Creation of Worlds — 11. Ідеї для моделювання
Модели, которые со временем стоило бы сделать...
"Біосфера", що складається з "екосистем", що можуть перебувати у стійких або нестійких станах. За яких умов така біосфера буде еволюціонувати до більшої стійкості без дарвіновського добору та репродукції окремих одиниць?
Проблема має сенс, наприклад, з точки зору дискусії Дж. Лавлока та Р. Докінза. Концепція Геї Лавлока передбачає доцільні механізми планетарної регуляції. Доцільність — результат добору. Докінз стверджує, що добір завжди пов'язаний з діференціальним відтворенням реплікаторів. Якщо б існувало багато біосфер, деякі гинули, а інші — розмножувалися, тоді б, за Докінзом, біосфери могли розвинути здатність реагувати доцільно. Згідно з припущенням, яке можна перевіряти за допомогою моделювання, доцільність може бути наслідком і іншого типу добору, який можна назвати добором за Больцманом — поширенням більш термодінамчіно ймовірних (більш стійких) станів (деякі пояснення — тут). Якщо це припущеня є вірним, підсистеми такої системи як біосфера можуть знаходитися у станах різної стійкості. Чи може добір (перехід у більш термодінамічно ймовірні стани) таких підсистем бути механізмом розвитку доцільності у системи, що не залучена до процесу диференціальної реплікації?
Аналогія. Чи може добір організмого рівня бути причиною доцільності популяційних механізмів регуляції без дарвіновського добору (диференціальної загибелі та диференціальної реплікації, тобто розселення) популяцій?
Чи буде зростати гіперболічно (як чисельність людства за фон Ферстером) чисельність популяції, що існує не необмеженому двомірному просторі та здатна з певною швидкістю розселюватися у незайнятих напрямах? Точніше, за якою моделлю вона зростатиме (відносне збільшення площі ареалу буде зменшуватися).
Конкуренція за спільний ресурс двох популяцій. Чисельність однієї контролюється хижаками, чисельність іншої — нестачею ресурсу. За яких умов хижаки забезпечать "перемогу" своїх жертв над їх конкурентами?
Вплив кризи перенаселення на ємність середовища. Слони, що живляться акаціями у саванні. Коли їжі достатньо — поїдають листя, коли не вистачає — ламають стебла та обдирають камбій. Розширення моделі — вплив на цей процес гомотеріїв та смилодонів.
Істоти Деннета! Детальніше тут і тут. Скінерова істота, що буде радіти виграшам та сумувати у разі програшів.
Перехід популяції птахів, що живуть на витягнутому вздовж меридіана континенті, від осілості до перельотів. У високих широтах кращі умови для розмноження, у низьких — вищі шанси пережити зиму.
Регуляція річного циклу у популяції, яка живе у помірному кліматі: на що краще реагувати, на динаміку температури чи довжину світлового дня? Порівняти різні способи регуляції за кількістю "помилкових" днів за різних рівнів мінливості клімату.
Жіночі та чоловічі репродуктивні стратегії людини.
Різні горизонти планування за різного рівня мінливості середовища.
«Небагато хто для вічності живе,
Та якщо ти миттєвим заклопотаний —
Твій жереб страшний і твій дім неміцний!
(О. Мандельштам)
Конкуренція двох популяцій із статевим розмноженням та двома статями в умовах стабілізуючого відбору. В одній мінливість і смертність (що прибирає особин, що відхиляються від норми) однакові для самців і самок, в іншій — самці мінливіше і, отже, сильніше страждають від негативного відбору. Ідея В. Геодакяна.
Порівняння швидкості вироблення адаптацій у разі клонального та статевого відтворення.
Порівняння поширення нових ознак при генетичному та культурному наслідуванні.
Фазовий перехід в еволюції (від генів до мемів, від мемів до т-мемів і далі).
Повільні та швидкі інфекції за умов різної популяційної структури.
Принцип Фішера при різній "ціні" особин різних статей або у разі зміни співвідношення статей під час онтогенезу.
Хроповик Меллера.
"Гіпотеза бабусі" та походження клімаксу у людини.
"Стокротковий світ" Лавлока.
