Usova (2014) Skeletochronological Study of Growth in Green Frogs from NPP "Homilshansky Forests"
Usova O. Ye. Skeletochronological study of growth in green frogs from ponds of the National Nature Park «Homilshansky Forests» and its environs // Biology and Valeology. — 2014. — Issue 16. — P. 51–61.
UDC: (57.022+57.032):591.84:591.158.1:597.851
http://orcid.org/0000-0001-5688-398X
SKELETOCHRONOLOGICAL STUDY OF GROWTH IN GREEN FROGS FROM PONDS OF THE NATIONAL NATURE PARK "HOMILSHANSKY FORESTS" AND ITS ENVIRONS
Usova O.Ye.
V. N. Karazin Kharkiv National University
The age and growth rates of 265 green frog individuals (71 Pelophylax ridibundus, 186 diploid and 8 triploid Pelophylax esculentus) from 5 ponds located in the National Nature Park "Homilshansky Forests" and its environs were determined. The study proposes that differences in growth rates of green frogs (unrelated to their form, i.e. genome composition) are explained by different intraspecific ontogenetic strategies: precocity and stunted growth. The differences between these strategies are especially pronounced in females. The oldest individuals are stunted growers; the largest individuals are precocious. Over their lifetime, most females retain a particular growth strategy, but some individuals whose size corresponds to stunted growers are capable of considerably accelerating their growth and catching up with the precocious ones.
Key words: Pelophylax esculentus complex, Pelophylax ridibundus, Seversky-Donetsk center of green frog diversity, skeletochronology, ontogenetic strategies, growth.
The skeletochronological study of growth of green frogs from ponds situated in the National Nature Park "Gomolshansky lissy" and its environs. Usova O.Ye. – The age and growth rate of 265 green frog individuals (71 Pelophylax ridibundus, 186 diploid and 8 triploid Pelophylax esculentus) from 5 ponds located in the National Nature Park "Gomilshansky lissy" and its environs were determined. It is suggested that differences in growth rates of green frogs (not related to their form, i.e. to composition of their genomes) can be explained by different intraspecific ontogenetic strategies: precocity and stuntedity. The differences between these strategies are especially contrasting in females. The eldest individuals are stunted; those who have the largest size are precocious. Over a lifetime, most females retain a certain growth strategy, but some individuals, whose size is corresponding to the stunted ones, can greatly accelerate its growth and catch up with the precocious ones.
Key words: Pelophylax esculentus complex, Pelophylax ridibundus, Seversko-Donetskiy center of green frogs diversity, skeletochronology, ontogenetic strategies, growth.
INTRODUCTION
The hybridogenetic complex of green frogs Pelophylax esculentus complex (=Rana esculenta complex) includes two parental species, Pelophylax lessonae (Camerano, 1882) and Pelophylax ridibundus (Pallas, 1771), as well as their diverse di-, tri-, and tetraploid hybrids (Plötner, 2005), for which we use (8) a species-level designation, Pelophylax esculentus (Linnaeus, 1758). Hybrid reproduction is maintained through hemiclonal inheritance (the formation of gametes that clonally, without recombination, transmit specific parental genomes). Assemblages of parental species representatives and diverse hybrids that are united by shared reproduction and the intergenerational transmission of both clonal and recombinant genomes have been termed HPS – hemiclonal population systems (8; 10). The most complex known HPS have been recorded in the upper Seversky Donets River basin, in a region designated the Seversky-Donetsk center of green frog diversity (8; 10). The most diverse HPS of Pelophylax esculentus complex have been found in the National Nature Park "Homilshansky Forests", located in the Zmiivsky district of Kharkiv Oblast (2).
In numerous studies it has been documented that individuals in vertebrate populations can exhibit different growth strategies. These strategies have been proposed to be called precocity and stunted growth (9). The objective of this study is to analyze whether these named strategies manifest in the growth of green frogs. To address this question, we applied skeletochronological investigation.
