Усова (2014) Скелетохронологічне вивчення росту зелених жаб НПП «Гомільшанські ліси»
Усова О. Є. Скелетохронологічне вивчення росту зелених жаб зі ставків Національного природного парку «Гомільшанські ліси» та його околиць // Біологія та валеологія. — 2014. — Вип. 16. — С. 51–61.
УДК: (57.022+57.032):591.84:591.158.1:597.851
http://orcid.org/0000-0001-5688-398X
СКЕЛЕТОХРОНОЛОГІЧНЕ ВИВЧЕННЯ РОСТУ ЗЕЛЕНИХ ЖАБ ЗІ СТАВКІВ НАЦІОНАЛЬНОГО ПРИРОДНОГО ПАРКУ «ГОМІЛЬШАНСЬКІ ЛІСИ» ТА ЙОГО ОКОЛИЦЬ
Усова О.Є.
Харківський національний університет імені В.Н. Каразіна
Визначено вік та темпи росту 265 особин зелених жаб (71 Pelophylax ridibundus, 186 диплоїдних та 8 триплоїдних Pelophylaxesculentus) із 5 ставків, що розташовані у Національному природному парку «Гомільшанські ліси» та поруч із ним. У роботі зроблено припущення, що відмінності у темпах росту зелених жаб (які не пов’язані з їх формою, тобто складом геномів) пояснюються різними внутрішньовидовими онтогенетичними стратегіями: скоростиглістю та тугорослістю. Відмінності між цими стратегіями є особливо контрастними для самок. Найстарші за віком особини є тугорослими; найкрупніші за розміром – скоростиглими. Протягом життя більшість самок зберігає певну стратегію росту, але деякі особини, що відповідають за розмірами тугорослим, здатні значно прискорити свій ріст та наздогнати скоростиглих.
Ключові слова: Pelophylax esculentus complex, Pelophylax ridibundus, Сіверсько-Донецький центр різноманіття зелених жаб, скелетохронологія, онтогенетичні стратегії, ріст.
The skeletochronological study of growth of green frogs from ponds situated in the National Nature Park "Gomolshansky lissy" and its environs. Usova O.Ye. – The age and growth rate of 265 green frog individuals (71 Pelophylax ridibundus, 186 diploid and 8 triploid Pelophylax esculentus) from 5 ponds located in the National Nature Park "Gomilshansky lissy" and its environs were determined. It is suggested that differences in growth rates of green frogs (not related to their form, i.e. to composition of their genomes) can be explained by different intraspecific ontogenetic strategies: precocity and stuntedity. The differences between these strategies are especially contrasting in females. The eldest individuals are stunted; those who have the largest size are precocious. Over a lifetime, most females retain a certain growth strategy, but some individuals, whose size is corresponding to the stunted ones, can greatly accelerate its growth and catch up with the precocious ones.
Key words: Pelophylax esculentus complex, Pelophylax ridibundus, Seversko-Donetskiy center of green frogs diversity, skeletochronology, ontogenetic strategies, growth.
ВСТУП
До гібридогенного комплексу зелених жаб Pelophylax esculentus complex (=Rana esculenta complex) належать два батьківських види Pelophylax lessonae (Camerano, 1882) та Pelophylax ridibundus (Pallas, 1771), а також їх різноманітні ди-, три- та тетраплоїдні гібриди (Plötner, 2005), для яких ми використовуємо (8) назву, аналогічну видовій, Pelophylax esculentus (Linnaeus, 1758). Відтворення гібридів підтримується завдяки геміклональному спадкуванню (утворення ними статевих клітин, що клонально, без рекомбінації, передають певні батьківські геноми). Сукупності представників батьківських видів і різноманітних гібридів, що поєднуються спільним відтворенням та передачею з покоління у покоління як клональних, так і рекомбінантних геномів, отримали назву ГПС – геміклональних популяційних систем (8; 10). Найскладніші із відомих ГПС зареєстровано в басейні верхньої течії р. Сіверський Донець, у регіоні, що названий Сіверсько-Донецьким центром різноманіття зелених жаб (8; 10). Найрізноманітніші ГПС Pelophylax esculentus complex знайдено у Національному природному парку «Гомільшанські ліси», що розташований у Зміївському районі Харківської області (2).
