
Студентські роботи після практики II курсу — 2010
Afoničeva Ya.V., Bondareva A.A., Balanyuk E.V., Bіbik Yu.S. Vивчення відповідностей між стадіями розвитку задніх кінцівок і ротових апаратів у процесі нормального розвитку головастиків зелених жаб. Bелкіна I.O., Bryleva O.P. Різниця у пропорціях і розмірах головастиків зелених жаб, проходя...
Студентські роботи після практики II курсу – 2008
Студентські роботи після практики II курсу – 2009
Студентські роботи після практики II курсу – 2011
Студентські роботи після практики II курсу – 2012 (I частина)
Студентські роботи після практики II курсу – 2012 (II частина)
Студентські роботи після практики II курсу – 2013
Студентські роботи після практики II курсу – 2014
Студентські роботи після практики II курсу – 2015
Студентські роботи після практики II курсу – 2016
Студентські роботи після практики II курсу – 2017
ВИВЧЕННЯ ВІДПОВІДНОСТІ МІЖ СТАДІЯМИ РОЗВИТКУ ЗАДНІХ КІНЦЕВ ТА РОТОВИХ АПАРАТІВ ПІД ЧАС НОРМАЛЬНОГО РОЗВИТКУ ГОЛОВАСТИКІВ ЗЕЛЕНИХ ЖАБ
Афонічева Я. В., Бондарєва А. А., Баланюк Є. В., Бібік Ю. С.
Харківський національний університет імені В. Н. Каразина, біологічний факультет, пл. Свободи, 4, м. Харків, Україна
У ході нормального розвитку головастиків зелених жаб (Pelophylax esculentus complex) одним і тим самим стадіям розвитку задніх кінцівок можуть відповідати різні типи ротових апаратів. Це можна пояснити за допомогою двох альтернативних гіпотез: (1) певній стадії розвитку задніх кінцівок відповідає кілька «підстадій», що відрізняються будовою ротових апаратів; (2) кожен головастик має характерний для нього тип ротового апарату, з яким він проходить усі стадії розвитку кінцівки. Ми провели експеримент для перевірки цих гіпотез.
Для експерименту використали головастиків, зловлених на лівому березі р. Сіверський Донеч нижче за течією м. Змієва Харківської обл. З зловлених головастиків були сформовані пари з однаковим розвитком задніх кінцівок (з 2‑ї по 7‑му стадії; див. тези Т. Виноградської тощо в цьому збірнику) і однаковими ротовими апаратами (типи A і D; див. тези Ю. Бібік в цьому збірнику). Ми утримували 15 пар головастиків у ємкостях по 330 мл води протягом 6 днів. Раз на добу воду міняли, а головастиків годували надмірною кількістю сухого акваріумного корму. Першій з перевірюваних гіпотез мав би відповідати результат, при якому типи ротових апаратів головастиків змінювалися б частіше (у крайньому випадку – з тією ж швидкістю), ніж стадії розвитку кінцівок. Другій гіпотезі відповідало б зміна стадій кінцівок у експериментальних при незмінності їх ротових апаратів.
На початку експерименту тип A ротового апарату був зафіксований лише у головастиків, що знаходилися на відносно ранніх стадіях (у всіх 4‑х головастиках на 2‑й стадії розвитку задньої кінцівки і у 2‑х з 16 головастиків на 4‑й стадії розвитку кінцівок). За час експерименту ці головастики змінили тип A на D (за винятком одного загиблого і одного травмованого). Усі 24 головастика з типом ротового апарату D, що перебували на стадіях від 3 до 7, зберегли свої ротові апарати незмінними. Розвиток задніх кінцівок у більшості головастиків просунувся на одну‑дві стадії. Найактивніше розвивалися кінцівки, що були на стадії 4 (у деяких особин розвиток просунувся на три‑чотири стадії), хоча у головастиків, які змінили ротовий апарат, задня кінцівка не змінилася.
Таким чином, ми зафіксували два процеси: (1) розвиток задніх кінцівок від початкових стадій до наступних, що відбувається з різною швидкістю; (2) перестройку ротового апарату від типу A до D (що відбувається на різних стадіях розвитку кінцівок у різних особин). Существенно, що зміна типу ротового апарату відбувається в одному напрямку і лише на відносно ранніх стадіях розвитку кінцівки. Цей результат не відповідає жодній з тестованих гіпотез, хоча в цілому ближче до другої. Ймовірно, характерний для кожного головастика тип ротового апарату формується не одразу, а проходить кілька етапів на ранніх стадіях розвитку.
Summary. It has been investigated hind limbs and oral apparatus development character in the course of normal ontogenesis of water frogs tadpoles to examine the fits between certain stages of hind limbs development and the types of oral apparatus. According to the results of our experiment it has been suggested that the kind of oral apparatus which is typical for each tadpole is not formed at once but it passes several phases in the early stages of development.
Описаний експеримент виконаний у ході УІРС на навчальній практиці з зоології хребетних. Автори вдячні доц. Шабанову Д. А. за керівництво даною роботою.
Афонічева Я. В., Бондарєва A. A., Баланюк Є. В., Бібік Ю. С. Вивчення відповідностей між стадіями розвитку задніх кінцівок і ротових апаратів у ході нормального розвитку головастиків зелених жаб.// "Біологія : від молекули до біосфери". Матеріали V Міжнародної конференції молодих науковців. – Харків: Оперативна поліграфія, 2010. – с. 337‑338.