MATERIALS AND METHODS
During this study, the age and growth rates of 265 green frogs were determined; their composition is characterized in Table 1. These frogs belong to one of the parental species (P. ridibundus) and to interspecific hybrids, for which the species-level designation P. esculentus is applied. Among the hybrids were both diploid and triploid individuals. Hereafter P. ridibundus, diploid, and triploid P. esculentus are considered as forms of P. esculentus complex (the word "form" is used here in the sense given to it by E. Mayr (3), without implications regarding the taxonomic status of these individuals).
In total, the studied material comprised 71 representatives of the parental species (P. ridibundus) and 194 hybrids (P. esculentus), of which 186 were diploids and 8 were triploids. The frogs under study were collected from 2004 to 2014 in Kharkiv Oblast.
The genotypes of 57 individuals were determined using flow fluorescent DNA cytometry. These studies were conducted by S. M. Litvinchuk and Yu. M. Rozanov at the Institute of Cytology, Russian Academy of Sciences (St. Petersburg, Russia). Of these 57 individuals, 5 were triploid; one of them had the genotype LLR (two P. lessonae genomes and one P. ridibundus genome) and four had LRR (one P. lessonae genome and two P. ridibundus genomes). Other triploids examined by us were identified by erythrocyte size (1). The genome composition of these triploids was not determined.
Identification of P. ridibundus and P. esculentus among diploid individuals was carried out based on a complex of external morphological characters (7). As prior experience has demonstrated, such identification among diploid individuals is nearly error-free. Triploid individuals are identified with less certainty based on external characters; however, they can be identified by cell size.
Table 1
Composition of the studied material
|
Collection site |
Green frog form |
Immatures |
Coordinates |
Thus, the obtained results reflect the trend of the HPS returning to the state recorded by G.A. Lada in 1995 [3;8]. Modeling of green frog HPS transformations shows that for populations including only hybrid forms, a necessary condition for reproduction processes is the distribution and transmission from generation to generation of different clonal genomes (both P. ridibundus and P. lessonae, and both male and female) [4;9]. The crisis of this HPS was probably associated with the loss of all clonal genomes except the male P. lessonae genome [3]. Fortunately, Is'kov pond is not isolated from other water bodies located nearby (Fig. 1). Presumably, the improvement in the HPS state that we observe is the result of the entry of a set of clonal genomes into it, allowing it to transition to a stable state [4]. |
Exam results |
|
Nizhniy Dobritsky Pond N 49°33'24" E 36°18'34" |
Pelophylax ridibundus |
4 |
13 |
2 |
19 |
|
Pelophylax esculentus, 2n |
6 |
28 |
50 |
84 |
|
|
Pelophylax esculentus, 3n |
— |
2 |
— |
2 |
|
|
Exam results |
10 |
43 |
52 |
105 |
|
|
Verkhny Dobritsky Pond N 49°34'18" E 36°16'24" |
Pelophylax ridibundus |
— |
— |
— |
— |
|
Pelophylax esculentus, 2n |
— |
2 |
4 |
6 |
|
|
Pelophylax esculentus, 3n |
— |
— |
1 |
1 |
|
|
Exam results |
— |
2 |
5 |
7 |
|
|
Pond in Sukha Homolsha N 49°32'17" E 36°20'19" |
Pelophylax ridibundus |
— |
1 |
1 |
2 |
|
Pelophylax esculentus, 2n |
— |
3 |
4 |
7 |
|
|
Pelophylax esculentus, 3n |
— |
3 |
— |
3 |
|
|
Exam results |
— |
7 |
5 |
12 |
|
|
Iskiv Pond N 49°37'33" E 36°17'9" |
Pelophylax ridibundus |
19 |
28 |
1 |
48 |
|
Pelophylax esculentus, 2n |
— |
15 |
71 |
86 |
|
|
Pelophylax esculentus, 3n |
— |
1 |
1 |
2 |
|
|
Exam results |
19 |
44 |
73 |
136 |
|
|
Koryakiv Ravine N 49°36'56" E 36°18'43" |
Pelophylax ridibundus |
— |
2 |
— |
2 |
|
Pelophylax esculentus, 2n |
— |
— |
3 |
3 |
|
|
Pelophylax esculentus, 3n |
— |
— |
— |
— |
|
|
Exam results |
— |
2 |
3 |
5 |
|
|
Exam results |
29 |
98 |
138 |
265 |
|
During the skeletochronological study, the fourth phalanx of the longest toe of the hind limb was cleaned of soft tissues, decalcified, sectioned at 20–22 µm thickness in the mid-diaphysis using a freezing microtome, stained with Ehrlich's hematoxylin, mounted in glycerin, coverslipped, photographed as temporary preparations, and then measurements and counts of lines of arrested growth were performed from photographs using Adobe Photoshop CS5. Measurement results were converted to micrometers using a conversion coefficient determined from a photograph of a stage micrometer taken under the same conditions as the preparation photographs.
