сумісними з власною системою поглядів.
II. ВЗАЄМОСПРИЙНЯТТЯ І ЗАМІНА АЛЬТЕРНАТИВНИХ ТЕОРІЙ
У теоріях, побудованих на якісно різних засадах, розуміння причинних зв’язків між явищами в загальному випадку несумісне, внаслідок чого судження однієї з них не можуть бути перенесені на мову іншої. Тому можливість змістовного діалогу між ними зазвичай виключена. У широкому сенсі це стосується не лише наукових узагальнень, а й взагалі будь‑яких різнорідних типів світосприйняття, властивих живим істотам. Носії кожного з таких варіантів здатні відображати явища навколишнього світу лише в рамках доступних їм інформаційних сигналів. Ця ситуація наочно передана в афористичній притчі В. Маяковського (1928) про коня і верблюда, де кожна з цих тварин бачить в іншій лише потворний («неправильний») екземпляр власного виду.
Ця неможливість комунікації між альтернативними системами сприйняття носить об’єктивний характер. Тобто, якщо говорити про теорії, уявлення однієї з них про зміст іншої завжди преломлюються через специфіку її власного бачення. Нерозуміння в цьому випадку може бути і одностороннім, якщо одна з конкуренційних концепцій, будучи узагальненням вищого порядку, здатна показати на своїй мові реальне місце причинних зв’язків, абсолютизованих її опонентом. Але саме одностороння сліпота чинить фатальний вплив на прогрес теоретичного знання, якщо вона властива домінуючій системі поглядів. Приклади такого роду змушують думати, що прогрес теоретичної думки визначається навіть не власне боротьбою ідей. Рішучу роль тут відіграє сама готовність наукової спільноти усвідомити, що домінуючі постулати не є єдиним можливим вибором.
Однією з наочних ілюстрацій сказаного є сприйняття еволюціонізмом концепції Р. Гольдшмідта про систему реагування (розвитку) як об’єкта еволюції. Ця ідея лежить в основі уявлень ЕТЕ. Її майбутнє визнання як ключового положення інтегральної еволюційної теорії здається нам неминучим. По суті Гольдшмідт показав, що аналіз розвитку генетичними методами підтверджує цілісність онтогенетичного процесу, виявлену раніше експериментальною ембріологією. Це тотожність результатів відкриває шлях до інтерпретації генетичних узагальнень у світлі загальних законів розвитку, тобто до розв’язання задачі, яка виглядає найбільш актуальною для еволюційної теорії (ср. Шишкін, 1987, 2006).
Проте зазначена ідея залишилася поза колом інтересів домінуючого напрямку еволюційної думки ХХ століття. Уже кілька поколінь дослідників обговорюють гіпотезу «системних мутацій» Гольдшмідта, з очевидністю не помічаючи причини, що її породила. Між тим вона полягала саме в усвідомленні, що еволюційні зміни не зводяться до ефектів незалежних генів. Але така позиція несумісна з генетичним мисленням і тому просто не сприймається ним. З тієї ж причини сьогоднішні спроби цього мислення знайти місце для цілісних механізмів розвитку в «новому еволюційному синтезі» мало що змінили в його поглядах на еволюцію (см. розділ VI).
Цю ситуацію ілюструє, зокрема, книга Р. Реффа і Т. Кофмена (1986) «Ембріони, гени і еволюція» — одне з перших великих узагальнень у напрямку «нового синтезу». Книга присвячена Р. Гольдшмідту, і автори вважають себе послідовниками його ідеї про еволюцію як системну трансформацію розвитку. Але вони переконані, що Гольдшмідт підрозумів під цим «зміни в генах, що регулюють онтогенез» (Рефф, Кофмен, 1986, с. 13), тобто процеси типу гомеозисних мутацій. Наскільки це далеко від позиції Гольдшмідта, можна зрозуміти хоча б з наступних його слів: «Факти генетики, звичайно, можуть описуватись у термінах генів, але теорія зародженої плазми повинна повністю звільнитися від концепції генів як одиниць» (Goldschmidt, 1938, с. 311). «Зароджена плазма як ціле контролює певну реактивну систему, яка є єдиною системою розвитку» (Goldschmidt, 1940, с. 218). Характерно, що автор цих висновків не жив ілюзіями щодо того, як їх сприймуть: «Для багатьох генетиків явно важко мислити в таких поняттях, оскільки більшість їх настільки пов’язана аксіоматичною вірою в атомістичну генну теорію, що не в змозі думати інакше» (там же).
Перехід від редукціоністського бачення законів еволюції до органістичної (системної) моделі ЕТЕ не був простим навіть для творців останньої. У І.І. Шмальгаузена і К. Вуд‑дінгтона ми знаходимо чимало еклектичних суджень, де змішуються обидва підходи (ср. Шишкін, 1987, с. 99; 2006, с. 184). Там, де ці протиріччя були подолані, це досягалося далеко не одразу. Характерний приклад — розуміння Шмальгаузеном стабілізуючої функції відбору. Запропонована ним концепція з самого початку передбачала принципово нове розуміння наследственної стійкості — як цілісного вираження кореляцій розвитку, створених відбором. Цей підхід логічно виключав розподіл фенотипічних властивостей на продукти мутацій і екзогенних впливів. І, проте, у ранніх роботах з цього питання Шмальгаузен (1940, 1941) описує механізм селективної стабілізації саме в цих редукціоністських термінах — як накопичення «співпадаючих мутацій» під покровом ненаслідувальної «фенокопії», тобто у згоді з гіпотезою співпадаючого (органічного) відбору Моргана‑Болдюїна! Цей приклад показує, що становлення якісно нової світоглядної позиції — процес багатоступеневий, і що ключова ідея тут може з’явитися набагато раніше, ніж її несуперечливе обґрунтування.