РОЗЛІКИ В ПРОПОРЦІЯХ ТА РОЗМІРАХ ГОЛОВАСТИКІВ ЗЕЛЕНИХ ЖАБ, ЩО РОЗВИВАЮТЬСЯ У РІЗНИЙ ЧАС
Білкіна І. О., Брілєва О. П.
Харківський національний університет імені В. Н. Каразина, біологічний факультет, пл. Свободи 4, м. Харків, Україна
Європейські зелені жаби (Pelophylax esculentus complex) – цікава для вивчення група тварин, у якій спостерігаються міжвидова гибридизація, поліплоїдія та напівклоний спадок. Для вивчення факторів, які впливають на склад популяційних систем зелених жаб, необхідно дослідити розвиток їх головастиків у природних умовах. Ми вивчили 3 вибірки головастиків, зібрані на лівому березі р. Сіверський Донеч нижче за течією м. Змієва: вибірка № 1 (24.06.2010) складала 130 головастиків, вибірка № 2 (01.07.2010) – 232 головастика, вибірка № 3 (10.07.2010) – 139 головастиків.
Для вимірювання довжини тіла і довжини тулуба використали штангенциркуль (вимірювання проводились з точністю до 0,1 мм); для вимірювання ваги – електронні ваги (з точністю до 0,001 г). Стадії розвитку головастиків визначали за задніми кінцівками за допомогою стереоскопічного мікроскопа (див. тези Виноградської тощо в цьому збірнику).
Дані обробляли за допомогою програми Statistica for Windows 6.5
Ми зафіксували, що головастики, які розвиваються пізніше, мають відносно довший тулуб (рис. 1). Дисперсійний аналіз (у якому розглядається вплив дати збору і стадії розвитку на відносну довжину тіла) показав, що різниця між вибірками високо значуща (p < 0,000001)).
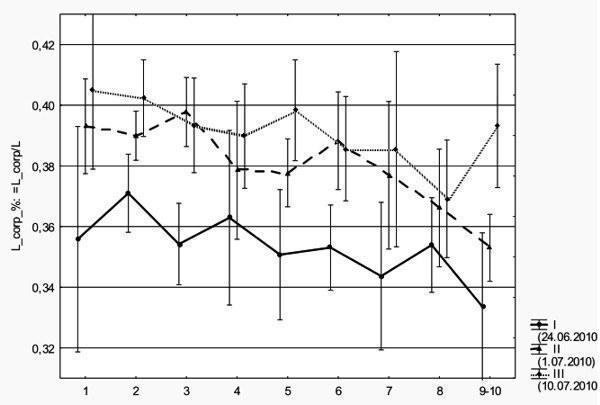
Рис. 1. Відносна довжина тулуба (відношення довжини тулуба до загальної довжини тіла) головастиків трьох вибірок, зібраних у різний час. По осі абсцис відкладені стадії розвитку задніх кінцівок
Водночас, головастики на однакових стадіях розвитку з різних вибірок значно менш відрізняються за загальною довжиною тіла і практично не відрізняються за вагою. Зафіксований результат міг би бути пов’язаний із систематичною помилкою при вимірюванні довжини тулуба в деяких з вивчених вибірок, проте автори вважають її малоймовірною і розраховують на переперевірку їх даних у наступному польовому сезоні. Отже, головастики, що розвиваються у різний час, відрізняються за пропорціями, причому подовження тулуба компенсується їх «стройністю», так що їх вага змінюється незначно.
Summary. At comparison of tadpoles of the green frogs caught at various times (on July, 24th, on July, 1st and on July, 10th 2010) differences in their proportions are registered. At tadpoles which educe later, the attitude of length of a trunk to body total length significantly increases.
Описані результати отримані в ході навчально‑дослідної роботи на навчальній практиці з зоології хребетних. Автори висловлюють подяку Ю. С. Бібік за допомогу у виконанні роботи та доц. Д. А. Шабанову за наукове керівництво.
Білкіна І. О., Брілєва О. П. Різниця в пропорціях і розмірах головастиків зелених жаб, що проходять розвиток у різний час.// "Біологія : від молекули до біосфери". Матеріали V Міжнародної конференції молодих науковців. – Харків: Оперативна поліграфія, 2010. – с. 341‑342.
ЯКІ ФАКТОРИ ВИЗНАЧАЮТЬ ТИП РОТОВОГО АПАРАТУ ГОЛОВАСТИКІВ ЗЕЛЕНИХ ЖАБ?
Бібік Ю. С.
Харківський національний університет імені В. Н. Каразина, біологічний факультет, пл. Свободи, 4, м. Харків, Україна. e‑mail: yulia.bibik@gmail.com
Гібридогенний комплекс європейських зелених жаб (Pelophylax esculentus complex) представлений двома батьківськими видами і кількома формами напівклоних гибридів. Особливості розвитку головастиків різних форм зелених жаб досі майже не описані. Ми вивчали головастиків з 7 вибірок. Вибірки № 1 і № 2 зібрані на лівому березі р. Сіверський Донеч нижче м. Змієва 24 червня і 1 липня 2010 р. відповідно. Вибірки № 3 і № 4 вирощені з кладок ікри, зібраних 6 травня 2010 р. у поймі Сіверського Донця поблизу біостанції ХНУ, а вибірка № 5 отримана від пари жаб (самка Pelophylax ridibundus, самець P. esculentus), зловлених там же. Ймовірно, у склад вибірок 1‑5 входили як P. ridibundus, так і P. esculentus. Вибірки № 6 і № 7 зібрані 18 червня 2010 р. у Донецькій області, у водоймах, населений виключно P. ridibundus.