У численних роботах зареєстровано, що особини в популяціях хребетних тварин можуть демонструвати різні стратегії росту. Ці стратегії запропоновано називати скоростиглістю та тугорослістю (9). Задачею цієї роботи є аналіз того, чи проявляються названі стратегії в рості представників зелених жаб. Щоб відповісти на це питання, ми застосували скелетохронологічне дослідження.
МАТЕРІАЛИ ТА МЕТОДИ ДОСЛІДЖЕНЬ
У ході роботи визначено вік та темпи росту 265 зелених жаб, склад яких охарактеризовано у таблиці 1. Ці жаби належать до одного з батьківських видів (P. ridibundus) та до міжвидових гібридів, відносно яких застосовується подібна до видової назва P. esculentus. Серед гібридів були як диплоїдні, так і триплоїдні особини. Далі P. ridibundus, диплоїдні та триплоїдні P. esculentus розглядаються як форми P. esculentus complex (слово «форма» застосовується тут у тому сенсі, який йому надавав Е. Майр (3), без припущень щодо статусу цих особин).
Всього у досліджуваний матеріал входив 71 представник батьківського виду (P. ridibundus) та 194 гібриди (P. esculentus), з яких 186 – диплоїдів та 8 триплоїдів. Досліджувані жаби спіймані у період з 2004 року по 2014 рік в Харківській області.
Генотипи 57 особин визначені за допомогою протічної флуоресцентної ДНК-цитометрії. Ці дослідження виконували С. М. Литвинчук та Ю. М. Розанов в Інституті цитології РАН (м. Санкт-Петербург, Росія). Із цих 57 особин 5 були триплоїдними; одна з них мала генотип LLR (два геноми P. lessonae і один – P. ridibundus) і чотири – LRR (один геном P. lessonae і два – P. ridibundus). Інші досліджені нами триплоїди визначені за розміром їх еритроцитів (1). Склад геномів цих триплоїдів не визначався.
Визначення P. ridibundus і P. esculentus серед диплоїдних особин проводилося за комплексом їх зовнішніх ознак (7). Як довів попередній досвід, серед диплоїдних особин таке визначення є майже безпомилковим. Непевно визначаються за зовнішніми ознаками триплоїдні особини, яких, втім, можна визначити за розміром їх клітин.
Таблиця 1
Склад дослідженого матеріалу
|
Місце збору |
Форма зелених жаб |
Незрілі |
Самки |
Самці |
Разом |
|
Нижній Добрицький ставок N 49°33'24" E 36°18'34" |
Pelophylax ridibundus |
4 |
13 |
2 |
19 |
|
Pelophylax esculentus, 2n |
6 |
28 |
50 |
84 |
|
|
Pelophylax esculentus, 3n |
— |
2 |
— |
2 |
|
|
Разом |
10 |
43 |
52 |
105 |
|
|
Верхній Добрицький ставок N 49°34'18" E 36°16'24" |
Pelophylax ridibundus |
— |
— |
— |
— |
|
Pelophylax esculentus, 2n |
— |
2 |
4 |
6 |
|
|
Pelophylax esculentus, 3n |
— |
— |
1 |
1 |
|
|
Разом |
— |
2 |
5 |
7 |
|
|
Cтавок у Сухій Гомольші N 49°32'17" E 36°20'19" |
Pelophylax ridibundus |
— |
1 |
1 |
2 |
|
Pelophylax esculentus, 2n |
— |
3 |
4 |
7 |
|
|
Pelophylax esculentus, 3n |
— |
3 |
— |
3 |
|
|
Разом |
— |
7 |
5 |
12 |
|
|
Іськів ставок N 49°37'33" E 36°17'9" |
Pelophylax ridibundus |
19 |
28 |
1 |
48 |
|
Pelophylax esculentus, 2n |
— |
15 |
71 |
86 |
|
|
Pelophylax esculentus, 3n |
— |
1 |
1 |
2 |
|
|
Разом |
19 |
44 |
73 |
136 |
|
|
Коряків яр N 49°36'56" E 36°18'43" |
Pelophylax ridibundus |
— |
2 |
— |
2 |
|
Pelophylax esculentus, 2n |
— |
— |
3 |
3 |
|
|
Pelophylax esculentus, 3n |
— |
— |
— |
— |
|
|
Разом |
— |
2 |
3 |
5 |
|
|
Разом |
29 |
98 |
138 |
265 |
|
У ході скелетохронологічного дослідження четверту фалангу найдовшого пальця задньої ноги жаб очищали від м’яких тканин, декальцінували, отримували на заморожуючому мікротомі зрізи в середній частині діафіза товщиною 20-22 мкм, фарбували гематоксиліном Ерліха, заливали гліцерином, накривали покривним склом, фотографували тимчасові препарати, а потім проводили вимірювання та підрахунки ліній склеювання за фотографіями з використанням програми Adobe Photoshop CS5. Результати вимірювань переводили в мікрометри з використанням коефіцієнта перерахунку, визначеного за допомогою фотографії об’єкт-мікрометра, зробленої за тих же умов, що і фотографії препаратів.
