
Стаття
Shabanov, Lytvynchuk (2010) Зелені жаби: життя без правил чи особливий шлях еволюції?
Розміщую тут у найбільш зручному для читання з екрана вигляді статтю, яка є для нас основним введенням у проблему вивчення гібридизації зелених жаб. Це огляд, підготовлений разом зі Спартаком Литвинчуком для російського журналу "Природа": Шабанов Д. А., Литвинчук С. Н. Зелені жаб...
{
"translated_text": "Шабанов Д. А., Литвинчук С. Н. Зелені жаби: життя без правил або особливий шлях еволюції? // Природа. – 2010. – № 3 (1135). – С. 29–36.\nЗелені жаби: життя без правил чи особливий спосіб еволюції?\n© Шабанов Д. А., Литвинчук С. Н., 2010\n\n

\n\n\nДмитро Андрійович Шабанов, кандидат біологічних наук, доцент кафедри зоології та екології тварин Харківського національного університету ім. В.Н.Каразіна. Область інтересів — популяційна екологія, батрахологія, еволюційна біологія\n\n\n\n\n

\n\n\nСпартак Миколайович Литвинчук, кандидат біологічних наук, старший науковий співробітник Інституту цитології РАН. Область інтересів — цитогенетика, зоологія, еволюційна біологія\n\nСучасні уявлення про організми, популяції та види створені зусиллями багатьох поколінь біологів. Пізнаючи світ, вони намагалися побудувати упорядковану картину дійсності, систематизувавши всіх мешканців планети. Основна одиниця систематики — вид — це не просто найменування організму, а й якісний етап еволюції, сукупність особин, пов'язаних процесом розмноження та визначеною еволюційною долею.\nБільшість тварин та рослин розмножуються статевим шляхом, під час якого новий організм отримує два набори спадкової інформації (геноми) — від матері та від батька. У геномах нащадків спадкові задатки батьків поєднуються (рекомбінуються) за ймовірністними законами, що породжує невичерпну генетичну різноманітність.\nОсобини одного виду існують у складі певних популяцій. Під впливом ізоляції дві частини колись єдиної популяції можуть змінитися настільки сильно, що схрещування між їхніми представниками стане ускладненим. Так виникають нові види.\nУсе це вірно, але не завжди. Як відомо, немає правил без винятків. Пошук таких винятків, точніше порушень відомих біологічних законів, — один із способів розуміння феномену життя. Розглядаючи їх, можна перейти від уже зрозумілих «правил гри» до глибших істин.\nВідступати від згаданих «правил гри» та від багатьох інших здавалося б цілком вивченим тваринам — наприклад, жабам. Звісно, у XX ст. — в епоху біохімії, генетики та молекулярної біології — класичними об'єктами досліджень стали миші, щури, дрозофили та круглий черв'як Caenorabditis. Але до цього довгий час досліджуваними тваринами біолога (та навіть нігіліста Базарова) були саме жаби, і найчастіше зелені (Rana sp.). Саме вони сидять уздовж берегів майже будь-якого прісноводного водоймища та шалепають у воду під час наближення людини.\n\nВсе не як у людей\nНа початку 60-х років XX ст. польський гідробіолог Лешек Бергер захотів розібратися в плутанині, що гуляла сторінками зоологічних визначників того часу. Було неясно, два чи три види (чи підвиди) жаб заселяють польські водоймища. Він пішов просто: розсадив жаб за парами у невеличкі ставки у власному городі та дочекався їхнього потомства. Один із «видів» жаб виявився гібридом двох інших. Ну що ж, буває й так. Парадоксальнішим було те, що при схрещуванні гібридів з особинами батьківського виду отримували або жаби батьківського виду, або такі ж самі гібриди [1]. Відступлення від канонів викликало пристальний інтерес дослідників. Загадку розв'язав німецький біолог Хайнц Тюннер. Він спочатку здогадався, а потім за допомогою генетичних маркерів довів, що у гібридних жаб у статеві клітини переходить лише один із батьківських геномів [2]. Аналогічне явище незадовго до того виявили у одного американського роду живородящих риб та назвали гібридогенезом.