Фіксованих у формаліні головастиків розглядали під стереоскопічним мікроскопом, визначали тип будови ротового апарату (рис. 1) і стадію розвитку задніх кінцівок (див. роботу Виноградської тощо в цьому ж збірнику). Результати визначення типів ротових апаратів наведені в табл. 1.
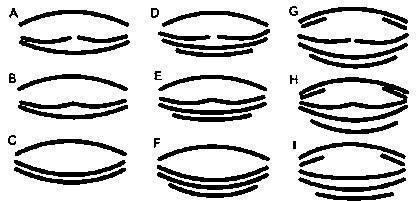
Рис. 1. Класифікація типів ротових апаратів головастиків зелених жаб (умовно показано взаєморозташування скоблячих рогових пластин)
Таблиця 1. Різноманітність типів ротових апаратів у вивчених вибірках головастиків
Вибірки | К‑к | Стадії розвитку задніх кінцівок | Типи ротових апаратів
--- | --- | --- | ---
| | | A | B | C | D | E | F | G | H | I
1 і 2 | 352 | 1–10 | 0 | 0 | 0 | 31 | 0 | 16 | 242 | 5 | 58
3–5 | 419 | 1–8 | 81 | 18 | 30 | 216 | 12 | 14 | 48 | 0 | 0
6 і 7 | 96 | 2–8, 10 | 0 | 0 | 0 | 0 | 0 | 0 | 82 | 0 | 14
Вибірки 3‑5, що розвивалися в однакових умовах, але ймовірно різняться за генотипом, значно відрізнялися за співвідношенням семи зафіксованих у них типів ротових апаратів (при порівнянні за критерієм χ² p < 0,00001). Схоже, на різноманітність ротових апаратів впливають генотипічні особливості жаб. У вибірках 1 і 2, отриманих з природного середовища, не виявлено типи A, B, C і E, але виявлено типи H і I. Нарешті, у вибірках 6 і 7, що складалися лише з головастиків P. ridibundus, зафіксовано лише 2 типи ротових апаратів: G і I. Найменше різноманіття типів ротових апаратів характерне для вибірок одного виду з природного середовища, а найбільше — для змішаних вибірок, що розвивалися в штучних умовах. Отже, на різноманіття ротових апаратів головастиків впливає і їх вид, і генотип, і умови їх розвитку, але жоден з цих факторів не може повністю пояснити спостережувану змінність їх ротових апаратів.
Summary. The classification of types of scrape oral apparatuses of water frogs tadpoles (Pelophylax esculentus complex) has been developed. The variety of oral apparatuses types has been described in a few selections of tadpoles. Species, genotype and conditions of tadpoles development influence on the oral apparatus structure. The complete explanation of observed picture of oral apparatus mutability was not successful.
Автор дякує Т. Виноградській, Н. Толокнеєвій, Є. Комаровій і В. Хлопоніні за допомогу у виконанні роботи, а доц. Д. А. Шабанову — за наукове керівництво.
Бібік Ю. С. Які фактори визначають тип ротового апарату головастиків зелених жаб? //"Біологія : від молекули до біосфери". Матеріали V Міжнародної конференції молодих науковців. – Харків: Оперативна поліграфія, 2010. – с. 339‑340.
ВПЛИВ ЕФЕКТУ ГРУПИ НА РОЗВИТОК ГОЛОВАСТИКІВ ЗЕЛЕНИХ ЖАБ
Бібік Ю. С., Коваленко М. С., Кучкова А. Г.
Харківський національний університет імені В. Н. Каразина, біологічний факультет, пл. Свободи, 4, м. Харків, Україна. e‑mail: yulia.bibik@gmail.com
Завдання даної роботи — встановити, чи впливає перебування у складі різновікового гурту на розвиток ротових апаратів і кінцівок головастиків представників Pelophylax esculentus complex. Як відомо (Міна, Клевезаль, 1976), групове утримання водних тварин змінює їх онтогенез у порівнянні з одиничним.
Ми використали головастиків з річки Сіверський Донеч, зібраних на лівому березі нижче м. Змієва 24.06.2010 р. Розглядаючи живих головастиків під стереоскопічним мікроскопом, ми обрали дві групи головастиків з однаковою будовою задніх кінцівок. 24 «малі» головастика були на 3‑й стадії розвитку кінцівок (див. тези Виноградської тощо), 12 великих — на 7‑й. В обох групах були представлені головастики типів ротових апаратів (див. тези Бібік) G, D, H і I. Головастиків зважували і розміщували у ємкостях (півлітрові банки) з 0,3 л річкової води. У 6 ємкостей поклали по 1 «малій» головастик, у 6 — по 1 «великій», і ще в 6 — по 3 «малі» і 1 «велику». Раз на добу воду міняли і годували головастиків надмірною кількістю сухого акваріумного корму. Через 10 днів провели повторні вимірювання маси, визначили стадії розвитку кінцівок і типи ротових апаратів.