Методика інтерпретації препаратів, визначення віку жаб та ретроспективного обчислення їх приросту в різні роки життя докладно описана в іншій публікації (6). Результати вимірювань заносили в електронну базу даних, виконану у програмі Statistica 8, StatSoft Inc.; у цій базі проводили подальшу обробку результатів.
Можливість скелетохронологічного визначення віку та швидкості росту жаб є наслідком того, що зростання цих тварин (як і інших пойкілотермних хребетних помірного клімату) є нерівномірним (4). Під час зимових зупинок росту у кістках виникають лінії склеювання, що є добре помітними на фарбованих зрізах. Інтерпретація цих зрізів ускладнюється тим, що з певного віку всередині трубчастих кісток починається розширення ендостальної порожнини. Ця порожнина може зруйнувати (резорбувати) ту чи іншу кількість ліній склеювання, або їх частину, в періості. З часом розростання ендостальної порожнини зупиняється і починається ріст ендостальної кісткової тканини у напрямку від межі періосту і ендосту до середини ендостальної порожнини. Внаслідок цього при інтерпретації препаратів важливо знайти на зрізах межу періосту і ендосту, порахувати лінії склеювання в періості та припустити, скільки таких ліній зникло під час розширення ендостальної порожнини.
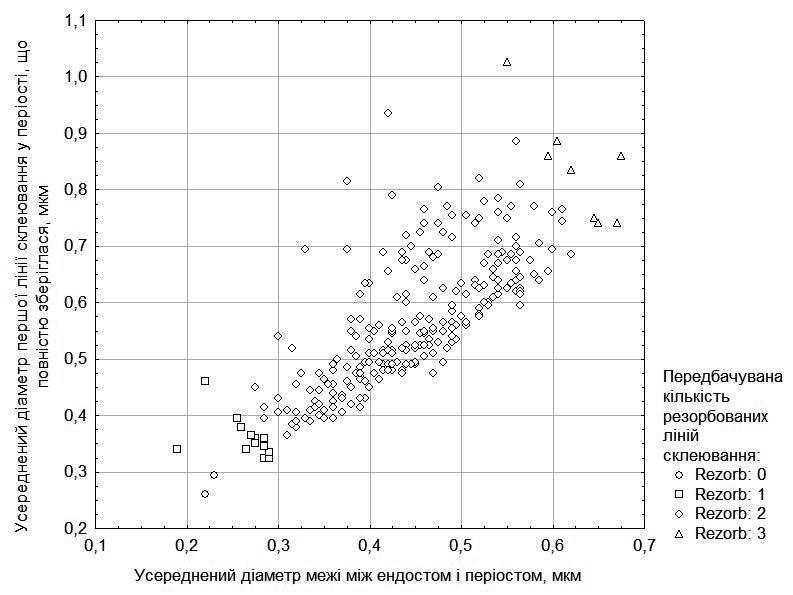
Щоб зробити припущення щодо кількості резорбованих періостальних ліній склеювання, слід зважати, який діаметр має ендостальна порожнина (враховуючи і ендост) та перша лінія у періості, що повністю зберіглася (ці дані показані на рис. 1). У ряді випадків враховується кількість частково резорбованих ліній склеювання у періості. Спираючись на припущення щодо кількості резорбованих ліній, ми визначили вірогідний вік досліджених особин. Його розподіл показано в табл. 2.