Сказане можна підсумувати наступним чином. Стійка система наукових поглядів, як і будь‑яка ефективно регульована система, нечутлива до радикальних нововведень, тобто реагує на викликані ними збурення як на оборотні (релаксуємкі) флуктуації, іншими словами, «не помічає» їх. І лише зниження здатності такої теорії до самопідтримки (у міру вичерпання її пояснювального потенціалу) робить її все більш схильною до змін. Іншими словами, між двома стабільними станами системи в процесі її якісної трансформації завжди лежить нестабільна область фазового переходу, де періоди погашення флуктуацій попереднього стану все довше тривають, аж до того, що стає можливим їх «запам’ятовування». З цим, в кінцевому підсумку, пов’язане відоме спостереження, що зміна теоретичних переконань зазвичай відбувається лише зі зміною поколінь, коли стара парадигма ще не повністю укріпилася у свідомості її нових носіїв.
III. ПРОЦЕС ПІЗНАННЯ
Специфічність відображення реальності будь‑яким реципієнтом означає неповноту цього відображення. Для людського сприйняття ця особливість ускладнюється переважним використанням мовних символів. Простір залежностей у реальному світі безмежний і взаємопов’язаний, тоді як наявні засоби мови обмежені і дискретні. У підсумку будь‑яка наукова теза завжди є продуктом абстрагування від безмежної множини неуловлених причинних зв’язків, що визначили досліджувану подію. Будь‑який висновок лише умовно відображає реальність, згідно з твердженням поета, що «думка виголошена — брехня». Це абстрагування досягає максимуму у висновках, отриманих з безпосереднього спостереження або досвіду.
Об’єктивно мета пізнання — осягнення загальних законів буття; але саме конкретне емпіричне знання є необхідною основою цього процесу в природничих науках. При цьому, здавалося б, незаперечна причинність, безпосередньо виявлена досвідом, з давніх часів фетишизувала цей підхід в очах натурфілософів як власне мету пізнання. Шлях до неї розуміли як розчленування вивченої явиці на складові елементи і пошук (локалізацію) безпосередньої причини для кожного з них (Bertalanffy 1969). Зокрема, з такою детерміністською програми почалася історія експериментальної ембріології, називаної на межі XIX‑XX ст. «механікою розвитку». Цей підхід, як справжній шлях пізнання рушійних сил онтогенезу, протиставлявся натурфілософським поясненням типу геккельського біогенетичного закону (Gould, 1977). У цій же течії проходили пошуки механізмів наслідуваності, починаючи від умозричних редукціоністських концепцій Негелі та Вейсмана і закінчуючи появою хромосомної генетики.
Але, як уже сказано, будь‑які висновки, отримані з досвіду, завжди спрощують реальні співвідношення. Зазвичай це виявляється вже при розширенні або зміні умов експерименту. Початкова пояснювальна схема, в якій для кожної властивості об’єкта передбачалась окрема причина (рис. 5, а), втрачає свою ефективність у міру того, як виявляється, що одна і та сама властивість може мати різні причини і навпаки (рис. 1; 5, б). Така асиметрія видимих причин і наслідків очевидно вказує, що між ними насправді немає лінійних залежностей і що ці дві категорії властивостей пов’язані лише опосередковано, через системні відносини (Waddington, 1957; Белоусов, 1979). Іншими словами, вони належать до двох різних ієрархічних рівнів системи, у яких зміни протікають у різному режимі і прямо не корелюють. Саме такі співвідношення між простором шляхів розвитку і лежачою в його основі структурою геному закладені у уявленні про епігенетичний ландшафт (рис. 2, б).
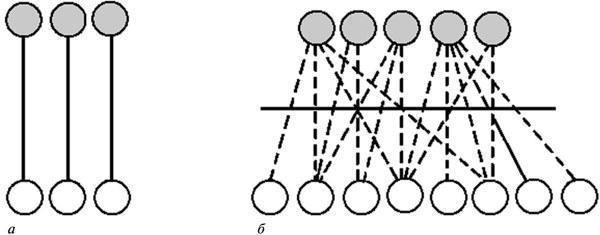
Рис. 5. Етапи оцінки причинно‑наслідкових відносин між двома категоріями факторів, спостережуваних у експерименті: а — початкова гіпотеза, що постулює прості лінійно‑каузальні залежності; б — виявлення асиметрії «причин» і «наслідків» (у міру розширення експериментальних даних), що веде до висновку про їх нелінійні зв’язки та приналежність до двох різних системних рівнів.
У теоретичній біології головним джерелом подібних труднощів для лінійно‑детерміністського мислення завжди був індивідуальний розвиток. Їх наростання у міру накопичення експериментальних даних і пошуку їх несуперечливого узгодження неминуче підштовхувало думку до ідеї про цілісність механізму онтогенезу. В ембріології ця зміна парадигми виявилась у концепції Г. Дрішa (Driesch, 1908) про розвиток як «гармонічну еквіпотенціальну систему», а у фізіологічній генетиці — у згаданій вище ідеї Гольдшмідта про систему реагування. Як ключовий аргумент незмінно підкреслювалося відсутність загального відповідності між конкретним підсумком розвитку і певним варіантом його початкового стану.
Загалом, чим змістовніше досвід науки у осмисленні своїх спостережень, тим більше вона схильна віддалятися від редукціоністських пояснень у бік системних інтерпретацій. І навпаки, кожен прорив науки в колись незнайому область фактичного знання зазвичай супроводжується на першому етапі відродженням механістичних причинних тлумачень. В історії поглядів генетики на детермінацію властивостей організму подібний хід подій повторювався принаймні двічі, характеризуючись спочатку декларацією лінійних причинних залежностей і потім відходом від них (табл. 2). Сучасне чергове відродження надій генетики на можливість редукціоністського тлумачення морфогенезу, пов’язане з відкриттям Nox‑генів, очевидно, матиме ту ж саму долю.