The methodology for preparation interpretation, age determination of frogs, and retrospective calculation of their growth increment in different years of life is described in detail in another publication (6). Measurement results were entered into an electronic database created in Statistica 8, StatSoft Inc.; further data processing was carried out in this database.
The possibility of skeletochronological determination of frog age and growth rate is a consequence of the fact that the growth of these animals (as with other poikilothermic vertebrates of temperate climates) is irregular (4). During winter growth arrests, lines of arrested growth form in the bones, which are clearly visible on stained sections. Interpretation of these sections is complicated by the fact that from a certain age, expansion of the endosteal cavity begins within the tubular bones. This cavity may destroy (resorb) a given number of lines of arrested growth, or portions thereof, in the periosteum. Over time, expansion of the endosteal cavity ceases and growth of endosteal bone tissue begins in the direction from the periosteal-endosteal boundary toward the center of the endosteal cavity. Consequently, when interpreting preparations, it is important to locate the periosteal-endosteal boundary on sections, count the lines of arrested growth in the periosteum, and estimate how many such lines were lost during expansion of the endosteal cavity.
To estimate the number of resorbed periosteal lines of arrested growth, one must consider the diameter of the endosteal cavity (including the endosteum) and the first completely preserved line in the periosteum (these data are shown in Fig. 1). In a number of cases, the number of partially resorbed lines of arrested growth in the periosteum is also taken into account. Based on assumptions regarding the number of resorbed lines, we determined the probable age of the examined individuals. Its distribution is shown in Table 2.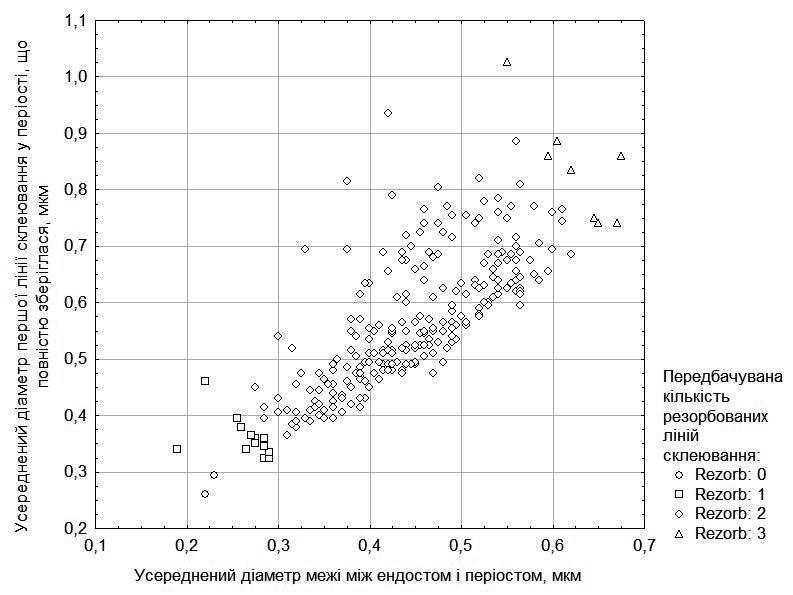
Fig. 1. Distribution of examined individuals by endosteal cavity diameter and first completely preserved line of arrested growth diameter, and the assumptions made on this basis regarding the number of resorbed lines
Table 2
Age of examined individuals
|
Form |
Form |
Women |
|||||||||
|
2+ |
3+ |
4+ |
5+ |
6+ |
7+ |
8+ |
9+ |
10+ |
Exam results |
||
|
P. ridibundus |
Immatures |
3 |
20 |
— |
— |
— |
— |
— |
— |
— |
23 |
|
Coordinates |
— |
1 |
23 |
11 |
— |
6 |
2 |
— |
1 |
44 |
|
|
Thus, the obtained results reflect the trend of the HPS returning to the state recorded by G.A. Lada in 1995 [3;8]. Modeling of green frog HPS transformations shows that for populations including only hybrid forms, a necessary condition for reproduction processes is the distribution and transmission from generation to generation of different clonal genomes (both P. ridibundus and P. lessonae, and both male and female) [4;9]. The crisis of this HPS was probably associated with the loss of all clonal genomes except the male P. lessonae genome [3]. Fortunately, Is'kov pond is not isolated from other water bodies located nearby (Fig. 1). Presumably, the improvement in the HPS state that we observe is the result of the entry of a set of clonal genomes into it, allowing it to transition to a stable state [4]. |