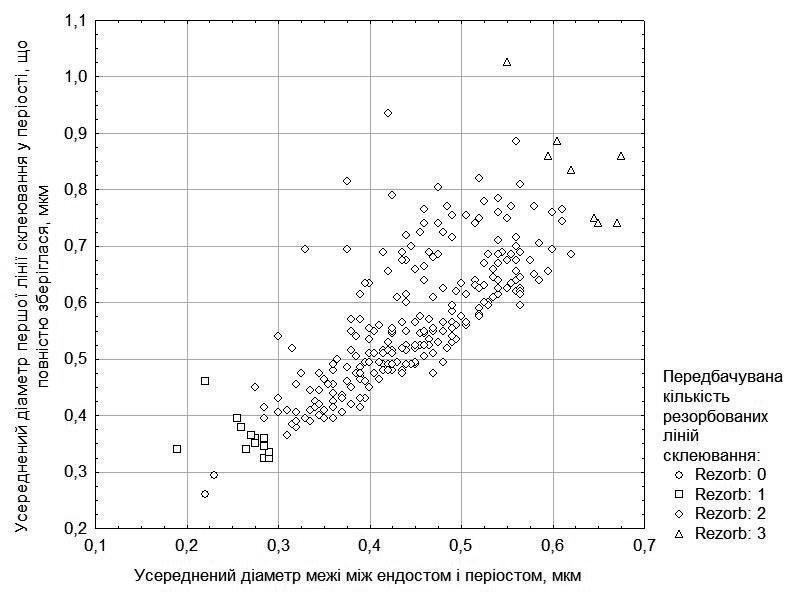
Рис. 1. Розподіл вивчених особин за розмірами ендостальної порожнини та першої повністю збереженої лінії склеювання, а також прийняті на цій основі припущення щодо кількості резорбованих ліній
Таблиця 2
Вік досліджених особин
|
Форма |
Стать |
Вік |
|||||||||
|
2+ |
3+ |
4+ |
5+ |
6+ |
7+ |
8+ |
9+ |
10+ |
Разом |
||
|
P. ridibundus |
Незрілі |
3 |
20 |
— |
— |
— |
— |
— |
— |
— |
23 |
|
Самки |
— |
1 |
23 |
11 |
— |
6 |
2 |
— |
1 |
44 |
|
|
Самці |
— |
— |
— |
2 |
1 |
1 |
— |
— |
— |
4 |
|
|
Разом |
3 |
21 |
23 |
13 |
1 |
7 |
2 |
— |
1 |
71 |
|
|
P. esculentus, 2 n |
Незрілі |
2 |
4 |
— |
— |
— |
— |
— |
— |
— |
6 |
|
Самки |
— |
2 |
12 |
14 |
8 |
6 |
1 |
5 |
— |
48 |
|
|
Самці |
— |
3 |
29 |
48 |
41 |
7 |
4 |
— |
— |
132 |
|
|
Разом |
2 |
9 |
41 |
62 |
49 |
13 |
5 |
5 |
— |
186 |
|
|
P. esculentus, 3 n |
Незрілі |
— |
— |
— |
— |
— |
— |
— |
— |
— |
— |
|
Самки |
— |
— |
3 |
3 |
— |
— |
— |
— |
— |
6 |
|
|
Самці |
— |
— |
1 |
— |
— |
1 |
— |
— |
— |
2 |
|
|
Разом |
— |
— |
4 |
3 |
— |
1 |
— |
— |
— |
8 |
|
|
Разом |
5 |
30 |
68 |
78 |
50 |
21 |
7 |
5 |
1 |
265 |
|
Дані щодо росту трьох досліджених форм P. esculentus complex обробляли спільно (окремо – самці, а самки разом зі статево незрілими особинами). Це є наслідком того, що у нашому дослідженні, як і в попередніх (5; 6), відмінність між цими формами виявилася незначною.
РЕЗУЛЬТАТИ ДОСЛІДЖЕНЬ ТА ЇХ ОБГОВОРЕННЯ
Аналіз росту жаб простіше за все почати з розподілу вивчених особин за розміром (рис. 2). Він має досить характерний вигляд. Розподіл самок виразно бімодальний, серед них багато дрібних (менших за 60 мм) та великих (більших за 80 мм), а середніх за розміром самок відносно мало. Цей розподіл не пов’язаний ані з формою жаб, ані з їх походженням із того або іншого з досліджених ставків.
