Бондарева, Шабанов (2011) Порівняння розмірів та форми еритроцитів...
Взагалі я вважаю неправильним дописуватись співавтором до студентських тез. Але в даному випадку від загальних правил я відступив: вважаю цей випадок нетривіальним. На жаль, час йде, а ми досі не можемо знайти жаб з маленькими еритроцитами, аналогічними описаним, для більш детального вивчення....
Бондарева А.А., Шабанов Д.А. Порівняння розміру та форми еритроцитів у диплоїдних і тріплоїдних гибридів зелених жаб (Pelophylax esculentus) // Всеукраїнський конкурс студентських наукових робіт з біологічних наук: матеріали підсумкової науково‑практичної конференції (Мелітополь, 29‑31 березня 2011 р.). – Мелітополь: «Люкс», 2011. – С. 11‑14.
УДК 004.942:597.851
ПОРІВНЯННЯ РОЗМІРУ ТА ФОРМИ ЕРИТРОЦИТІВ У ДИПЛОЇДНИХ І ТРІПЛОЇДНИХ ГІБРИДІВ ЗЕЛЕНИХ ЖАБ (PELOPHYLAX ESCULENTUS)
Бондарева А. А., Шабанов Д.А.
У склад гибридного комплексу зелених жаб (Pelophylax esculentus complex) входять два батьківські види, ставкові жаби, Pelophylax lessonae (Camerano,1882) і озерні жаби Pelophylax ridibundus (Pallas, 1771), а також їх гибриди, їстівні жаби, Pelophylax esculentus (Linnaeus, 1758). У типічному випадку гибридні особини виробляють гамети, що несуть клональний геном одного з батьківських видів. Таке розмноження називається геміклональним, або напівклональним.
Ще однією особливістю зелених жаб є те, що їх гибриди можуть бути не лише диплоїдними, а й тріплоїдними. Ди‑ і тріплоїди практично не відрізняються на організмовому рівні, тому що вони схожі за розміром, формою тіла і тривалістю життя. На сьогодні не описані біохімічні відмінності між ди‑ і тріплоїдними гибридами (не рахуючи ефектів, пов’язаних з дозою генів — наслідком наявності у тріплоїдів двох геномів одного з батьківських видів). Тим цікавіші суттєві відмінності між ди‑ і тріплоидами на клітинному рівні. Оскільки у тріплоїдів у ядрі міститься не два, а три набори хромосом, їхнє ядро має більший, ніж у диплоїдів, об’єм. Водночас важливим параметром клітини є ядерно‑цитоплазматичне відношення, коливання якого можуть впливати на функціонування окремих клітин і організму в цілому. Тому для збереження цього відношення у тріплоїдів після ядра має збільшуватись і об’єм цитоплазми, а отже й розмір клітини [1, 3].
Для диплоїдних і тріплоїдних жаб зафіксовано значуще різницю в розмірах еритроцитів, яку використовують як один із способів визначення плоїдності жаб [3, 4]. Для риб показано, що тріплоїдність у різній мірі впливає на різні морфологічні параметри еритроцитів [2]. Для жаб докладне вивчення змін різних параметрів еритроцитів не проводилось. Це і стало метою даної роботи.
Ми досліджували 58 жаб, визначених за морфологічними ознаками як P. esculentus. Для кожної особини був зроблений мазок крові за стандартною методикою. Участок препарату фотографували під великим збільшенням мікроскопа за допомогою USB‑камери. Під тим же збільшенням сфотографували об’єкт‑мікрометр. За допомогою програми PDF‑XChangeViewer вимірювали велику і малу осі еритроцитів і їх ядер (по 10 клітин на мазок); потім ці вимірювання переводили в мікрометри. На підставі цих даних розраховували площі перерізу (площа на препараті), площі поверхні і об’єми еритроцитів і їх ядер. Обробку даних проводили за допомогою програми Statistica 8.0.