У «малих» головастиків‑одинаків стадії розвитку кінцівок змінилися з 3‑ї до 6‑ї, а в групах — з 3‑ї до 4‑ї. У «великих» головастиків незалежно від умов утримання відбулося зміна з 7‑ї до 9‑ї стадії. Групове утримання значно (p = 0,004) зменшило відносне прирост маси в обох групах, практично однаково для «великих» і «малих». Таким чином, групове утримання уповільнило ріст (за оцінкою збільшення маси) в обох групах, а розвиток (за оцінкою стану кінцівок) — лише у «малих».
Тип ротового апарату у «малих» головастиків змінювався значно (при порівнянні за Манном‑Уїтні p = 0,005) частіше, ніж у «великих». Групове утримання не вплинуло на частоту зміни типів ротових апаратів у «великих» головастиків і незначно (p = 0,32) знизило її у «малих». У «малих» головастиків зафіксовано такі комбінації початкових і кінцевих типів ротових апаратів: D→D (13 випадків); D→E (1); D→G (3); G→G (2); G→H (2); H→H (1); I→G (2). Для «великих» головастиків зафіксовано комбінації G→G (2 випадки); G→H (5); G→редукція ротового апарату (5 випадків). Остання група випадків нагадує події, що відбуваються під час метаморфози (коли скоблячий ротовий апарат редукується і розвивається ротовий апарат, призначений для захоплення здобичі), проте ці процеси відбуваються на пізніших стадіях розвитку.
Summary. Development of hind limbs and oral apparatus as well as mass growth have been studied in small and big tadpoles of water frogs(Pelophylax esculentus complex) maintained solitary or in groups (1 big and 3 small tadpoles in each group). Maintenance in groups inhibits growth both in small and big tadpoles, whereas hind limbs development is inhibited only in small tadpoles. Alteration of structure in various types of oral apparatus is also described.
Описаний експеримент виконаний як навчально‑дослідна робота на навчальній практиці II курсу. Автори дякують доц. Д. А. Шабанову за наукове керівництво даною роботою.
Бібік Ю. С., Коваленко М. С., Кучкова А. Г. Вплив ефекту групи на розвиток головастиків зелених жаб. // "Біологія : від молекули до біосфери". Матеріали V Міжнародної конференції молодих науковців. Харків: Оперативна поліграфія, 2010. – с. 338‑339.
РОСТ І РОЗВИТОК ГОЛОВАСТИКІВ ЗЕЛЕНИХ ЖАБ У ПРИРОДНОМУ СЕРЕДОВИЩІ
Виноградська Т. І., Толокнеєва А. А., Комарова Є. В., Хлопоніна В. В., Бібік Ю. С.
Харківський національний університет імені В. Н. Каразина; пл. Свободи, 4, м. Харків, Україна
Європейські зелені жаби (Pelophylax esculentus complex) — гібридогенний комплекс, що привертає увагу багатьох дослідників (Шабанов, Литвинчук, 2010). Для комплексного вивчення цієї групи тварин потрібно встановити, як проходить їх онтогенез, зокрема — личинковий розвиток. Після опису розвитку змішаних вибірок головастиків, взятих з природних ГПС, можна буде порівняти особливості личинкового розвитку окремих форм жаб.
Ми вивчили головастиків, зібраних на лівому березі Сіверського Донця нижче за течією м. Змієва в заростях водної рослинності на мілководді. Вибірка № 1, зібрана 24 червня 2010 р., складає 130 особин, а вибірка № 2, зібрана 1 липня 2010 р. — 232 особини.
Мірами росту головастиків були загальна довжина тіла з хвостом і довжина тулуба (які вимірювали штангенциркулем з точністю до 0,1 мм), а також вага (яку вимірювали електронними вагами з точністю до 0,001 г). Стадії розвитку визначали за станом задніх кінцівок. Виділяли такі стадії: 1) відсутність кінцівок; 2) зачатки задніх кінцівок мають вигляд круглого бугорка; 3) зачатки кінцівок починають згинатися; 4) дистальний кінець кінцівки приймає форму лопаточки; 5) зачатки 3‑х пальців; 6) зачатки 4‑х пальців; 7) зачатки 5‑ти пальців, починає розвиватися колінний суглоб; 8) сформований колінний суглоб; 9) сформовані колінний суглоб і суглоб між гомілкою та другорядною гомілкою, перепонки між 3‑ма пальцями; 10) повністю сформована міжпальцева перепонка. Дані обробляли за допомогою програми Statistica for Windows 6,5.
В цілому зареєстрований зв’язок довжини (L, мм) і ваги (W, г) головастиків може бути екстраполований рівнянням W = 0,26 – 0,27 × L + 0,098 × L. Значення довжини і ваги для окремих стадій наведено в табл. 1.