Розподіл самців, хоча також відрізняється від нормального, має один головний пік.
Щоб врахувати вплив віку жаб, розглянемо залежність розміру від кількості зимівель окремо для самок (рис. 3) та самців (рис. 4).
На рис. 3 можна побачити, що самки виразно поділяються на тих, які швидко ростуть (їх вік не перевищує 7 років), та тих, що ростуть значно повільніше. Триплоїдних гібридів серед досліджених самок відносно небагато і усі вони (вірогідно, випадково) відносно молоді. Але на рис. 3 можна побачити, що особини, які ростуть швидко і повільно, є як серед представників батьківського виду, так і серед диплоїдних гібридів.
Поділ за швидкістю росту серед самців (рис. 4) є менш виразним: для особин кожного віку більшість особин характеризується розмірами, близьким до середніх. Втім, найкрупніші особини переважно мають вік до 5 років, а більшість найстарших особин менші за розміром.
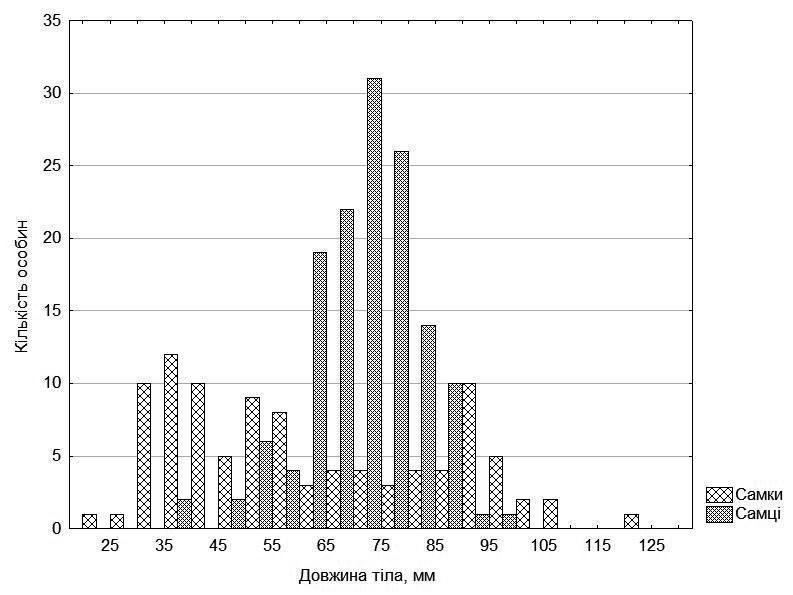
Рис. 2. Розподіл за розміром вивчених особин
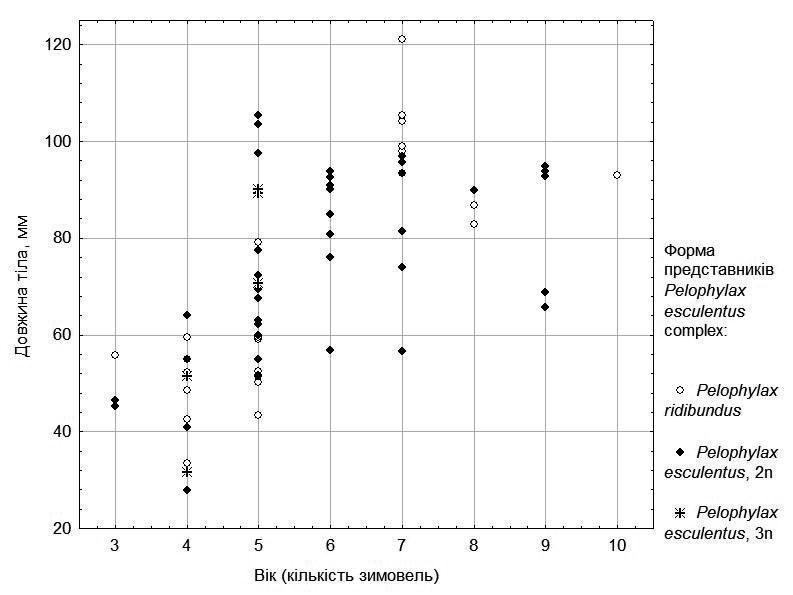
Рис. 3. Залежність розміру від віку для самок з п’яти ставків у околицях НПП «Гомільшанські ліси»