Таблиця 2. «Циклічна еволюція» поглядів генетики на детермінацію розвитку
Область дослідження
Оцінка причинних залежностей
Початкова
Подальша
I. Хромосомна генетика
ознаки визначаються незалежними генами
ознака — продукт взаємодії всіх генів
I. Молекулярна генетика
«один ген — один фермент»
всі етапи матричного синтезу поліваріантні
III. Відкриття гомеобокс‑послідовностей
загальні Nox‑гени керують гомологічними процесами в різних організмах
?У підсумку можна стверджувати, що обмеженість суб’єктивного відображення реальності не дозволяє пізнавати її іншим шляхом, окрім наближення, тобто послідовної ревізії часових пояснювальних моделей. Це просунуте знання не може бути отримане безпосередньо з досвіду.
Мислителі минулого розуміли це давно. За словами Галилея (1948, с. 89), якщо «божественний розум простим сприйняттям сутності… [предмета] охоплює… всю безмежність його властивостей», то «наш спосіб полягає в роздумах і переходах від висновку до висновку».
IV. АХІЛЛЕСОВА П’ЯТА ЕВОЛЮЦІЙНОГО МИСЛЕННЯ
Отже, самоочевидність пояснень, отриманих з прямого спостереження, є ілюзією. Насправді їх характер завжди обумовлений узагальненнями з попереднього досвіду, які, у свою чергу, спираються на певну концептуальну установку. Наприклад, якщо для хромосомної генетики менделєвське розщеплення стало шуканим підтвердженням корпускулярної природи успадкування, то для ЕТЕ (і відчасти класичного менделізму) воно виражає різницю двох цілісних генотипів, виявлювану у схрещуванні як пороговий ефект розвитку (Камшилов, 1935; Шишкін, 1987). У підсумку розуміння діючих причин явища визначається рівнем і способом нашого абстрагування від повноти реальних взаємозв’язків.
Проте в будь‑якому випадку первинне уявлення про причини є необхідним етапом пізнавального процесу. Як уже сказано, ця початкова пояснювальна модель повинна слугувати основою для подальшої корекції та ревізії у міру її випробування (процедури верифікації — фальсифікації за К. Поппером) у світлі нових спостережень. По суті її початкові поняття можуть і після цього зберігати своє значення як робоча мова для опису експерименту. Але як базові положення, що забезпечують неконфліктне тлумачення фактів, вони можуть виявитися вже несуттєвими (Шишкін, 2006, с. 188).
Тут ми переходимо до проблеми, яка здається ключовою для методології сучасного еволюціонізму і, в підсумку, пов’язана з його переважною опорою на генетичне мислення. зіткнення теорії з закономірностями, що не піддаються поясненню в її рамках, є сигналом про те, що рівень спрощення реальності, прийнятий теорією, не відображає повноти нашого фактичного знання. Тобто принципи теорії мають бути так чи інакше переглянуті. Це єдиний можливий шлях теоретичного прогресу.
Між тим оцінка головного напрямку еволюційної думки останнього століття дозволяє говорити про фундаментальне порушення цього правила. Шлях, який був обраний замість нього, фактично можна охарактеризувати як двоїстість колективної наукової свідомості. Вона полягає в тому, що в біології для одного і того ж феномена (а саме розвитку) визнаються два взаємовиключних тлумачення. При цьому використання одного з них генетикою як основи для еволюційної теорії не супроводжувалося жодними спробами довести несуттєвість альтернативного варіанту.
Цю ситуацію можна узагальнити в двох основних пунктах. (1) У редукціоністській ідеології генетики механізм нормального відтворення (успадкування) зводиться до преформативної моделі, де результат — сукупність дії незалежних причин. Навпаки, для ембріології детермінація розвитку має цілісну (системну) природу, що не зводиться до елементарних причин. (2) Власний досвід генетики у вивченні розвитку також давно змушує її до формального відмови від преформативної моделі. Але саме остання саме і лежить в основі ГТЕ (Шишкін 1987, 2006).
Те саме протиріччя виявляється між зазначеною декларативною корекцією поглядів генетики та принципами, що лежать в основі її реальної практики (табл. 3). Нарешті, подібна невідповідність часто має місце і в індивідуальних уявленнях авторитетів ГТЕ, відображаючи приховані розбіжності між власним досвідом дослідника та вимогами зазначеної теорії (табл. 4).
Таблиця 3. Розбіжності між робочими постулатами генетики (лівий стовпець) і положеннями, що визнаються нею декларативно або за замовчуванням. Перші засновані на абсолютизації типових експериментальних залежностей, другі — на узагальненнях, що мають ширшу основу
Висновки, екстраполовані з гібридного аналізу чистих ліній
Поділені генетикою положення, засновані на ширшому (теоретичному та емпіричному) знанні
Признаки діляться на генетично обумовлені (успадковані) і екзогенні (неуспадковані)
Всі ознаки — це вирази генотипічно обумовленої норми реакції
Признаки детермінуються незалежними генами
Кожен ознак визначається всім генотипом
Успадкування ознак підпорядковується менделєвським законам
Менделєвське успадкування не характерне для «сирової» природної змінливості
Таблиця 4. Приклади невідповідностей між власними узагальненнями дослідника та підтримуваною ним теорією (ГТЕ)
Е. Майр (1968)
Генетична теорія еволюції
Еволюція починається з поведінки
Еволюція починається з змін у генах
Усі мутації з фенотипічним проявом шкідливі
Мутації з корисним проявом зберігаються відбором
Мутації самі по собі не змінюють видову приналежність особини
Мутації лежать в основі видових змінТака двоїстість у оцінці механізмів розвитку та еволюції виявляє головну проблему генетичного мислення, пов’язану з його редукціоністським розумінням феномену спадковості. Як уже не раз зазначалося ембріологами, воно фактично передбачає, що спадковість є чимось окремим від онтогенетичного здійснення і тому може бути пояснена незалежно від нього, тобто поза межами реального знання закономірностей розвитку (Gurwitsch, 1912; Гурвич, 1944; Светлов, 1964). Але ці застереження не вплинули на домінуючу еволюційну доктрину (ГТЕ), окрім визнання, що вона допускає значні спрощення. Сучасні спроби узгодити генетичне розуміння спадкової детермінації з принципом морфогенетичних полів, що прийшов з ембріології, залишаються в межах тих самих початкових преформаційних уявлень (Шишкін, 2006, с. 193). Точно так само і спроби досягти “нового еволюційного синтезу” шляхом об’єднання ГТЕ з закономірностями ембріогенезу віднюдь не ставлять собі за задачу перегляд постулатів неодарвінізму (см. розділ VI). Початковий протиріччя залишається нерозв’язаним.