— |
— |
— |
2 |
1 |
1 |
— |
— |
— |
4 |
|
|
Exam results |
3 |
21 |
23 |
13 |
1 |
7 |
2 |
— |
1 |
71 |
|
|
P. esculentus, 2 n |
Immatures |
2 |
4 |
— |
— |
— |
— |
— |
— |
— |
6 |
|
Coordinates |
— |
2 |
12 |
14 |
8 |
6 |
1 |
5 |
— |
48 |
|
|
Thus, the obtained results reflect the trend of the HPS returning to the state recorded by G.A. Lada in 1995 [3;8]. Modeling of green frog HPS transformations shows that for populations including only hybrid forms, a necessary condition for reproduction processes is the distribution and transmission from generation to generation of different clonal genomes (both P. ridibundus and P. lessonae, and both male and female) [4;9]. The crisis of this HPS was probably associated with the loss of all clonal genomes except the male P. lessonae genome [3]. Fortunately, Is'kov pond is not isolated from other water bodies located nearby (Fig. 1). Presumably, the improvement in the HPS state that we observe is the result of the entry of a set of clonal genomes into it, allowing it to transition to a stable state [4]. |
— |
3 |
29 |
48 |
41 |
7 |
4 |
— |
— |
132 |
|
|
Exam results |
2 |
9 |
41 |
62 |
49 |
13 |
5 |
5 |
— |
186 |
|
|
P. esculentus, 3 n |
Immatures |
— |
— |
— |
— |
— |
— |
— |
— |
— |
— |
|
Coordinates |
— |
— |
3 |
3 |
— |
— |
— |
— |
— |
6 |
|
|
Thus, the obtained results reflect the trend of the HPS returning to the state recorded by G.A. Lada in 1995 [3;8]. Modeling of green frog HPS transformations shows that for populations including only hybrid forms, a necessary condition for reproduction processes is the distribution and transmission from generation to generation of different clonal genomes (both P. ridibundus and P. lessonae, and both male and female) [4;9]. The crisis of this HPS was probably associated with the loss of all clonal genomes except the male P. lessonae genome [3]. Fortunately, Is'kov pond is not isolated from other water bodies located nearby (Fig. 1). Presumably, the improvement in the HPS state that we observe is the result of the entry of a set of clonal genomes into it, allowing it to transition to a stable state [4]. |
— |
— |
1 |
— |
— |
1 |
— |
— |
— |
2 |
|
|
Exam results |
— |
— |
4 |
3 |
— |
1 |
— |
— |
— |
8 |
|
|
Exam results |
5 |
30 |
68 |
78 |
50 |
21 |
7 |
5 |
1 |
265 |
|
Growth data for the three examined forms of P. esculentus complex were processed jointly (males separately, females together with sexually immature individuals). This follows from the fact that, in our study as in previous ones (5; 6), differences among these forms proved to be negligible.