За розмірами еритроцитів ми розділили вивчених жаб на три групи (рис. 1). У групу I ввійшли три особи з аномально малими еритроцитами, у групу II — 43 особи (ймовірно, диплоїдних), а у групу III — 12 особей (ймовірно, тріплоїдних). Результати проточної ДНК‑цитометрії 14 особин з II і III груп, виконані С.Н. Литвинчуком і Ю.М. Розановим в Інституті цитології РАН (м. Санкт‑Петербург), підтвердили, що у склад II групи особин входять диплоїдні P. esculentus, а у склад III — тріплоїдні P. esculentus.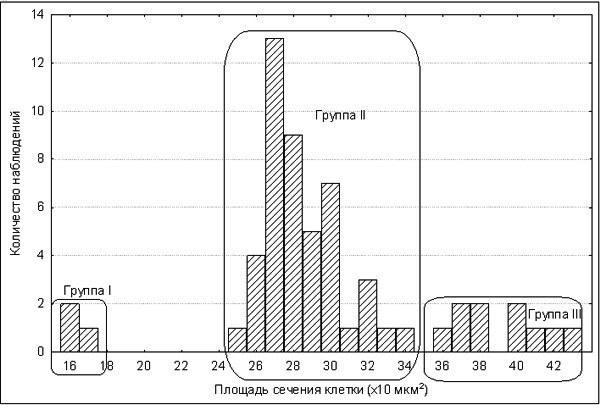
Рис. 1. Розподіл жаб за середньою площею перерізу їх еритроцитів
Порівняння морфологічних параметрів типових ди‑ і тріплоїдних клітин показало, що тріплоїдність призводить до значного (р < 0,001) збільшення майже всіх параметрів клітини, за винятком двох: відбувається незначне зменшення ядерно‑цитоплазматичного відношення (р = 0,16) і значне зменшення відносної поверхні клітини (на 10,6 %; р < 0,001). Збільшення відбувається в різній мірі для різних показників. Так, збільшення великої осі клітини і ядра становить 22,9 і 18,6 % відповідно, а малих осей — 9,7 і 9,4 %. Це призводить до того, що тріплоїдна клітина набуває більш еліпсоїдної форми. Збільшення більшості морфологічних параметрів еритроцитів і зменшення відносної поверхні клітини у тріплоїдів мають призводити до зміни інтегральної інтенсивності газообміну. Зміна форми еритроцита, можливо, пов’язана з адаптацією більших еритроцитів до відносно вузьких кровоносних судин.
Найбільший інтерес викликають еритроцити особин групи I. Різниця між групою I і II за розмірними параметрами виявилася такою ж або більшою, ніж різниця між ди‑ і тріплоидами (рис. 1, 2).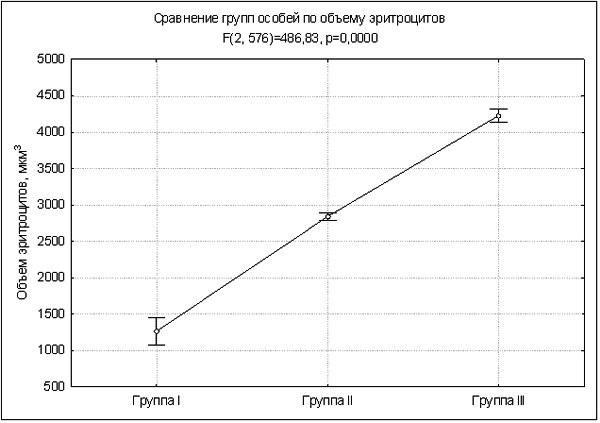
Рис. 2. Об’єм еритроцитів, характерний для трьох груп жаб (вертикальні лінії демонструють довірчі інтервали)
Саме такі параметри, які зареєстровані для еритроцитів жаб групи I, мали б мати гіпотетичні гаплоїдні еритроцити. Пояснити спостережуваний феномен могли б два альтернативних припущення: (1) в організмі жаб відбувається гаплоїдація соматичних клітин, аналогічна премейотичній елімінації одного з геномів у клітинах зародкової лінії; (2) в еритроцитах особин I групи відбувається функціональна гаплоїдація клітин, пов’язана, ймовірно, з гіперконденсацією одного з геномів. Дослідження, які будуть виконані протягом польового сезону 2011 року, мають дати відповідь, чи відповідає дійсності одне з цих пояснень.
Автори щиро вдячні С. Н. Литвинчуку і Ю. М. Розанову (Санкт‑Петербург) за неоціненну допомогу в ідентифікації вивчених особин, а також Я. B. Афонічевій, Е. В. Баланюк, А. Е. Кечеджи і Т. І. Махній за допомогу у зборі матеріалу і виконанні практичної частини роботи.
СПИСОК ЛІТЕРАТУРИ
1. Shabanov D. A., Lytvynchuk S. N. Зелені жаби: життя без правил або особливий спосіб еволюції // Природа. — 2010. — № 3 (1135). — С. 29‑36.
2. Cal R.M., Vidal S., et al. Effect of triploidy on turbot haematology. // Comparative Biochemistry and Physiology, Part A (141). - 2005. - P. 35-41.
3. Plötner J. Die Westpaläarktichen Wasserfrösche. Bielefeld, 2005 - 161 S.
4. Polls Pelaz M, Graf J.-D. Erythrocyte size as an indicator of ploidy level in Rana kl. esculenta before and after metamorphosis. // Alytes. - №7 (1988). - P. 53- 61.