У підсумку важливо підкреслити наступне. Поширення причинного пояснення за межі тих умов досвіду, в яких воно було отримано, саме по собі є неминучим кроком пізнавального процесу. Але з того моменту, коли виявилася неадекватність цього пояснення для нових або розширених умов, його слід змінити. Ідея мозаїчної детермінації ознак, що становить основу генетики, на практиці уникла такого перегляду, незважаючи на її очевидну непридатність для пояснення розвитку (ср. Светлов, 1978). Це початкове порушення логіки пізнавального процесу лежить в основі всіх з часом виявлених головних труднощів генетичної теорії еволюції.
Причини такого ходу подій, тобто затягнутого неявного співіснування двох несумісних концепцій розвитку, значною мірою лежать на поверхні. Реальна відмова генетики від принципу мозаїчної детермінації зробила б за означенням неможливим опис еволюції в термінах генів, що становить основу ГТЕ. Це означало б надто радикальний поворот у домінуючому еволюційному мисленні, щоб можна було розраховувати на його історично швидке здійснення.
Інші причини мають у природничих науках ще більш давні корені. Це, передусім, традиція редукціоністського розуміння причинності, що віддає перевагу механістичним поясненням і каналізує пошуки рішень у цьому напрямку (см. вище, розділ III). І, нарешті, треба вказати на невисокий рівень вимог теоретичної біології до узгодження своїх узагальнень. Останнє історично пов’язане з різноманіттям органічного світу і складністю виявлення в ньому універсальних закономірностей. Тому, зокрема, типово для еволюціонізму є не прагнення до створення цілісної теорії, а “синтез” тих чи інших факторів, розумітих як незалежні учасники еволюційного процесу (Шишкін, 2006, 2009).
V. ІЛЛЮЗІЇ ЕВОЛЮЦІЙНИХ РЕКОНСТРУКЦІЙ
Абсолютизація залежностей, отриманих безпосередньо з експерименту або порівняння, без оцінки їх реальної значущості в світлі інших біологічних узагальнень, становить типову практику тлумачення еволюційних процесів на основі дослідного знання. Такий підхід характерний як для ембріології, так і для генетики (Шишкін, 2006; 2007, с. 180).
Предметом вивчення для обох названих дисциплін є простір варіантів розвитку та контрольні за ними механізми. І обидві вони найчастіше (у випадку генетики — завжди) судять про еволюційне значення спостережуваних явищ, спираючись за замовчуванням на дві презумпції. (1) Історичні зміни протікають у тому ж напрямку, що й онтогенетичні (від ранніх стадій розвитку до пізніх); тому другі можуть бути прямим відображенням перших. (2) Зв’язки між дорослими структурами і їх причинами у ранньому розвитку або зародковій клітині можуть бути спрощені до лінійних відносин, тобто до преформаційної моделі.
Як уже сказано, друге з цих припущень у теорії відкидається обома дисциплінами, тобто опора на нього завідомо нелогічна. Що стосується припущення (1), то в ембріології воно не має спеціального обґрунтування (крім лише розширювального тлумачення біогенетичного закону) і приймається скоріше емпірично — як найпростіше з можливих (Шишкін, 2007). Навпаки, для генетики це припущення прямо випливає з її ідеології, що приймає, що властивості організму детермінуються генами, а зміни цих властивостей — мутаціями генів. Це обставина, разом з фактом, що саме генетика є основою домінуючих еволюційних уявлень, визначає нашу головну увагу до практики історичних реконструкцій на основі генетичних даних.
Як уже говорилося (см. Вступ), генетичному розумінню еволюції протистоїть позиція ЕТЕ, згідно з якою еволюційні зміни поширюються у поколіннях від дорослої стадії до початку розвитку. Іншими словами, згідно з даною теорією, зміни проявляються раніше, ніж формується організація зародкової клітини, що забезпечує їх стабільне відтворення (рис. 4, б: F1—F4). Якщо ГТЕ прямо проєктує на еволюцію результати експериментів, що визначають у її розумінні зв’язки генів і ознак (у класичному випадку — на основі гібридного аналізу), то позиція ЕТЕ базується на осмисленні загального різноманіття емпіричних залежностей, спостережуваних в онтогенезі і не вкладаються як цілість у лінійні схеми. Відмінності цих двох підходів грають вирішальну роль в оцінці можливостей еволюційних реконструкцій.
Вище підкреслювалося, що при порівнянні ходу розвитку в різних умовах експерименту виявляється загальна асиметрія причин і наслідків (рис. 1; 2, б; 5, б). І що особливо важливо, виявляється обмеженість простору других щодо різноманіття перших. Головний висновок з цього феномена (особливо у застосуванні до нормального фенотипу), відомий ще з кінця XIX ст., зводиться до правила: “результат розвитку більш стійкий, ніж спосіб його здійснення”. Цей висновок закладений в узагальненнях, відомих в ембріології як “правило Ру”, принцип еквіфінальності Дріша, принцип нормування Гурвіча, а також у багатьох узагальненнях, що позначають еволюційне стискання і спрощення розвитку при збереженні його попереднього результату, виявлених порівняльним шляхом (Шишкін, 2006, с. 182). У генетиці ця ж закономірність виражається в правилі Четверикова (генетична гетерогенність під покровом “дикого” фенотипу), у уявленнях Гольдшмідта про систему розвитку, у факті гетерогенності однотипних природних аберацій і в безлічі прикладів реалізації однакових властивостей на різній генетичній основі (см., наприклад, Шмальгаузен, 1982; Дубінін, 1966). Одним із вражаючих прикладів цього правила є детермінування статі, реалізоване в органічному світі на основі найрізноманітніших механізмів — від чоловічої або жіночої гетерогаметності до визначення факторами середовища, причому іноді різними способами у предельно близьких таксонах.