ІмІтаційне доведення парадоксу Монти Голла. Модель, яка демонструватиме успішність двох різних стратегій (міняти вибір чи не змінювати) можна зробити за аналогією з моделлю про нарожденння хлопчиків та дівчинок (що вирішує одну из відомих демографічних задач). Якось Великий Султан зіштовхнувся з складною проблемою. Представники збіднілої знаті стали скаржитися, що їм складно набирати собі гареми, що відповідають їхньому статусу. Дівчаток народжується приблизно стільки ж, скільки і хлопчиків, і після того, як багатії заберуть до себе в гареми по сотні дружин, їм, власне, нікого й не лишається. Щоб вирішити проблему, Султан вирішив збільшити кількість народжуваних дівчаток по відношенню до кількості хлопчиків. Для цього він видав такий указ: «Жінці дозволяється народжувати дітей доти, доки вона не народить першого хлопчика. Після цього їй народжувати забороняється. Таким чином, у кожній родині може бути не більше одного хлопчику, а дівчаток скільки завгодно». Наскільки зміниться відношення кількості дівчаток до кількості хлопчиків після виконання цього указу?
Ймовірно, у такій моделі треба буде згенерувати безліч випробувань з випадковим розміщенням кози та автомобілів, відкриванням одного з дверей з козою та підрахунком виграшів двох гравців, один з яких змінює двері, а другий – ні. Збирати статистику результатів цих спроб слід за допомогою лічильника ітерацій, аналогічному такому в моделі про хлопчиків або моделі "Гаузе".
Динаміка концентрації кисню під час кисневої революції. Треба врахувати:
— запас відновників, який поступово витрачається;
— приплив відновників з надр, що поступово слабшає;
— відносно постійну втрату атмосфери;
— зниження ефективності примітивних фотосинтетиків у міру виникнення локальних острівців кисню;
— появу і поширення більш досконалих фотосинтетиків, що ефективно працюють в окисному середовищі.
А потім можна спробувати підібрати параметри так, щоб динаміка концентрації кисню приблизно відповідала тому, що є відомим з геологічного літопису.
Перехід від гермафродитизму з перехресним заплідненням до роздільної порожнини. Модель та її опис є тут, але тема далеко ще не вичерпана. Аналогічна ситуація з переходом від ізогамії до оогамії (і, взагалі, від злиття при заплідненні одноклітинних організмів до використання спеціалізованої стадії розвитку — гамет).
Розмножуватись інтенсивно і жити недовго, чи розмножуватись стримано і намагатися жити довго? Внутрішньопопуляційні онтогенетичні стратегії у постійному чи мінливому, сприятливому чи суворому середовищі. У якому разі вони працюють ефективніше: за великої спадкової компоненти у визначенні стратегії чи залежності стратегії від короткострокових умов?
Зв'язок між вибагливостю однієї статі та мінливостю іншої.
Гра, що обговорюється в цьому тексті; розподіл її результатів.
Чому всі євреї є нащадками царя Соломона, а всі українці — Ярослава Мудрого? Яка ймовірність того, що якийсь із членів репродуктивної сукупності, який жив певну кількість років тому і залишив нащадків, є предком усіх сучасних членів цієї сукупності?
Партеногенез у хлистохвостих ящірок
Перш за все, пропоную подивитися наступний фрагмент фільму. Йдеться про хлистохвостих ящірок.
Як можна зрозуміти, йдеться про популяції з клональним розмноженням, де нащадки генетично ідентичні своїм батькам. Усі особини в ній — самки, але вони здатні під впливом сплеску чоловічих гормонів виконувати чоловічу роль, імітуючи спарювання. Така імітація спарювання необхідна для самок, щоб у них відбувалося дозрівання яєць. У зв’язку з цим фрагментом фільму виникає безліч питань.
1. Якщо в популяції таких ящірок виникне особина, не здатна зазнавати сплеску чоловічих гормонів і допомагати розмножуватися іншим самкам, чи буде добір сприяти цій особині і її клональним нащадкам? Це видається досить імовірним. Як мінімум, її нащадки не будуть зазнавати конкуренції з боку нащадків інших клонів. У загальному випадку відносини між самками, які допомагають іншим особинам вагітніти, і тими, які цього не роблять, можна розглядати як відносини між "альтруїстами" і "егоїстами". Чи завжди добір буде сприяти "егоїстам"? Чи існують умови, в яких переважно будуть поширюватися "альтруїсти"?
2. Як міг відбутися перехід від нормального розмноження з двома статями, рекомбінацією і заплідненням, до варіанту, показаного в даному відеофрагменті? Для цього, ймовірно, знадобилося закріплення у клонів одразу двох незвичних особливостей: здатності до партеногенетичного розвитку яйцеклітин і здатності самок тимчасово переходити в гормональний стан, характерний для самців. Як міг відбуватися такий перехід?
