Creation of Worlds — 11. Ideas for Modeling
{ "title": "Models that would be worth making over time...", "summary": "", "body": "Модели, которые со временем стоило бы сделать... BATRIMG1>BATR BATRIMG2>BATR" }
Examples of various models, their concept and design
Ideas for modelling
A biosphere consisting of ecosystems that can be in stable or unstable states. Under what conditions will such a biosphere evolve towards greater stability without Darwinian selection and reproduction of individual units?
The problem makes sense, for example, from the perspective of the discussion between J. Lovelock and R. Dawkins. Lovelock's Gaia concept implies planetary regulation mechanisms. Purposefulness is the result of selection. Dawkins argues that selection is always associated with differential reproduction of replicators. If there were many biospheres, some of which died out while others multiplied, then, according to Dawkins, biospheres could develop the ability to react purposefully. According to an assumption that can be tested by simulation, purposefulness can be a consequence of another type of selection, which can be called Boltzmann selection - the spread of more thermodynamically probable (more stable) states.
Analogy. Can selection at the organism level be the cause of purposefulness of population regulation mechanisms without Darwinian selection (differential death and differential reproduction, i.e., resettlement) of populations?
Will the population, which exists in an unlimited two-dimensional space and is able to resettle in unoccupied directions at a certain speed, grow hyperbolically (like the population of humanity according to von Foerster)? More precisely, according to what model will it grow (will the relative increase in the area of the range decrease)?
Competition for a common resource of two populations. The number of one is controlled by predators, the number of the other - by the lack of resources. Under what conditions will predators ensure the "victory" of their victims over their competitors?
Impact of overpopulation crisis on the environment capacity. Elephants feeding on acacia in the savannah. When there is enough food, they eat leaves; when there is not enough, they break stems and strip cambium. Extension of the model - the impact of homotherms and smilodonts on this process.
Dennett's creatures! More details here and here. Skinner's creature, which will rejoice in wins and be sad in case of losses.
Transition of a population of birds living on a continent stretched along the meridian from sedentary to migratory. In high latitudes, better conditions for reproduction, in low latitudes - higher chances of surviving the winter.
Regulation of the annual cycle in a population living in a temperate climate: what is better to react to, to changes in temperature or daylight hours? Compare different ways of regulation by the number of "false" days under different levels of climate variability.
Male and female reproductive strategies of humans.
Different planning horizons under different levels of environmental variability.
Competition of two populations with sexual reproduction and two sexes under stabilizing selection. In one, variability and mortality (which takes away individuals that deviate from the norm) are the same for males and females, in the other - males are more variable and, therefore, suffer more from negative selection. V. Geodakyan's idea.
Comparison of the rate of adaptation development in the case of clonal and sexual reproduction.
Comparison of the spread of new traits in genetic and cultural inheritance.
Phase transition in evolution (from genes to memes, from memes to t-memes, and further).
Slow and fast infections under conditions of different population structures.
Fisher's principle at different "prices" of individuals of different sexes or in the case of a change in the sex ratio during ontogenesis.
Simulation proof of the Monty Hall paradox. A model that demonstrates the success of two different strategies (to change the choice or not to change) can be made by analogy with a model about the birth of boys and girls (which solves one of the well-known demographic problems).
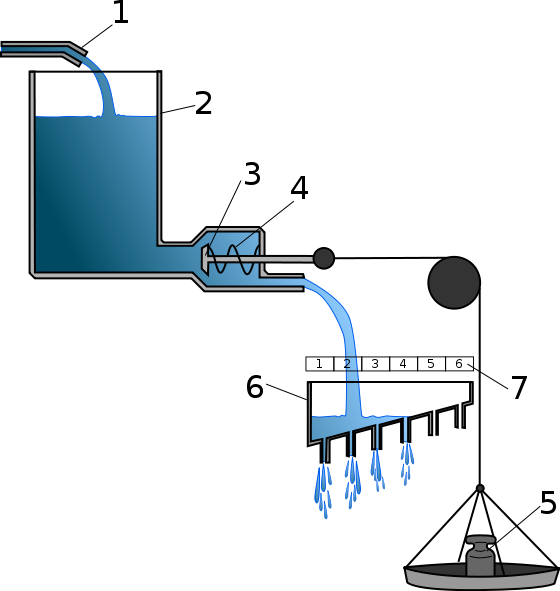
Lorenz himself recognized the limitations of the hydraulic model. Although it explains some observed phenomena, it has a number of serious drawbacks. First of all, it does not take into account the variability of behavior. Secondly, it assumes the existence of some extra entity - hypothetical energy. It should be noted that the nervous structure responsible for the accumulation of specific action potential has not been found. Thirdly, there are facts that contradict this theory.
Due to its simplicity and clarity, the hydraulic theory is still studied in university courses of zoopsychology.
I propose to make a simulation model that allows testing whether the Lorenz hydraulic model explains the phenomenon of chorus singing of frogs. Consider a round pond, along the perimeter of which frogs sit evenly. The transition of frogs from silence to singing is explained by the figure below (reflecting a conceptual model that describes the transition of frogs from silence to singing and back).
As can be seen from the model, to describe the behavior of frogs, the following basic parameters are sufficient:
- the rate of accumulation of energy in a silent frog (by how many units the energy level increases per each model step);
- the rate of decrease in energy in a singing frog;
- the threshold energy value for the transition of a frog from silence to singing;
- the threshold energy value for the transition of a frog from singing to silence (this threshold is lower than the previous one);
- the effect of singing neighboring frogs (increase in energy in a frog that heard the singing of another frog at the previous model step) depending on the distance to the neighbor (probably, the effect of singing of the nearest neighbor is the highest, the neighbor through one - lower, and through two and through three - even lower).
Probably, it is worth providing for the variability of a certain random variable, on which the value of the "on" and "off" thresholds depends for each specific individual.
In fact, for each step, for each individual, it is necessary to carry out the following calculations:
1. Determine how much energy an individual had at the previous step, as well as whether it was silent or singing. If it was silent, its energy increases (in accordance with a given general rate of energy increase in a silent frog, taking into account an individual correction that depends on a random number).
2. Calculate the strength of the external stimulus. To the achieved energy level, add its increase from the singing of neighboring frogs, with a decreasing coefficient - from neighbors "through one", and with an even lower coefficient - from neighbors "through two"...
3. If the energy level of a silent frog with an addition to stimulation by neighbors exceeds the "on" threshold (probably taking into account an individual correction that depends on a random number) - the frog starts singing.
4. If the frog is silent, its energy decreases (in accordance with a given general rate of energy decrease in a singing frog, taking into account an individual correction that depends on a random number).
5. If the energy level of a singing frog with an addition to stimulation by neighbors turns out to be below the "off" threshold (probably taking into account an individual correction that depends on a random number) - the frog stops singing.
The simplest output of the model can be the dynamics of the number of singing frogs at each step. It is also possible to visualize the distribution of singing and silent frogs along the perimeter of the pond.