У результаті, навіть безвідносно до постулатів ЕТЕ, весь досвід експериментального знання про розвиток, врахований цією теорією, може бути узагальнений у вигляді одного базового принципу, що проливає світло на еволюційну оцінку подій онтогенезу. А саме, що програма розвитку, закладена в зародковій клітині, завжди історично молодша, ніж дефінітивний результат, реалізований цією програмою. Остання характеризує лише один із варіантів такої реалізації, поряд із багатьма іншими — як часто існуючими паралельно, так і необхідно передувшими даному варіанту в процесі його становлення. Іншими словами, історія кожної нової елементарної особливості починається з її появи на основі недосконалого морфогенезу, який далі перетворюється все глибше. (У уявленнях ЕТЕ цей процес описується як стабілізація нововведення; см. рис. 4, б: F1—F4). Повторимо, що такий погляд на еволюцію ходу розвитку є не лише одним із ключових положень ЕТЕ, а й висновком з загальної оцінки властивостей онтогенезу як системи. За висновком ембріолога Л.Б. Белоусова (1979) еволюція — це посилення і поширення на нижчі системні рівні тих змін (“нестабільностей”) онтогенезу, які були спочатку передбачені на верхньому (морфологічному) рівні.
З сказаного зрозуміло, що шукати у функціональних властивостях геному або зародкової клітини прямі вказівки на спосіб історичного виникнення реалізованих ними дорослих особливостей — надто наївне очікування. На практиці все, що ми спостерігаємо в сучасному онтогенезі, що створює даний дорослий план будови, є максимально трансформованою (до зиготи) моделлю його розвитку, що пройшла довгий шлях стабілізації. Вона далека від початкової моделі (ср. рис. 4, б: F1, F4), яка була відносно нестабільною, слабо інтегрованою (без багатьох кореляцій, виявлених у сучасному онтогенезі) і залежною від зовнішніх факторів. Тому особливості розвитку будь‑якої структури, зареєстровані експериментатором, є продуктом усієї історії її існування, а не вказівкою на механізм її появи в еволюції. Дозволяти протилежне означає приймати наслідки за причини. Насправді, словами С.Г. Крижановського (1939), розвиток безпосередньо не містить у собі методу для своєї історичної інтерпретації.
Між тим, практично всі судження про природу еволюційних нововведень, що випливають з даних генетики, основані саме на зазначеному ототожненні причин і наслідків історичного процесу. Такі, перш за все, безліч гіпотез про походження різних фенотипічних властивостей за рахунок ефектів мутацій, розумітих як одночасні стійкі зміни фенотипу (наприклад, у випадку меланізму або альбінізму ссавців, редукції крил у комах, торсіонного процесу у брюхоногих молюсків тощо). Ці гіпотези, не передбачаючи жодної історичної зміни початкових факторів розвитку щодо його конкретного результату (ср. рис. 4, а), неминуче передбачають і інші несуттєві припущення. По‑перше, вони спираються на преформаційну модель розвитку, переносячи її механічно далі на еволюцію. По‑друге (у відповідності до першого припущення), вони виходять з переконання, що “генетична основа” ознаки, виявлена гібридним аналізом, є її історичним “детермінуючим фактором”. Насправді ж мова йде лише про порогову функцію цього фактора (при переключенні батьківських онтогенетичних траєкторій у гибридів), що виникає виключно в даній комбінації схрещування (детальніше см. Шишкін, 1987, с. 95‑98, 101).
Те саме розуміння еволюційного процесу закладено і в більш складних реконструкціях, наприклад, при освітленні еволюції комах від предків, близьких до багатоножок, у напрямку двокрилих (Lewis, 1985; Рефф, Кофмен, 1986). Ця трансформація описується як результат послідовних мутацій (або дуплікацій з подальшою діверсією функцій генів), що спочатку призвели до втрати кінцівок у абдомінальних сегментах у ранніх криластих комах, а пізніше (у Diptera) також до редукції крил задньої грудей. Уся схема побудована на аналізі гомеозисних переключень в уявних дисках у вищих комах при мутаціях або послідовних делеціях у відповідних генних комплексах. Тобто мова йде про особливості історично глибоко трансформованого типу морфогенезу. Далі їх екстраполюють на ранні етапи еволюції комах, на яких не було ні розвитку з повним перетворенням, ні властивого йому передбачуваного відокремлення зачатків дорослих тканин (уявних дисків), ні, відповідно, високо інтегрованих формативних залежностей, виявлених як альтернативні програми розвитку цих зачатків. Таким чином, формоутворювальні гомеозисні реакції, що характеризують вищих комах, у даній реконструкції подій (а) приписуються специфічним генам, (б) переносяться на примітивні типи розвитку, що не мали таких властивостей, і (в) ототожнюються безпосередньо з еволюційними механізмами. Це ще один приклад змішування причин і наслідків еволюційного процесу, заснований на уявленні, що причинні зв’язки ходу розвитку, спостережувані в експерименті, і еволюція — це в принципі одне і те ж (ср. рис. 4, а: A—D).
У підсумку вся логіка умозаключень у цих реконструкціях об’єктивно випливає з переконання, що мозаїчна детермінованість розвитку є єдино можливим висновком з даних генетики — незалежно від того, наскільки він сумісний з узагальненнями ембріології.
VI. СИМПТОМИ КРИЗИСУ ТЕОРІЇ
Повернемося до питання про фактори, що визначають втрату теорією статусу достовірного пояснення реальності. Використовуючи знову еволюційне вчення як модельний об’єкт, ми приходимо до частково неочікуваного висновку, що для такого ходу подій взагалі не потрібна поява протиставної альтернативної концепції, принаймні в оформленому вигляді.