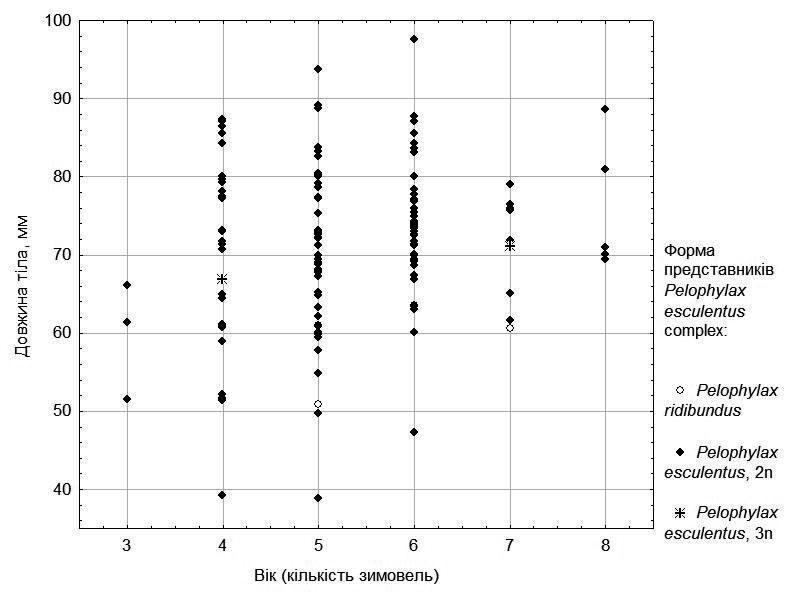
Рис. 4. Залежність розміру від віку для самців з п’яти ставків у околицях НПП «Гомільшанські ліси»
Можна припустити, що ці відмінності за швидкістю росту відбивають різницю між внутришньопопуляційними онтогенетичними стратегіями скоростиглості та тугорослості (9). Це припущення відображене на рис. 5 для самок та на рис. 6 (зі значно більшою часткою невизначеності) – для самців.
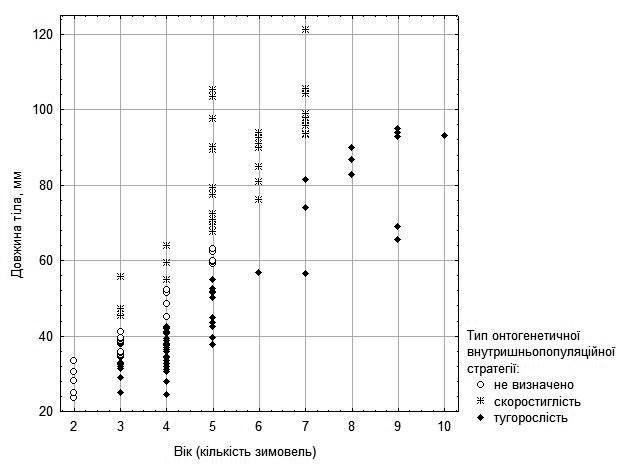
Рис. 5. Припущення щодо вірогідних онтогенетичних стратегій самок та статевонезрілих особин, які можна зробити на основі їх віку та розмірів
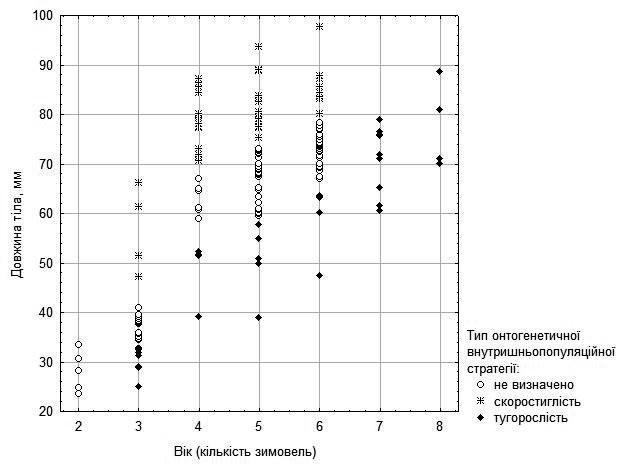
Рис. 6. Припущення щодо вірогідних онтогенетичних стратегій самців та статевонезрілих особин, які можна зробити на основі їх віку та розмірів
Цікаво встановити, чи може стратегія росту змінюватися протягом життя. Ми порівняли тип стратегії кожної особини, який визначений за її розміром та віком, з оцінками її росту протягом життя, що спираються на ретроспективне встановлення змін її розмірів із віком за даними скелетохронології (рис. 7).