\nВідкриття Бергера та Тюннера були лише початком череди чудес, пов'язаних із зеленими жабами. Відступивши від історії, коротко опишемо особливості цієї групи. Східноєвропейські зелені жаби представлені двома батьківськими видами та їхніми гібридами: озерна жаба (Rana ridibunda)* та ставкова жаба (R. lessonae), що схрещуючись, породжують їстівних жаб (R. esculenta) (рис. 1).\n*У останні роки багато систематиків виділяють цих жаб у окремий рід Pelophylax і називають відповідно — P. ridibundus, P. lessonae та P. esculentus.\n\n

\nРис.1. За цим екземпляром сам Карл Лінней описав вид Rana esculenta! Можна вважати, що це — жаба №1, тобто носій видового імені, або голотип. Фото С.Н.Литвинчука\n\nСтрого кажучи, звичайні гібриди видових імен не отримують, і їстівні жаби зберегли своє ім'я не тому, що його дав у 1758 р. Карл Лінней, а внаслідок того, що вони представляють окремий ланцюг еволюції, здатний не лише підтримувати себе у взаємодії з батьківськими видами, а й навіть відтворюватися самостійно (рис. 2, 3).\n\n
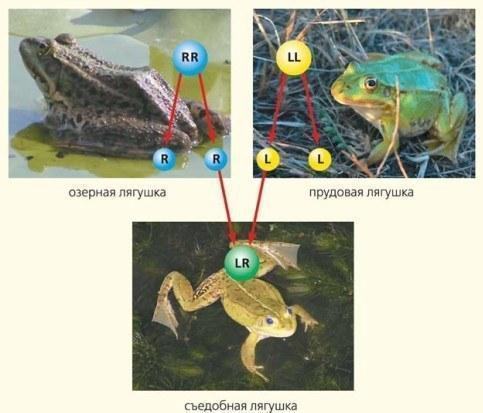
\nРис.2. Схема первісного виникнення гібридів. L — геном ставкової жаби, R — озерної. Особини з генотипом LR отримуються від схрещування двох батьківських видів — LL та RR. Тут і далі фото А. В. Коршунова та Д. А. Шабанова\n\n
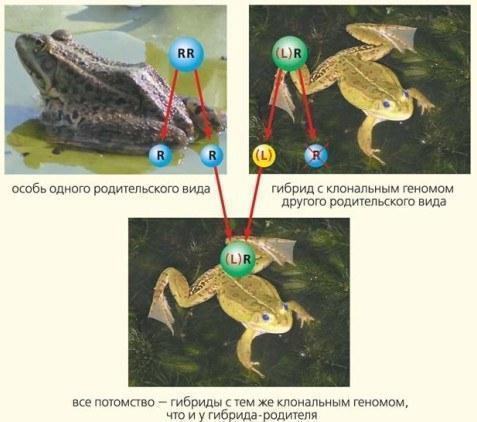
\nРис.3. Схема відтворення гібридних жаб при їхньому схрещуванні з особинами батьківського виду. Символ клонового геному (у даному випадку — геному ставкової жаби L) взято у дужки.\n\nЩоб простіше усвідомити нестандартність способу розмноження гібридних жаб, порівняємо утворення статевих клітин у них та у людей. Клітини нашого тіла мають по 46 хромосом, а жаб — по 26. У обох випадках половина цих хромосом отримана від матері, а половина — від батька. У людей у статевих клітинах формується фактично новий геном, що складається з перемішаних «материнських» та «батьківських» фрагментів. Кількість можливих комбінацій при цьому неймовірна. Навіть проста перетасування хромосом (у яку статеву клітину увійде «материнська», а в яку — «батьківська») дає 2^23 комбінацій. Насправді варіантів незрівнянно більше, адже після обміну ділянками між хромосомами кожна з них (крім статевих, X та Y) перестає бути «материнською» чи «батьківською», стаючи новою, рекомбінантною. Серед сотень мільярдів сперматозоїдів, які чоловік виробляє за своє життя, немає двох однакових. Сенс такої «перетасування» — пошук вдалих комбінацій спадкових задаток. У цьому відношенні людина — цілком звичайний представник тварин, а от жаби — не зовсім.\n\nУ жаб рекомбінація спадкової інформації відбувається під час розмноження особин одного виду, а от у гібридів, що виникають при схрещуванні жаб різних видів, все інакше! Такі особини виробляють ідентичні (клонові*) статеві клітини, що несуть один із батьківських геномів. А що трапляється із другим? Він просто знищується, елімінується. Таке спадкування називається геміклональним (від грец. γεμι — пів- та κλων— нащадок, гілка).\n*Клоновість геному ми позначаємо взяттям його символу у дужки: якщо клоново передається геном ставкової жаби — (L), якщо озерної — (R).\n\nА до чого призводить схрещування гібридних жаб з особинами батьківського виду? Усі їхні нащадки отримують один і той самий клоновий геном та різні рекомбінантні геноми (рис.3).