Будь‑яка модель функціонування, притаманна живим організмам, включаючи і людську діяльність, передбачає певну систему відображення реальності, з якою узгоджуються всі виконувані дії. Наукова теорія — така ж система, що організує пізнавальний процес, незалежно від того, чи усвідомлює це її носій. Зазвичай вона включає невелику кількість вихідних припущень. Цінність її для пізнання тим вище, чим ширший круг явищ вона дозволяє пояснити на цій основі, показуючи їх загальні причинні зв’язки. Таким чином, змістова теорія максимально економна (рис. 6, б). Вона не замінює дослідного знання, але сильно полегшує вибір можливих для нього напрямків пошуку, відсікаючи шляхи, завідомо безперспективні в світлі її уявлень. У цьому суть відомого узагальнення, що немає практичнішої речі, ніж хороша теорія.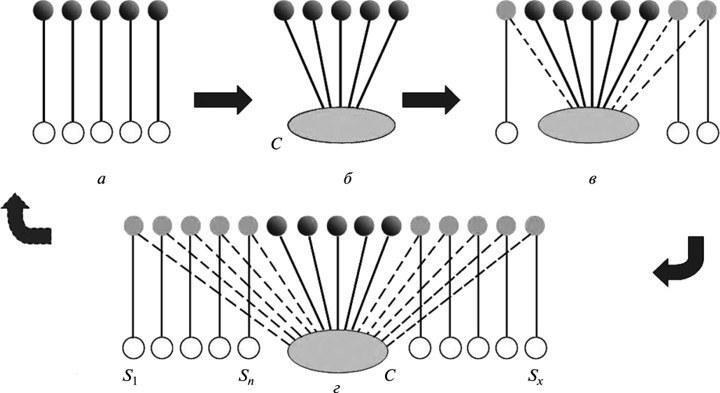
Рис. 6. Симптоми кризи теорії: а — вихідний етап пізнання (відсутність загального пояснення для спостережуваних явищ); б — появлення теорії (перехід до тлумачення фактів на єдиній основі); в — виявлення фактів, не пояснених теорією без додаткових часткових припущень; г — наростання ситуації, що виникла на етапі (в). Пояснювальні можливості теорії скорочуються, наближаючи її до стану, що існував на етапі (а). Позначення: С — концептуальна основа теорії; S1, Sn, Sx, — додаткові припущення.
Прогрес у сфері знань, до якої належить теорія, може бути стимулом до її удосконалення і конкретизації її понять. Наприклад, для неодарвінізму таким кроком було відкриття хромосом і їх комбінування разом з мутаційним процесом, а для класичного дарвінізму (з великим запізненням) — відкриття системних властивостей онтогенезу. Ці зміни можуть сильно змінювати мову теорії, але не зачіпають її постулатів і не порушують її пояснювальної логіки.
Положення стає іншим, коли теорія стикається з фактами і узагальненнями, що суперечать її очікуванням. Усвідомлення нею таких перешкод не означає, що вони не були відомі науці задовго до цього. Але спосіб їх сприйняття, якщо говорити про біологію, змінюється з часом і залежить від багатьох причин. По‑перше, потреба в узгодженні узагальнень, отриманих у різних галузях біологічного знання, мало властива колективному науковому свідомості (Шишкін, 2006; ср. вище, розділ IV) — доки вона не стає самоочевидною. По‑друге, у своїй оцінці “незручних” фактів теоретичне мислення схильне керуватись не стільки загальними вимогами логіки, скільки інерцією свого концептуального вибору, що передбачає правоту і незмінність захищуваної теорії. Очевидно, з цих причин радикальне протиріччя між принципом мозаїчної детермінації і свідченнями цілісності розвитку залишалося протягом багатьох років поза полем зору неодарвінізму (ГТЕ) у після‑Вейсманівську епоху. В іншому випадку це вчення ніколи не змогло б стати домінуючою доктриною в біології ХХ ст. Історики науки називають і більш конкретну причину такого ходу подій (по суті включаючу обидві попередні). Вони вказують, що ідея морфогенетичного поля, що визначає хід розвитку, відкидалась моргановською генетикою як небезпечна альтернатива уявленню про ген як фактор детермінації (Гілберт та ін., 1997, с. 327—330).
Оцінка адекватності ГТЕ стала змінюватись у колективному свідомості лише по мірі того, як детальний аналіз механізмів розвитку (як щодо продуктів синтезу, так і морфологічних особливостей) став однією з головних задач генетики. Як і слід було очікувати, він приніс нові підтвердження відомого правила, що співвідношення між причинами і їх результатами в розвитку неоднозначні на всіх його рівнях, починаючи з експресії генів (ср. табл. 2). Зрозуміло, що для теорії, що зводить пояснення еволюції до зміни генетичного складу популяцій, це означає нерозв’язні проблеми. Зазначена невизначеність стала стимулом до пошуків “нового синтезу”, у якому знайшло б місце для причинних співвідношень і механізмів, виявлених ембріологією.
Головна труднощі окреслювалась ініціаторами цієї нової програми наступним чином. Щоб зрозуміти зв’язок між геномними змінами і еволюцією, необхідно знати, як структура геному перетворюється в певну морфологію. Але тут між сигналом і відповіддю існує “чорний ящик”. Процеси, ізоморфні геному, закінчуються на визначенні первинної структури білків і знаходяться далеко від процесів розвитку, що формують організм (Evolution..., 1982). Отже, пояснення останніх треба шукати іншими шляхами, тобто звернувшись до ембріологічного розуміння закономірностей морфогенезу.