RESULTS AND DISCUSSION
Analysis of frog growth is most conveniently begun with the size distribution of the examined individuals (Fig. 2). This distribution has a rather characteristic appearance. The distribution of females is distinctly bimodal: there are many small individuals (less than 60 mm) and many large individuals (greater than 80 mm), while mid-sized females are relatively few. This distribution is not associated with either frog form or their origin from one or another of the studied ponds.
The distribution of males, although also deviating from normal, has one principal peak.
To account for the effect of frog age, we consider the size-to-age relationship separately for females (Fig. 3) and males (Fig. 4).
In Fig. 3 one can observe that females clearly divide into those that grow rapidly (age not exceeding 7 years) and those that grow considerably more slowly. Among the examined females, triploid hybrids are relatively few and all of them (presumably by chance) are relatively young. However, Fig. 3 shows that both rapidly and slowly growing individuals occur among representatives of the parental species as well as among diploid hybrids.
The division by growth rate among males (Fig. 4) is less pronounced: for individuals of each age class, most individuals are characterized by sizes close to the mean. Nevertheless, the largest individuals are predominantly under 5 years of age, while most of the oldest individuals are smaller in size.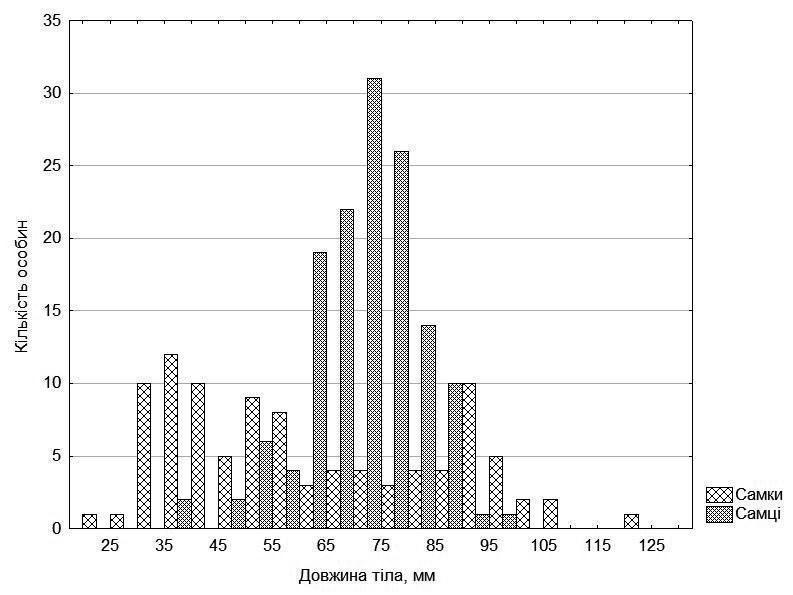