Табл. 1. Середні значення довжини і ваги головастиків на різних стадіях їх розвитку
Стадія | К‑к особин (вибірки № 1+№ 2) | Довжина (L), мм | Вага (W), г
--- | --- | --- | ---
1 | 4 + 24 | 2,02 | 0,107
2 | 37 + 74 | 2,74 | 0,265
3 | 23 + 22 | 3,24 | 0,408
4 | 5 + 12 | 3,84 | 0,657
5 | 7 + 29 | 3,88 | 0,659
6 | 22 + 20 | 4,22 | 1,000
7 | 19 + 8 | 4,76 | 1,261
8 | 11 + 17 | 5,30 | 1,698
9 | 1 + 11 | 5,52 | 1,788
10 | 1 + 15 | 6,42 | 2,597{"translated_text":"Як видно з табл. 1, вибірки відрізняються за відносною чисельністю головастиків, що перебувають на різних стадіях розвитку. При порівнянні зареєстрованого розподілу з теоретичним, побудованим на підставі припущення про відсутність зв’язку вибірки і стадії розвитку, критерій Пірсона χ²= 34,4, що при даній кількості ступенів свободи відповідає високозначимим різницям (p = 0,000075). Перевага у вибірці № 2 головастиків на пізніх стадіях, ймовірно, пов’язана з більш тривалим часом їх розвитку.\n\nSummary. Described stage of the development of the hind limbs water frog (Pelophylax esculentus complex). The studied polliwogs caught in the river Severskiy Donec in a neighborhood of Zmiev. Dependency of the weight (W) polliwog from their lengths of the body (L) is of the form of W = 0,26 - 0,27 L + 0,098 L 2.\nОписані результати отримані в ході навчально‑дослідної роботи на навчальній практиці з зоології хребетних. Автори висловлюють подяку доц. D. A. Shabanovu за керівництво даною роботою.\n\nVinogradska T. I., Toloknyeva A. A., Komarova E. V., Khloponina V. V., Bіbik Yu. S. Зростання і розвиток головастиків зелених жаб у природному середовищі // \"Біологія : від молекули до біосфери\". Матеріали V Міжнародної конференції молодих науковців. – Харків: Оперативна поліграфія, 2010. с. 344‑345.\nВИВЧЕННЯ ФЛУКТУЮЧОЇ АСИМЕТРІЇ ЗАБАРВЛЕННЯ У ЗЕЛЕНИХ ЖАБ\nKryshchalʹ O. A., Matasova E. V.\nХарківський національний університет імені В. N. Каразина, біологічний факультет, пл. Свободи, 4, м. Харків, Україна\nВизначення рівня флуктуючої асиметрії, FA (ненаправлених випадкових відхилень від біліатеральної симетрії) — один із способів оцінки стабільності онтогенезу. Розвиток у міжвидових гібридів зазвичай менш стабільний, ніж у батьківських видів. У околицях біостанції ХНУ в с. Гайдари (Зміївський р-н Харківської області) серед європейських зелених жаб живуть представники батьківського виду, Pelophylax ridibundus (озерні жаби) і геміклональні міжвидові гибриди, Pelophylax esculentus (їстівні жаби). У 2008 році FA їхньої забарвлення вивчали за загальноприйнятими ознаками — асиметричністю дорзомедіальної смуги, кількістю плям на спині та плям‑смуг на задніх кінцівках (Deriazhentsova et al., 2008). Як не дивно, FA гібридів у цьому дослідженні не перевищував FA батьківського виду. Ми застосували для вимірювання FA жаб новий метод, припускаючи, що він виявиться більш чутливим до відхилень від симетрії забарвлення.\nУ заплаві р. Сіверський Донець у околицях с. Гайдари було спіймано 186 зелених жаб. Їх визначали за зовнішніми ознаками: низький і косий п'ятковий бугор, відсутність жовтуватого відтінку на стегнах, темні резонатори у самців відповідає P. ridibundus; високий бугор, наявність жовтого відтінку на стегнах, світлі резонатори у самців відповідає P. esculentus. Ми визначили 107 особин (46 самок, 36 самців і 25 напівдорослих) як P. esculentus, 55 особин (31+12+12 відповідно) — як P. ridibundus, і 24 особи (12+6+6) залишилися невизначеними через нехарактерне поєднання їхніх ознак.\nСпинну сторону тулуба і кінцівок кожної жаби фотографували (для цього жабу давали схопитися передніми кінцівками за опору і розтягували її в довжину за пальці задніх кінцівок), а потім випускали у місця відлову. У програмі CorelDraw на кожну фотографію була накладена сітка, що розбиває її вздовж осі тіла жаби, від рила до п'яткових бугорів, на 20 зон. Для всіх зон, для яких це було можливо, визначали, чи симетрична в ній забарвлення (S), слабо асиметрична (H) або сильно асиметрична (A). Підраховували кількість зон кожного типу: NS, NH і NA. Результати обробляли в програмі Statistica. Асиметричність особи обчислювали як FA = (NA+ NH/2) / NS.\nОзнаки FA, що враховують знак відхилення від симетрії, мають нормальний розподіл зі середнім значенням 0. Наша міра FA приймає лише позитивні значення. Найбільш чисельними виявилися значення, близькі до 0; зі зростанням показника FA частота зустрічальності значень зменшувалась. Такий розподіл підтверджує, що використана міра характеризує FA і може застосовуватись для вивчення стабільності розвитку жаб, і, ймовірно, деяких інших тварин. P. esculentus виявилися значно (при порівнянні за Манном‑Уїтні p=0,031) асиметричнішими, ніж P. ridibundus. Медіана використаної міри FA для P. esculentus становить 0,4, для P. ridibundus — 0,33, а для невизначених особин — 0,39. Вплив статі та віку на FA забарвлення виявився незначним.