Хоровий спів зелених жаб
Усі ми знаємо, що зелені жаби, які сидять по берегах водойми, співають не хаотично, а хором: то разом кричать (перекрикуючи одна одну), то разом мовчать. Альфред Брем розповідає про старе німецьке повір’я, що в кожному ставі серед жаб є свій "хормейстер". Чи може хоровий спів жаб бути результатом самоорганізації, який не вимагає жодного "хормейстера"?
Один зі способів відповіді на це питання — побудова моделі. Наприклад, для побудови простої моделі можна скористатися гідравлічною моделлю інстинктивної поведінки, запропонованою Конрадом Лоренцем. Незважаючи на критику цієї моделі, вона в простій формі відображає кілька найпростіших спостережень. Так, у тому випадку, якщо тварина в ході своєї життєдіяльності повинна переходити до певної форми поведінки, на кожному кроці вдається спостерігати наступне. Перехід до очікуваної поведінки часто є відповіддю на певний стимул (релізер). Чим довше тварина очікує цього релізера, тим чутливішою до нього вона виявляється. Якщо тварина перебуває в очікуванні стимулу-релізера, а його немає, вона може переходити до відповідної форми поведінки самовільно.
Простою моделлю (!), що описує ці закономірності, і є гідравлічна модель Лоренца. У ній описані особливості інстинктивної поведінки пояснюються накопиченням у тварині якоїсь специфічної енергії. Щоб прояснити гідравлічну модель, можна скористатися її описом (досить вдалим) з Вікіпедії.
Конрад Лоренц, зокрема, для пояснення феномена спрацьовування інстинктивних реакцій за відсутності специфічного подразника, запропонував оригінальну модель здійснення інстинктивної поведінки. Модель була побудована на основі принципів гідравліки і отримала назву «гідравлічної моделі Лоренца». У гідравлічній моделі специфічна енергія дії представлена водою, що безперервно тече в резервуар. Відтік води з резервуара перегороджений клапаном, до якого кріпиться вантаж. Вантаж позначає дію специфічних подразників, його вага прямо пропорційна інтенсивності подразника. Вода з резервуара може витекти у двох випадках: якщо сумарний тиск води і вантажу відкриють клапан, або рівень води перевищить допустимий і клапан відкриється під напором води.[13]
Таким чином, у міру накопичення специфічного потенціалу дії знижується поріг, необхідний для запуску відповідної інстинктивної реакції. У випадку, коли необхідний подразник відсутній протягом тривалого часу, реакція може здійснитися «вхолосту», без специфічного стимулу.
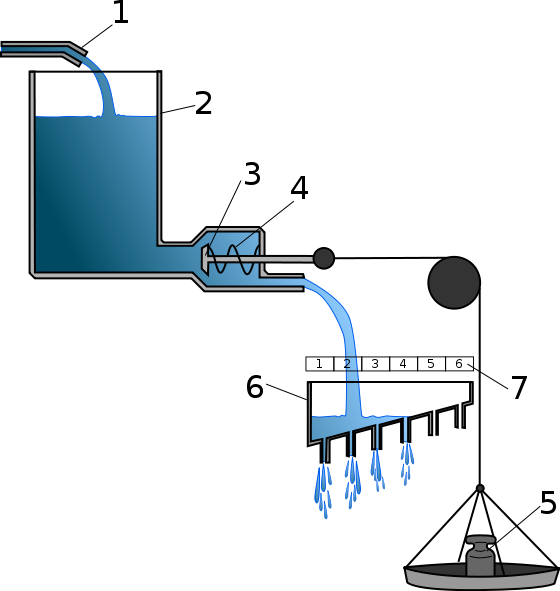
1-приплив специфічної енергії дії (СЕД) створює тиск у резервуарі (2)
3,4 - Вроджений спусковий механізм. Зазнає тиску з боку СЕД (3) і релізера (5)
Лоренц сам визнавав обмеженість гідравлічної моделі. Хоча вона і пояснює деякі спостережувані феномени, але має ряд серйозних недоліків. Перш за все, вона не враховує мінливість поведінки. По-друге, вона передбачає існування деякої зайвої сутності — гіпотетичної енергії. Слід зазначити, що нервова структура, відповідальна за накопичення специфічного потенціалу дії, не була виявлена. По-третє, є факти, що суперечать даній теорії.