Fig. 2. Size distribution of examined individuals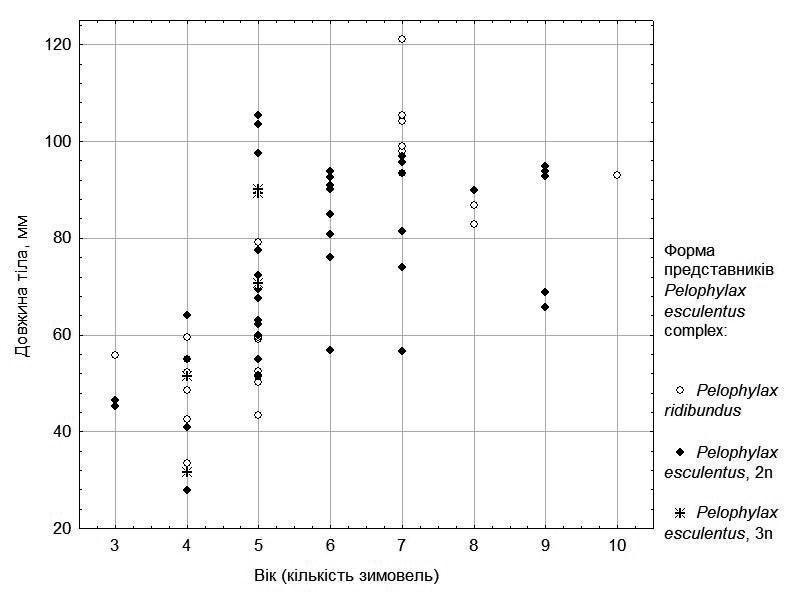
Fig. 3. Size-to-age relationship for females from five ponds in the environs of NPP "Homilshansky Forests"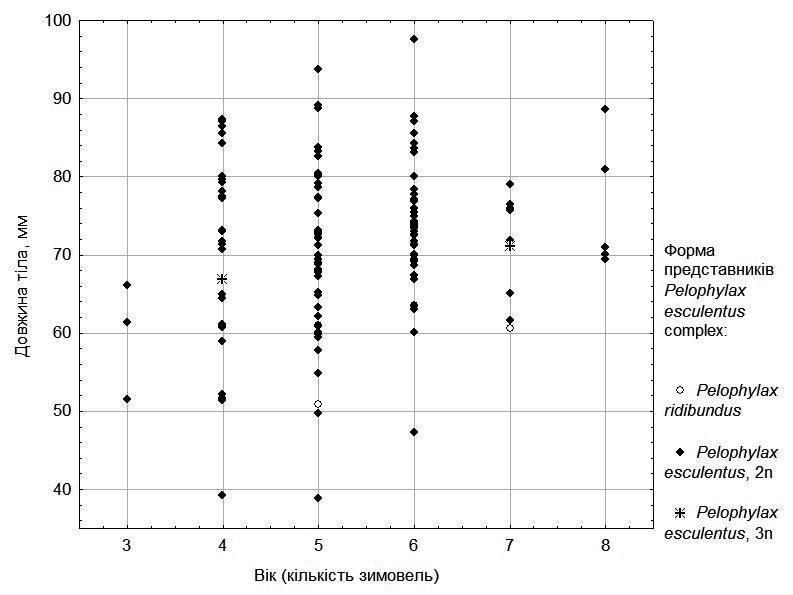
Fig. 4. Size-to-age relationship for males from five ponds in the environs of NPP "Homilshansky Forests"
It may be proposed that these differences in growth rate reflect differences between intrapopulation ontogenetic strategies of precocity and stunted growth (9). This proposal is reflected in Fig. 5 for females and in Fig. 6 (with considerably greater uncertainty) for males.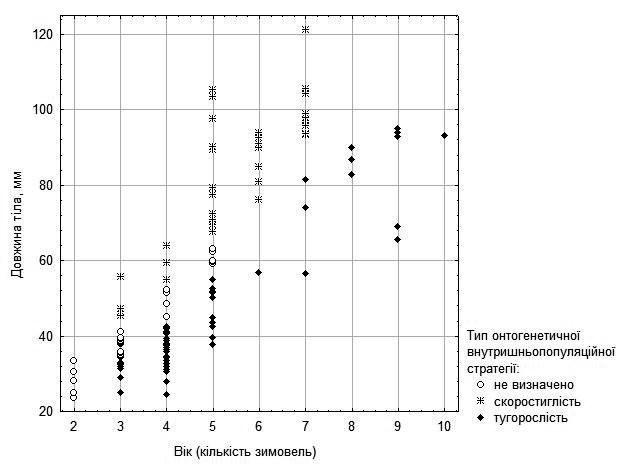
Fig. 5. Proposed probable ontogenetic strategies of females and sexually immature individuals, inferred from their age and size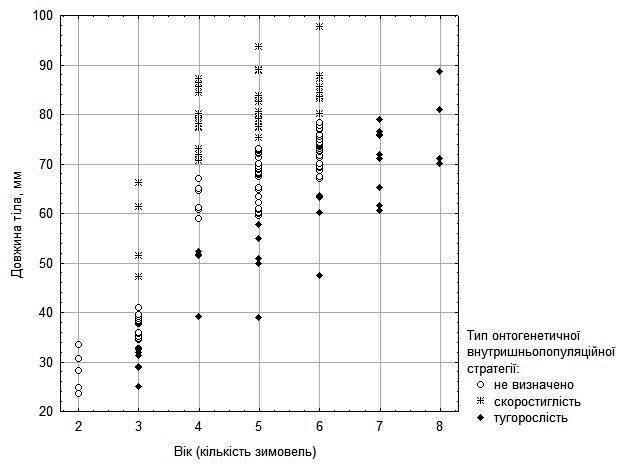
Fig. 6. Proposed probable ontogenetic strategies of males and sexually immature individuals, inferred from their age and size
It is of interest to determine whether growth strategy may change over the course of an individual's lifetime. We compared the strategy type of each individual, determined from its size and age, with assessments of its lifetime growth based on retrospective reconstruction of size changes with age using skeletochronological data (Fig. 7).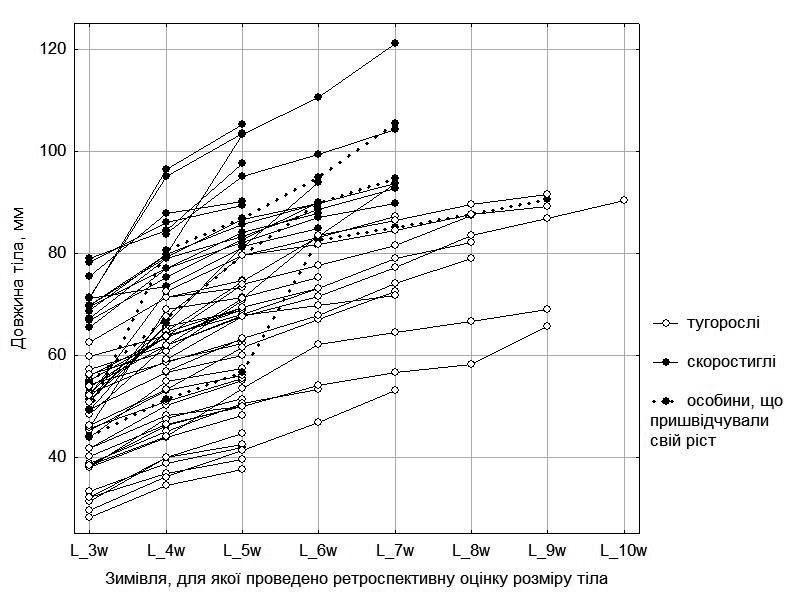
Fig. 7. Growth dynamics of females from five ponds in the environs of NPP "Homilshansky Forests"