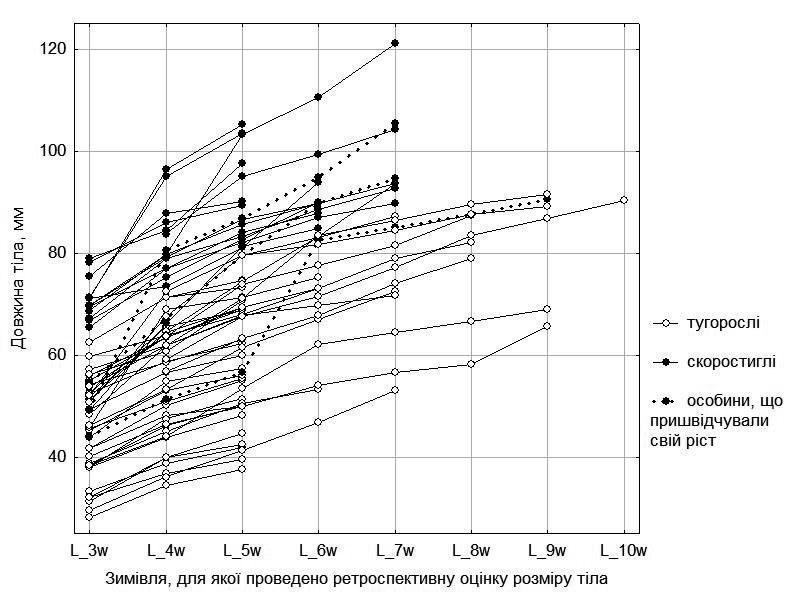
Рис. 7. Динаміка росту самок з п’яти ставків у околицях НПП «Гомільшанські ліси»
На рис. 7 можна побачити, що вірогідний тип стратегії росту, що є характерним для кожної особини, зберігається протягом тієї частини онтогенезу, яка доступна для нашого вивчення. Втім, серед самок є три особини, яких у 3 роки можна було вважати тугорослими, але після цього вони пришвидшили свій ріст і перейшли до скоростиглих (2 особини) або наблизилися до них (1 особина).
ВИСНОВКИ
У роботі зроблено припущення, що відмінності у темпах росту зелених жаб (що не пов’язані з їх формою, тобто складом геномів) пояснюються різними внутрішньовидовими онтогенетичними стратегіями: скоростиглістю та тугорослістю. Відмінності між цими стратегіями є особливо контрастними для самок. Найстарші за віком особини є тугорослими; найкрупніші за розміром — скоростиглими. Протягом життя більшість самок зберігає певну стратегію росту, але деякі особини здатні значно прискорити своє зростання та перейти з тугорослих до скоростиглих.
Подяки. Автор щиро вдячна С. М. Литвинчуку та Ю. М. Розанову за визначення форм зелених жаб із застосуванням проточної ДНК-цитометрії. Неоцінену допомогу вона отримала від консультацій Е. М. Сміріної щодо отримання та інтерпретації препаратів зрізів кісток. Виконання цієї роботи стало можливим завдяки допомозі членів групи вивчення популяційної екології амфібій кафедри зоології та екології тварин Харківського національного університету імені В. Н. Каразіна: О. В. Коршунова, М. О. Кравченко, О. В. Бірюк, О. В. Мелешко, М. В. Строїлова, К. П. Віноградової та інших. Особливої подяки заслуговує Д. А. Шабанов як ініціатор та науковий керівник цієї роботи.
Література
- Бондарева А. А., Бибик Ю. С., Самило С. М., Шабанов Д. А. Цитогенетичні особливості еритроцитів зелених жаб із Сіверсько-Донецького центру різноманіття Pelophylax esculentus complex / А. А. Бондарева, Ю. С. Бибик, С. М. Самило , Д. А. Шабанов // Вісник Харківського національного університету імені В. Н. Каразіна. Серія: біологія. – 2012. – Вип. 15 (№ 1008). – С. 116-123.