\nSummary. For study fluctuating asymmetry (FA) tinge in water frogs, their photographed from back side. Photography was dividing along axis body on 20 zones. We counted the symmetrical, weak and hard asymmetrical zones. The tinge of hybrids form water frogsP. esculentus was found a more asymmetrical, then tinge of representatives by generation species water frogsP. ridibundus.\nРобота виконана як УІРС на навчальній практиці з зоології хребетних. Автори дякують доценту D. A. Shabanovu за ідею роботи та наукове керівництво нею.\n\nKryshchalʹ O. A., Matasova E. B. Вивчення флуктуючої асиметрії забарвлення у зелених жаб. // \"Біологія : від молекули до біосфери\". Матеріали V Міжнародної конференції молодих науковців. – Харків: Оперативна поліграфія. 2010. – с. 355‑356."}
{
"translated_text": "B\n\nC\n\nD\n\nE\n\nF\n\nG\n\nH\n\nI\n\n1 і 2\n\n352\n\n1–10\n\n0\n\n0\n\n0\n\n31\n\n0\n\n16\n\n242\n\n5\n\n58\n\n3–5\n\n419\n\n1–8\n\n81\n\n18\n\n30\n\n216\n\n12\n\n14\n\n48\n\n0\n\n0\n\n6 і 7\n\n96\n\n2–8, 10\n\n0\n\n0\n\n0\n\n0\n\n0\n\n0\n\n82\n\n0\n\n14\n\nВибірки 3-5, що розвивалися в однакових умовах, але ймовірно відрізнялися генотипом, значно відрізнялися за співвідношенням семи зареєстрованих у них типів ротових апаратів (при порівнянні за критерієм χ² p<0,00001). Схоже, на різноманітті ротових апаратів відображаються генотипічні особливості жаб. У вибірках 1 і 2, отриманих з природного середовища, не виявлені типи A, B, C і E, проте виявлені типи H і I. Нарешті, у вибірках 6 і 7, що складалися лише з головастиків P. ridibundus, зареєстровано лише 2 типи ротових апаратів: G і I. Найменше різноманіття типів ротових апаратів характерне для вибірок одного виду з природного середовища, а найбільше — для змішаних вибірок, що розвивалися в штучних умовах. Отже, на різноманіття ротових апаратів головастиків впливає і їх вид, і генотип, і умови їх розвитку, але жоден з цих факторів не може вичерпно пояснити спостережувану змінність їх ротових апаратів.\nSummary. The classification of types of scrape oral apparatuses of water frogs tadpoles (Pelophylax esculentus complex) has been developed. The variety of oral apparatuses types has been described in a few selections of tadpoles. Species, genotype and conditions of tadpoles development influence on the oral apparatus structure. The complete explanation of observed picture of oral apparatus mutability was not successful.\nАвтор дякує Т. Vinogradskoi, N. Tolokneevi, E. Komarovi та V. Khloponini за допомогу у виконанні роботи, а доцента D. A. Shabanova — за наукове керівництво.\nБібік Ю.С. Які фактори визначають тип ротового апарату головастиків зелених жаб? //\"Біологія : від молекули до біосфери\". Матеріали V Міжнародної конференції молодих науковців. - Харків: Оперативна поліграфія, 2010. - с. 339-340.\nВЛИЯННЯ ЕФЕКТУ ГРУПИ НА РОЗВИТОК ГОЛОВАСТИКІВ ЗЕЛЕНИХ ЖАБ\nБібік Ю. С., Коваленко М. С., Кучкова А. Г.\nХарківський національний університет імені В. Н. Каразина, біологічний факультет, пл. Свободи, 4, м. Харків, Україна. e-mail: yulia.bibik@gmail.com\nЗавдання даної роботи — встановити, чи впливає перебування у складі різновікового гурту на розвиток ротових апаратів і кінцівок головастиків представників Pelophylax esculentus complex. Як відомо (Mina, Klevezal, 1976), групове утримання водних тварин змінює їх онтогенез у порівнянні з одиничним.\nМи використали головастиків з річки Північний Донець, зібраних на лівому березі нижче м. Змієва 24.06.2010 р. Розглядаючи живих головастиків під стереоскопічним мікроскопом, ми обрали дві групи головастиків з однаковим будовою задніх кінцівок. 24 «малі» головастика знаходилися на 3‑ій стадії розвитку кінцівок (див. тези Vinogradskoi та ін.), 12 великих — на 7‑ій. В обох групах були представлені головастики з типами ротових апаратів (див. тези Бібік) G, D, H і I. Головастиків зважували і розміщували в ємкостях (половинні банки), що містили по 0,3 л річкової води. У 6 ємкостей помістили по 1 «малій» головастик, у 6 — по 1 «великій», і ще в 6 — по 3 «малі» і 1 «велику». Раз на добу воду міняли і годували головастиків надмірною кількістю сухого акваріумного корму. Через 10 днів провели повторні вимірювання маси, визначили стадії розвитку кінцівок і типи ротових апаратів.\nУ «малих» головастиків‑одинаків стадії розвитку кінцівок змінилися з 3‑ї до 6‑ї, а в групах — з 3‑ї до 4‑ї. У «великих» головастиків незалежно від умов утримання відбулося зміна з 7‑ї до 9‑ї стадії. Групове утримання значно (p=0,004) зменшило відносний приріст маси в обох групах, практично однаково для «великих» і «малих». Таким чином, групове утримання затримало ріст (оцінений за збільшенням маси) в обох групах, а розвиток (оцінений за станом кінцівок) — лише у «малих».