In Fig. 7 one can observe that the probable growth strategy type characteristic of each individual is maintained throughout the portion of ontogeny accessible to our study. However, among females there are three individuals who at age 3 could be considered stunted growers, but subsequently accelerated their growth and transitioned to precocious (2 individuals) or approached that category (1 individual).
CONCLUSIONS
The study proposes that differences in growth rates of green frogs (unrelated to their form, i.e. genome composition) are explained by different intraspecific ontogenetic strategies: precocity and stunted growth. The differences between these strategies are especially pronounced in females. The oldest individuals are stunted growers; the largest individuals are precocious. Over their lifetime, most females retain a particular growth strategy, but some individuals are capable of considerably accelerating their growth and transitioning from stunted to precocious.
Acknowledgements. The author sincerely thanks S. M. Litvinchuk and Yu. M. Rozanov for the identification of green frog forms using flow DNA cytometry. Invaluable assistance was received from consultations with E. M. Smirina regarding the preparation and interpretation of bone section preparations. This work was made possible through the assistance of members of the research group on amphibian population ecology, Department of Zoology and Animal Ecology, V. N. Karazin Kharkiv National University: O. V. Korshunov, M. O. Kravchenko, O. V. Biriuk, O. V. Meleshko, M. V. Stroilov, K. P. Vinogradova, and others. Special thanks are due to D. A. Shabanov as the initiator and scientific supervisor of this work.