- Коршунов А. В. Екологічні особливості біотопічного розподілу представників Pelophylax esculentus complex у Харківській області / А. В. Коршунов // Вісник Харківського національного університету імені В. Н. Каразіна. Серія: біологія. – 2008. – Вип. 8 (№ 828). – С. 48-57.
- Майр Е. Принципи зоологічної систематики / Е. Майр. – М. : Мир, 1971. – 454 с.
- Смирина Е. М. Прижиттєве визначення віку і ретроспективна оцінка розмірів тіла сірої ропухи (Bufo bufo) / Е. М. Смирина // Зоол. журн. – 1983. – Т. 63, № 3. – С. 437-444.
- Усова Е. Е. Визначення природної смертності статевозрілих зелених жаб (Pelophylax esculentus complex; Amphibia, Ranidae) з використанням скелетохронології / Е. Е. Усова // Вісник Харківського національного університету імені В. Н. Каразіна. Серія: біологія. – 2010. – Вип. 12 (№ 920). – С. 104-110.
- Усова Е. Е. Вік і швидкість росту зелених жаб (Pelophylax esculentus complex) Нижнього Добрицького ставка (Зміївський район Харківської області / Е. Е. Усова // Вісник Харківського національного університету імені В. Н. Каразіна. Серія: біологія. – 2014 – Вип. 20, № 1100. – С. 204–212.
- Шабанов Д. А., Зиненко А. И., Коршунов А. В., Кравченко М. А., Мазепа Г. А. Вивчення популяційних систем зелених жаб (Rana esculenta complex) у Харківській області: історія, сучасний стан і перспективи / Д. А. Шабанов, А. И. Зиненко, А. В. Коршунов, М. А. Кравченко, Г. А. Мазепа // Вісник Харківського національного університету імені В. Н. Каразіна. Серія: біологія. – 2006. – Випуск 3 (№ 729). – С. 208-220.
- Шабанов Д. А., Коршунов О. В., Кравченко М. О. Які ж зелені жаби населяють Харківську область? Термінологічний і номенклатурний аспекти проблеми / Д. А. Шабанов, А. В. Коршунов, М. А. Кравченко // Біологія та валеологія. – Вип. 11. – Харків: ХДПУ, 2009. – С. 164-125.
- Шабанов Д. А., Коршунов А. В., Кравченко М. А., Шабанова А. В., Усова Е. Е. Внутрішньопопуляційні онтогенетичні стратегії скоростиглості і тугорослості: визначення на прикладі безхвостих амфібій / Д. А. Шабанов, А. В. Коршунов, М. А. Кравченко, А. В. Шабанова, Е. Е. Усова // Вісник Харківського національного університету імені В. Н. Каразіна. Серія: біологія. – 2014 (у друці).
- Шабанов Д. А., Литвинчук С. Н. Зелені жаби: життя без правил чи особливий спосіб еволюції? / Д. А. Шабанов, С. Н. Литвинчук // Природа. – 2010. – № 3 (1135). – С. 29-36.
Скелетохронологічне вивчення росту зелених жаб зі ставків Національного природного парку «Гомільшанські ліси» та його околиць. Усова Е.Е. – Визначено вік і темпи росту 265 особин зелених жаб (71 Pelophylax ridibundus, 186 диплоїдних і 8 триплоїдних Pelophylax esculentus) з 5 ставків, розташованих у Національному природному парку «Гомільшанські ліси» і поряд з ним. Висловлено припущення, що відмінності в темпах росту зелених жаб (не пов’язані з їх формою, тобто складом геномів) пояснюються різними внутрішньовидовими онтогенетичними стратегіями: скоростиглістю і тугорослістю. Відмінності між цими стратегіями особливо помітні для самок. Найстаріші за віком особини є тугорослими; найбільші за розміром – скоростиглими. Протягом життя більшість самок зберігає певну стратегію росту, але деякі особини, що відповідають за розмірами тугорослим, здатні значно прискорити свій ріст і наздогнати скоростиглих.
Ключові слова: Pelophylax esculentus complex, Pelophylax ridibundus, Сіверсько-Донецький центр різноманіття зелених жаб, скелетохронологія, онтогенетичні стратегії, ріст.