\nТип ротового апарату у «малих» головастиків змінювався значно (при порівнянні за Манном‑Уїтні p=0,005) частіше, ніж у «великих». Групове утримання не вплинуло на частоту зміни типів ротових апаратів у «великих» головастиків і значно (p=0,32) знизило її у «малих». У «малих» головастиків зафіксовано такі комбінації початкового і кінцевого типів ротових апаратів: D→D (13 випадків); D→E (1); D→G (3); G→G (2); G→H (2); H→H (1); I→G (2). Для «великих» головастиків зафіксовано комбінації G→G (2 випадки); G→H (5); G→редукція ротового апарату (5 випадків). Остання група випадків нагадує події, що відбуваються під час метаморфози (коли скоблячий ротовий апарат редукується і розвивається ротовий апарат, призначений для захоплення здобичі), але ці процеси відбуваються на пізніших стадіях розвитку.\nSummary. Development of hind limbs and oral apparatus as well as mass growth have been studied in small and big tadpoles of water frogs(Pelophylax esculentus complex) maintained solitary or in groups (1 big and 3 small tadpoles in each group). Maintenance in groups inhibits growth both in small and big tadpoles, whereas hind limbs development is inhibited only in small tadpoles. Alteration of structure in various types of oral apparatus is also described.\nОписаний експеримент виконаний як навчально‑дослідницька робота на навчальній практиці II курсу. Автори дякують доцента D. A. Shabanova за наукове керівництво даною роботою.\nБібік Ю. С., Коваленко М. С., Кучкова А. Г. Вплив ефекту групи на розвиток головастиків зелених жаб. // \"Біологія : від молекули до біосфери\". Матеріали V Міжнародної конференції молодих науковців. Харків: Оперативна поліграфія, 2010. -с. 338-339.\nРОСТ І РОЗВИТОК ГОЛОВАСТИКІВ ЗЕЛЕНИХ ЖАБ В ПРИРОДНОМУ СЕРЕДОВИЩІ\nVinogradska T. I., Tolokneeva A. A., Komarova E. V., Khloponina V. V., Bibik Yu. S.\nХарківський національний університет імені В. Н. Каразина; пл. Свободи, 4, м. Харків, Україна\nЄвропейські зелені жаби (Pelophylax esculentus complex) — гібридогенний комплекс, що привертає увагу багатьох дослідників (Shabanov, Lytvynchuk, 2010). Для комплексного вивчення цієї групи тварин треба встановити, як проходить їх онтогенез, зокрема — личинковий розвиток. Після опису розвитку змішаних вибірок головастиків, взятих з природних ГПС, можна буде порівняти особливості личинкового розвитку окремих форм жаб.\nМи вивчили головастиків, зібраних на лівому березі Північного Донуця нижче за течією м. Змієва в заростях водної рослинності на мілководді. Вибірка № 1, зібрана 24 червня 2010 р., складається з 130 особин, а вибірка № 2, зібрана 1 липня 2010 р. — з 232 особин.\nМірами росту головастиків слугували загальна довжина тіла з хвостом і довжина тулуба (які ми вимірювали штангенциркулем з точністю до 0,1 мм), а також вага (яку вимірювали електронними вагами з точністю до 0,001 г). Стадії розвитку визначали за станом задніх кінцівок. Виділяли такі стадії: 1) відсутність кінцівок; 2) зачатки задніх кінцівок мають вигляд круглого бугорка; 3) зачатки кінцівок починають згинатися; 4) дистальний кінець кінцівки приймає форму лопаточки; 5) зачатки 3‑х пальців; 6) зачатки 4‑х пальців; 7) зачатки 5‑ти пальців, починає розвиватися колінний суглоб; 8) сформований колінний суглоб; 9) сформовані колінний суглоб і суглоб між гомілкою та другорядною гомілкою, перепонки між 3‑ма пальцями; 10) повністю сформована міжпальцева перепонка. Дані обробляли за допомогою програми Statistica for Windows 6,5.\nВ цілому зареєстрований зв’язок довжини (L, мм) і ваги (W, г) головастиків може бути екстраполований виразом W= 0,26 - 0,27 × L+ 0,098 × L². Значення довжини і ваги для окремих стадій показані в табл. 1.\nТабл. 1. Середні значення довжини і ваги головастиків на різних стадіях їх розвитку\n\nСтадія\n\nКількість особин (вибірки №1+№2)\n\nДовжина (L), мм\n\nВага (W), г\n\nСтадія\n\nКількість особин (вибірки №1+№2)\n\nДовжина (L), мм\n\nВага (W), г\n\n1\n\n4 + 24\n\n2,02\n\n0,107\n\n6\n\n22 + 20\n\n4,22\n\n1\n\n2\n\n37 + 74\n\n2,74\n\n0,265\n\n7\n\n19 + 8\n\n4,76\n\n1,261\n\n3\n\n23 + 22\n\n3,24\n\n0,408\n\n8\n\n11 + 17\n\n5,3\n\n1,698\n\n4\n\n5 + 12\n\n3,84\n\n0,657\n\n9\n\n1 + 11\n\n5,52\n\n1,788\n\n5\n\n7 + 29\n\n3,88\n\n0,659\n\n10\n\n1 + 15\n\n6,42\n\n2,597\n\nЯк видно з табл. 1, вибірки відрізняються за відносною чисельністю головастиків, що перебувають на різних стадіях розвитку. При порівнянні зареєстрованого розподілу з теоретичним, побудованим на підставі припущення про відсутність зв’язку вибірки і стадії розвитку, критерій Пірсона χ²= 34,4, що при даній кількості ступенів свободи відповідає високозначимим різницям (p = 0,000075). Переважаюча у вибірці № 2 кількість головастиків на пізніх стадіях, ймовірно, пов’язана з довшим часом їх розвитку.