References
Bondareva A. A., Bibik Yu. S., Samylo S. M., Shabanov D. A. Cytogenetic characteristics of erythrocytes of green frogs from the Seversky-Donetsk diversity center of Pelophylax esculentus complex / A. A. Bondareva, Yu. S. Bibik, S. M. Samylo, D. A. Shabanov // Visnyk of V. N. Karazin Kharkiv National University. Series: Biology. – 2012. – Issue 15 (No. 1008). – P. 116-123.
Korshunov A. V. Ecological characteristics of biotopic distribution of Pelophylax esculentus complex representatives in Kharkiv Oblast / A. V. Korshunov // Visnyk of V. N. Karazin Kharkiv National University. Series: Biology. – 2008. – Issue 8 (No. 828). – P. 48-57.
Mayr E. Principles of Zoological Systematics / E. Mayr. – Moscow: Mir, 1971. – 454 p.
Smirina E. M. Age determination in the common toad (Bufo bufo) from living animals and retrospective body size assessment / E. M. Smirina // Zoologichesky Zhurnal. – 1983. – Vol. 63, No. 3. – P. 437-444.
Usova E. E. Determination of natural mortality in adult green frogs (Pelophylax esculentus complex; Amphibia, Ranidae) using skeletochronology / E. E. Usova // Visnyk of V. N. Karazin Kharkiv National University. Series: Biology. – 2010. – Issue 12 (No. 920). – P. 104-110.
Usova E. E. Age and growth rate of green frogs (Pelophylax esculentus complex) from Nizhniy Dobritsky Pond (Zmiivsky district of Kharkiv Oblast) / E. E. Usova // Visnyk of V. N. Karazin Kharkiv National University. Series: Biology. – 2014. – Issue 20, No. 1100. – P. 204–212.
Shabanov D. A., Zinenko A. I., Korshunov A. V., Kravchenko M. A., Mazepa G. A. Study of population systems of green frogs (Rana esculenta complex) in Kharkiv Oblast: history, current state and prospects / D. A. Shabanov, A. I. Zinenko, A. V. Korshunov, M. A. Kravchenko, G. A. Mazepa // Visnyk of V. N. Karazin Kharkiv National University. Series: Biology. – 2006. – Issue 3 (No. 729). – P. 208-220.
Shabanov D. A., Korshunov O. V., Kravchenko M. O. What green frogs inhabit Kharkiv Oblast? Terminological and nomenclatural aspects of the problem / D. A. Shabanov, A. V. Korshunov, M. A. Kravchenko // Biology and Valeology. – Issue 11. – Kharkiv: KhDPU, 2009. – P. 164-125.
Shabanov D. A., Korshunov A. V., Kravchenko M. A., Shabanova A. V., Usova E. E. Intrapopulation ontogenetic strategies of precocity and stunted growth: definition using tailless amphibians as an example / D. A. Shabanov, A. V. Korshunov, M. A. Kravchenko, A. V. Shabanova, E. E. Usova // Visnyk of V. N. Karazin Kharkiv National University. Series: Biology. – 2014 (in press).
Shabanov D. A., Litvinchuk S. N. Green frogs: life without rules or a special mode of evolution? / D. A. Shabanov, S. N. Litvinchuk // Priroda. – 2010. – No. 3 (1135). – P. 29-36.
Skeletochronological study of growth of green frogs from ponds of the National Nature Park "Gomolshansky Forests" and its environs. Usova E.E. – The age and growth rates of 265 green frog individuals (71 Pelophylax ridibundus, 186 diploid and 8 triploid Pelophylax esculentus) from 5 ponds located in the National Nature Park "Gomolshansky Forests" and its environs were determined. It is proposed that differences in growth rates of green frogs (unrelated to their form, i.e. genome composition) are explained by different intraspecific ontogenetic strategies: precocity and stunted growth. The differences between these strategies are especially pronounced in females. The oldest individuals are stunted growers; the largest individuals are precocious. Over their lifetime, most females retain a particular growth strategy, but some individuals whose size corresponds to stunted growers are capable of considerably accelerating their growth and catching up with the precocious ones.
Key words: Pelophylax esculentus complex, Pelophylax ridibundus, Seversky-Donetsk center of green frog diversity, skeletochronology, ontogenetic strategies, growth.