Завдяки своїй простоті і наочності, гідравлічна теорія вивчається в університетських курсах зоопсихології до цього дня.
Підкреслю, що перелічені у статті Вікіпедії недоліки гідравлічної моделі в жодному разі не є протипоказаннями для її використання в моделюванні. Не треба вимагати від моделі, щоб вона була ідентична оригіналу за своєю структурою; якщо модель забезпечує ту ж поведінку, яка характерна для оригіналу, її використання (поки вона корисна) виявляється цілком виправданим. Описана Лоренцем модель приводить до реалізації тих особливостей поведінки (підвищення готовності прореагувати на релізер з часом і здатність до спонтанного включення певної форми інстинктивної поведінки), які були описані в багатьох експериментах.
До речі, ви ж зрозуміли, що на схемі з Вікіпедії позначено цифрами 6 і 7? Реалізація інстинктивної поведінки і її інтенсивність.
Отже, я пропоную зробити імітаційну модель, що дозволяє перевірити, чи пояснює гідравлічна модель Лоренца феномен хорового співу жаб. Розглянемо круглу водойму, по периметру якої рівномірно сидять жаби. Перехід жаб від мовчання до співу пояснюється на наведеному нижче малюнку (що відображає концептуальну модель, яка описує перехід жаб від мовчання до співу і назад).
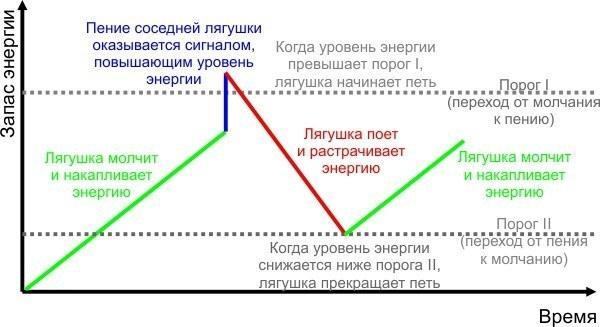
Як випливає з моделі, для опису поведінки жаб достатньо наступних основних параметрів:
— швидкість накопичення енергії у мовчазної жаби (на скільки одиниць підвищується рівень енергії за кожен крок моделі);
— швидкість зниження енергії у співаючої жаби;
— порогове значення енергії для переходу жаби від мовчання до співу;
— порогове значення енергії для переходу жаби від співу до мовчання (цей поріг нижчий за попередній);
— ефект співу сусідніх жаб (підвищення енергії у жаби, яка на попередньому кроці моделі почула спів іншої жаби) залежно від відстані до сусідки (ймовірно, ефект співу найближчої сусідки найвищий, сусідки через одну — нижчий, а через дві і через три — ще нижчий).
Ймовірно, слід передбачити ще мінливість деякої випадкової величини, від якої залежить значення порогів "включення" і "виключення" у кожної конкретної особини.
Фактично, для кожного кроку для кожної особини треба проводити наступні обчислення:
1. Визначається, скільки енергії було у особини на попередньому кроці, а також мовчала вона чи співала. Якщо мовчала, її енергія збільшується (відповідно до заданої загальної швидкості збільшення енергії у мовчазної жаби, з урахуванням індивідуальної поправки, що залежить від випадкового числа.
2. Обчислюється сила зовнішнього стимулу. До досягнутого рівня енергії додається його збільшення від співу найближчих сусідів, з понижуючим коефіцієнтом — від сусідів "через одного", і з ще нижчим — від сусідів "через два"...
3. Якщо рівень енергії мовчазної жаби з добавкою на стимулювання сусідами перевищує поріг "включення" (ймовірно, з урахуванням індивідуальної поправки, що залежить від випадкового числа) — жаба починає співати.
4. Якщо жаба мовчить, її енергія знижується (відповідно до заданої загальної швидкості зниження енергії у співаючої жаби, з урахуванням індивідуальної поправки, що залежить від випадкового числа).
5. Якщо рівень енергії співаючої жаби з добавкою на стимулювання сусідами виявляється нижчим за поріг "виключення" (ймовірно, з урахуванням індивідуальної поправки, що залежить від випадкового числа) — жаба припиняє співати.
Щойно перелічені п’ять кроків є застосуванням концептуальної моделі управління співом жаби (показаної на схемі) до алгоритму такого управління в моделі.
Найпростішим виходом моделі може бути динаміка кількості співаючих жаб на кожному кроці. Можна також візуалізувати розподіл співаючих і мовчазних жаб по периметру ставу.