Цей план реорганізації теорії фактично покладає надії на її удосконалення за рахунок альтернативного їй принципу (системне пояснення розвитку), без перегляду її попередньої основи. Справжній новий синтез зазвичай уявляється як “поєднання” цих різнорідних передумов. Приймається, що сьогоднішня ГТЕ все ще адекватна для тлумачення мікро‑ево‑люційних процесів, але що зміни макро‑еволюційного рівня мають описуватись на основі знань законів онтогенезу (Gould, 1982; Maderson et al., 1982).Такий же підхід закладений і в спробах побудови концепції розвитку на основі синтезу уявлень генетики та ембріології — як ряду етапів каскадної експресії генів у ході поступової детермінації. Він зводиться до ідеї чергування активності генів і морфогенетичних полів, контрольованих генами, причому вся послідовність подій має ініціюватися генами вищого ієрархічного порядку (Gilbert та ін., 1997). По суті це лише сильно ускладнений варіант преформативної моделі розвитку, у якій дія системних факторів грає суто підпорядковану роль (Shishkin, 2006). Підсумовуючи, мова йде про спроби відновити пояснювальну цінність генетичної теорії шляхом включення в неї чужорідних принципів, сфера дії яких обмежується довільним чином.
Труднощі генетичного пояснення еволюції не лише прямо сфокусовані в його ключовому постулаті (відбір мозаїчних детермінантів розвитку), а й мають безліч інших пов’язаних з ним проявів. Багато з них досі не усвоєні колективною свідомістю, хоча, здавалося б, лежать на поверхні. Так, не усвідомлюється, що генетична (“еволюційно значуща”) обумовленість ознак, приписувана неодарвінізмом дії мутацій, є насправді властивістю будь‑якої реалізованої особливості, оскільки всі вони — вирази норми реакції (табл. 3; ср. Shishkin, 2006, с. 183, 188). Або що ефект менделївської успадкованості, спостережуваний у гібридному аналізі, створюється власними зусиллями дослідника, що спочатку стабілізує чисті батьківські лінії. І тому бачити в цьому феномені фактор еволюції, що керує природною змінливістю, означає приймати продукт відбору за його сирий матеріал (Shishkin, 1987, с. 117).
Там, де такі труднощі очевидні для теорії, вона намагається пояснити їх за допомогою додаткових допущень ad hoc (ср. табл. 1). Але ці спроби лише позначають проблему, не дозволяючи її усунути. Якщо, наприклад, ефекти мутацій завжди успадковуються гірше, ніж норма, то, яким би шляхом це не пояснювали, вони завідомо не володіють тією історичною стійкістю, яку їй приписує неодарвінізм. Або, якщо співвідношення геному і фенотипу принципово неоднозначні, то незрозуміло, як у цьому випадку можна пояснити механізм відбору мутацій, на якому базується зазначена теорія.
Схематично весь розглянутий хід подій в історії існування теорії описується наступним чином. Чим більше зростає область фактів, важко сумісних з нею, тим ширше вона змушена вдаватися до пояснень, що вимагають часткових додаткових гіпотез (ср. табл. 1). У результаті “пояснювальна структура” теорії все більше наближається до того стану, який передував її появі (рис. 6, а). Іншими словами, ранній етап, на якому пропонувалася загальна принципова відповідь на безліч різнорідних питань (рис. 6, б), змінюється етапами, на яких зростає симетрія між кількістю питань, що виникають перед теорією, і кількістю пропонованих відповідей (рис. 6, в, г). У підсумку ця пояснювальна схема як ціле перестає бути економною і прогностичною, тобто все більше втрачає статус теорії. Намагаючись захиститися, теорія руйнує саму себе.
Якщо в еволюції теорії позначився такий хід подій, то його фінал передбачуваний незалежно від того, чи існує для даної теорії складена альтернатива. Остання все одно має рано чи пізно виникнути в пошуках відповіді на ті питання, для яких не знайшлося пояснення в рамках існуючих уявлень. У випадку розвитку ідеї еволюції на основі природного відбору так воно фактично і відбувається. Хоча системна альтернатива неодарвінізму у вигляді епігенетичної теорії Шмальгаузена‑Уоддінгтона (ЕТЕ) існує вже понад півстоліття, її вплив на колективну еволюційну думку був досить обмеженим через багато причин (ср. Shishkin, 2006, с. 184). Проте незадоволеність прихильників ГТЕ її нинішнім станом стимулює пошуки нового, більш широкого “синтетичного” узагальнення на тих самих шляхах, на яких склалася ЕТЕ. Незважаючи на опортуністичний характер більшості таких спроб, що прагнуть зберегти в новій теорії центральну доктрину неодарвінізму, саме їх напрямок показовий. Він говорить на користь кінцевої неминучості визнання принципів, що лежать в основі ЕТЕ.
**ВИСНОВКИ (ПЕРСПЕКТИВИ ЕВОЛЮЦІОНІЗМУ)**
Зі сказаного зрозумілий найймовірніший прогноз щодо подальшого розвитку еволюційної теорії. Він базується на переконанні, що при всій своїй традиційній мозаїчності теоретичне мислення в біології все ж є єдиною системою з внутрішніми зворотними зв’язками. Тому воно в кінцевому підсумку здатне до поступової самокорекції, заснованої на критичному порівнянні та узгодженні власних узагальнень. Цей процес неминуче має привести колективну еволюційну думку до усвідомлення несумісності системних властивостей розвитку з принципом корпускулярної успадкованості. Відповідно до цього завтрашня еволюційна теорія розглядатиме зміну органічних форм як перетворення цілісних систем розвитку. Усі властивості розвитку, розкриті ембріологією та генетикою, будуть інтерпретовані в цьому контексті. Спроби фетишизації будь‑яких структур живого як самостійних факторів еволюції, що виникають і діють поза природним відбором, залишаться для теорії в минулому.
**СПИСОК ЛІТЕРАТУРИ**
Белоусов Л.В. Цілісні та структурно‑динамічні підходи до онтогенезу // Журн. общ. біол. 1979. Т. 40. № 4. С. 514‑529.
Галілей Г. Діалог про дві головні системи світу — Птолемеївську і Коперникову. М‑Л.: ОГІЗ — Держ. вид-во Техн.-теор. літ., 1948. 379 с.
Gilbert C.F., Opitz D.M., Ref R.A. New synthesis of evolutionary biology and developmental biology // Ontogeny. 1997. Т. 28. № 5. С. 325—343.