\nSummary. Described stage of the development of the hind limbs water frog (Pelophylax esculentus complex). The studied polliwogs caught in the river Severskiy Donec in a neighborhood of Zmiev. Dependency of the weight (W) polliwog from their lengths of the body (L) is of the form of W = 0,26 - 0,27 L + 0,098 L 2.\nОписані результати отримані в ході навчально‑дослідницької роботи на навчальній практиці з зоології хребетних. Автори висловлюють подяку доц. D. A. Shabanovu за керівництво даною роботою.\nVinogradska T. I., Tolokneeva A. A., Komarova E. V., Khloponina V. V., Bibik Yu. S. Рост і розвиток головастиків зелених жаб в природному середовищі // \"Біологія : від молекули до біосфери\". Матеріали V Міжнародної конференції молодих науковців. - Харків: Оперативна поліграфія, 2010. с. 344-345.\nДОСЛІДЖЕННЯ ФЛУКТУЮЧОЇ АСИМЕТРІЇ ПОКРИТТЯ У ЗЕЛЕНИХ ЖАБ\nKryshchal O. A., Matasova E. V.\nХарківський національний університет імені В. Н. Каразина, біологічний факультет, пл. Свободи, 4, м. Харків, Україна\nВизначення рівня флуктуючої асиметрії, FA (ненаправлених випадкових відхилень від біліатеральної симетрії) — один із способів оцінки стабільності онтогенезу. Розвиток у міжвидових гібридів зазвичай менш стабільний, ніж у батьківських видів. У околицях біостанції ХНУ в с. Гайдари (Зміївський р-н Харківської області) серед європейських зелених жаб живуть представники батьківського виду, Pelophylax ridibundus (озерні жаби) і геміклональні міжвидові гібриди, Pelophylax esculentus (їстівні жаби). У 2008 році FA їхнього покриття вивчали за загальноприйнятими ознаками — асиметричністю дорзомедіальної смуги, кількістю плям на спині та плям‑смуг на задніх кінцівках (Deryazhenkova et al., 2008). Як ни дивно, FA гібридів у цьому дослідженні не перевищував FA батьківського виду. Ми застосували для вимірювання FA жаб новий метод, припускаючи, що він виявиться більш чутливим до відхилень від симетрії покриття.\nУ поймі р. Північний Донець в околицях с. Гайдари було спіймано 186 зелених жаб. Їх визначали за зовнішніми ознаками: низький і косий п’ятковий бугор, відсутність жовтуватого відтінку на стегнах, темні резонатори у самців відповідає P. ridibundus; високий бугор, наявність жовтого відтінку на стегнах, світлі резонатори у самців відповідає P. esculentus. Ми визначили 107 особин (46 самок, 36 самців і 25 напівдорослих) як P. esculentus, 55 особин (31+12+12 відповідно) — як P. ridibundus, і 24 особи (12+6+6) залишилися невизначеними через нехарактерне поєднання їх ознак.\nСпинну сторону тулуба і кінцівок кожної жаби фотографували (для цього жабу давали схопитися передніми кінцівками за опору і розтягували її в довжину за пальці задніх кінцівок), а потім випускали в місця лову. У програмі CorelDraw на кожну фотографію була накладена сітка, що розбиває її вздовж осі тіла жаби, від рила до п’яткових бугорів, на 20 зон. Для всіх зон, для яких це було можливо, визначали, чи симетрична в ній покриття (S), слабо асиметрична (H) або сильно асиметрична (A). Підраховували кількість зон кожного типу: NS, NH і NA. Результати обробляли в програмі Statistica. Асиметричність особини обчислювали як FA = (NA+ NH/2) / NS.\nОзнаки FA, що враховують знак відхилення від симетрії, мають нормальний розподіл зі середнім значенням 0. Наша міра FA приймає лише позитивні значення. Найбільш чисельними виявилися значення, близькі до 0, з ростом показника FA частота зустрічальності значень зменшувалась. Таке розподілення підтверджує, що використана міра характеризує FA і може застосовуватись для вивчення стабільності розвитку жаб, і, ймовірно, деяких інших тварин. P. esculentus виявилися значно (при порівнянні за Манном‑Уїтні p=0,031) асиметричнішими, ніж P. ridibundus. Медіана використаної міри FA для P. esculentus становить 0,4, для P. ridibundus — 0,33, а для невизначених особин — 0,39. Вплив статі та віку на FA покриття виявився незначним.\nSummary. For study fluctuating asymmetry (FA) tinge in water frogs, their photographed from back side. Photography was dividing along axis body on 20 zones. We counted the symmetrical, weak and hard asymmetrical zones. The tinge of hybrids form water frogsP. esculentus was found a more asymmetrical, then tinge of representatives by generation species water frogsP. ridibundus.\nРобота виконана як УІРС на навчальній практиці з зоології хребетних. Автори дякують доцента D. A. Shabanova за ідею роботи і наукове керівництво нею.\nKryshchal O. A., Matasova E. B. Дослідження флуктуючої асиметрії покриття у зелених жаб. // \"Біологія : від молекули до біосфери\". Матеріали V Міжнародної конференції молодих науковців. - Харків: Оперативна поліграфія. 2010. - с. 355-356."
}