Гурвіч А.Г. Теорія біологічного поля. М.: Сов. наука, 1944. 156 с.
Дарвін Ч. Походження видів шляхом природного відбору. М.: Тайдекс Ко, 2003. 495 с.
Дубінін Н.П. Еволюція популяцій і радіація. М.: Атоміздат, 1966. 743 с.
Камшилов М.М. Чи є плейотропія властивістю гена? // Біол. журн. 1935. Т. 4. № 1. С. 113—144. Крижановський С.Г. Принцип рекапітуляції і умови історичного розуміння розвитку // Пам’яті акад. А.Н. Севецького. М.; Л.: Вид-во АН сСР, 1939. С. 281—366.
Любищев А.А. Про природу спадкових факторів // Ізв. Біол. наук.-досл. ін-ту при Пермському університеті. 1925. Т. 4. Дод. 1. С. 1—142.
Маяковський В. Вірші про різницю смаків // Збірн. соч. у 3 томах. Т. 2 (1928). М.: Худ. літ-ра, 1978. С. 358.
Mair E. Zoological species and evolution. М.: Мир, 1968. 597 с.
Reff R., Kofmen T. Embryos, genes and evolution. М.: Мир, 1986. 402 с.
Svetlov P.G. Про цілісний і елементаристичний методи в ембріології // Арх. анатомії, гістології та ембріології. 1964. Т. 46. № 4. С. 3—26.
Svetlov P.G. Фізіологія (механіка) розвитку. Л.: Наука, 1978. Т. 1. 279 с.; Т. 2. 262 с.
Filippenko Yu.A. Еволюційна ідея в біології. М.: Наука, 1977. 227 с.
Shishkin M.A. Онтогенез і еволюційна теорія // Еволюція і біоценотичні кризи / Ред. Л.П. Тат-аринов, А.П. Расницин. М.: Наука, 1987. С. 76—123.
Shishkin M.A. Закономірності еволюції онтогенезу // Сучасна палеонтологія / Ред. В.В. Меннер, В.П. Макридин. Т. 2. М.: Недра, 1988. С. 169‑209.
Shishkin M.A. Індивідуальний розвиток і уроки еволюціонізму // Онтогенез. 2006. Т. 37. № 3. 179‑198.
Shishkin M.A. Морфогенез як об’єкт історичного прочитання: продукт еволюції чи її запис? // Клітинні, молекулярні та еволюційні аспекти морфогенезу. Симпозіум з міжнародною участю / Ред. Л.В. Белоусов. М.: КМК, 2007. С. 179‑183.
Shishkin M.A. Еволюційна теорія та особливості наукового мислення // Наукова спадщина Шмальгаузена і його розвиток. Конф. присв. 125‑річчю зі дня народження акад. І.І. Шмальгаузена. М.: ІПЕЕ РАН ім. А.Н. Севецького, 2009. С. 46‑50.
Schmalhausen I.I. Шляхи і закономірності еволюційного процесу. М‑Л.: Вид-во АН СССР, 1940. 231 с.
Schmalhausen I.I. Стабілізуючий відбір і його місце серед факторів еволюції // Журн. общ. біол. 1941. Т. 2. № 3. С. 307‑354.
Schmalhausen I.I. Організм як ціле в індивідуальному та історичному розвитку. М.: Наука, 1982. 228 с.
Bertalanffy L. General system theory. N.Y.: Braziller, 1969. 289 p.
Driesch H. The science and philosophy of the organism. L.: Black, 1908. V. 1. 329 p.; V. 2. 381 p.
Evolution and development // Report of the Dahlem Workshop / Ed. J.T Bonner. Berlin, Heidelberg, N.Y.: Springer, 1982. 357 p.
Goldschmidt R. Physiological genetics. N.Y.: L. McGraw Hill Book Co., 1938. 375 p.
Goldschmidt R. The material basis of evolution. New Haven: Yale Univ. Press, 1940. 436 p.
Gould S.J. Ontogeny and phylogeny. Cambridge: Belknap Press, 1977. 501 p.
Gould S.J. The meaning of punctuated equilibrium and its role in validating a hierarchical approach to macroevolution // Perspectives in evolution / Ed. R. Milkman. Sunderland, MA: Sinauer, 1982. P. 83‑104.
Gurwitsch A. Die Vererbung als Wirklichkeitsvorgang // Biol. Zbl. 1912. Bd 32. S. 458‑486.
Lewis E.B. Regulation of the genes of the bithorax complex in Drosophila // Cold Spring Harbor Symp. Quant. Biol. 1985. У 50. P. 155‑164.
Maderson P.F.A., Alberch P., Goodwin B.C. et al. The role of development in macroevolutionary change // Evolution and development. Dahlem Konferenzen / Ed. J.T. Bonner. Berlin: Springer, 1982. P. 279‑312.
Waddington C.N. The strategy of the genes: a discussion on some aspects of theoretical biology. L.: Allen and Unwin, 1957. 262 p.
Evolutionary Theory and Scientific Thinking M. A. Shishkin
In natural sciences, the advance of evolutionary thought and growth of empirical knowledge are not strictly correlated. The state of theory primarily tends to be controlled by a mode of collective thinking that historically dominates a given branch of science. This particularly holds true for the natural selection concept, which has two alternative interpretations known as the genetic and epigenetic theories of evolution. The final result of their competition, albeit predictable, will not be based upon any kind of “crucial evidence” giving advantage to either of them. The above result will be in fact attained as soon as the evolutionary biology can overcome the tradition of mosaic thinking which enables the incompatible concepts to be combined. In this respect, the key point to be realized is that the idea of corpuscular determination of the ontogeny is incompatible with understanding the development as a systemically controlled process.
Key words: evolution, genetics, epigenetic theory, ontogeny.
1Такий же шлях еволюції постулюється для провізорних адаптацій, реалізованих у ході розвитку (Shishkin, 1988, с. 183‑186).