\n\nТепер уявімо ситуацію, коли в одному водоймищі перебувають особини, що належать до різних видів та форм. Разом розмножуючись, вони передають як рекомбінантні, так і клонові геноми. Таку сукупність особин можна назвати геміклональною популяційною системою (ГПС) (рис. 4, 5). У ГПС, де, припустімо, розмножуються озерні жаби — RR та гібриди, що клоново передають геном ставкової жаби — (L)R, можливі три варіанти схрещування. Під час розмноження озерних жаб усі їхні нащадки належать до одного виду: RR×RR→RR. Схрещування озерної жаби з гібридами призведе до появи гібридного потомства: RR×(L)R→(L)R. А що буде при схрещуванні гібридів?\n\n

\nРис. 4. Розмноження в складі геміклональної популяційної системи: у загальному нерестовому котлі зустрілися особини батьківського виду та різноманітні гібриди. Їх легко розрізнити за кольором резонаторів самців: у їстівної жаби (гібрида озерної та ставкової жаб) вони білі, у самців озерної — сірі. Фото Д.А.Шабанова\n\n
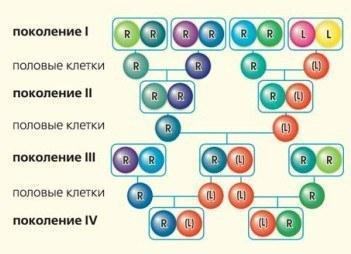
\nРис.5. Схема історії кількох поколінь жаб, заснована на аналізі передачі геномів (кольорові кульки). У разі спадкування з рекомбінацією у статеві клітини переходить геном, що утворюється при змішуванні спадкової інформації батьків (показано змішуванням кольорів). Клонові геноми передаються як єдине ціле (колір зберігається).\n\nЯк би не було дивно, у багатьох випадках схрещування гібридів, що передають клоновий геном одного виду, призводить до появи нежиттєздатного потомства: (L)R×(L)R→[(L)(L)]→†. Цей феномен традиційно пояснюють накопиченням мутацій у клоновому геномі. Оскільки він передається з покоління в покоління без оновлення в процесі рекомбінацій, він не може позбутися неминуче виникаючих у ньому летальних мутацій. Запам'ятаймо цей факт та повернімося до розгляду долі системи, що складається з особин з генотипами RR та (L)R. Схрещування «гібрид-батьківський вид» призведе до збільшення частки гібридів, доки в такій геміклональній популяційній системі не зникнуть представники батьківського виду. ГПС, що складається з гібридів, які не здатні породити життєздатне потомство, загине...\n\nНа щастя гібридних жаб та їх дослідників, так відбувається не завжди. По-перше, перевага у відтворенні гібридів може бути врівноважена їхньою зниженою життєздатністю та плодовитістю, що призведе до стабілізації чисельності двох форм у популяції у певному співвідношенні. По-друге, існують різні форми гібридів, і деякі з них, що мешкають разом, здатні породжувати цілком стійкі популяційні системи.\n\nОсобливості життя без правил\nНам пощастило взяти участь у відкритті 2002 р., а також подальшому вивченні незвичайного центру різноманіття зелених жаб — регіону, де одночасно зустрічається багато їхніх форм. Цей центр різноманіття пов'язаний із р. Північно-Донець [3-5].\n\nКрім озерних жаб та диплоїдних (з двома геномами) гібридів, у цьому центрі масово представлені триплоїдні гібриди, тобто особини з трьома геномами (таблиця, рис. 6)! Серед молодих особин зареєстровано навіть одиничних тетраплоїдів (по два генома ставкової та озерної жаб) та ставкових жаб. Ймовірно, тетраплоїдні гібриди та ставкові жаби гинуть у молодому віці. І це ще не все. Диплоїдні гібриди у цих ГПС представлені трьома формами, що відрізняються за складом їхніх статевих клітин. Ці форми можна позначити (L)R, L(R) та (L)(R).\n\nТаблиця\nСклад геміклональної популяційної системи у Північно-Донецькому центрі різноманіття зелених жаб (околиці біостанції Харківського університету у с. Гайдари та Національного парку «Гомільшанські ліси»)\n\n\n\n\nГенотипи\n\n\nR\n\n\nLR\n\n\nLLR\n\n\nLRR\n\n\nLLRR\n\n\nLL\n\n\n\n\nСеголетки\n\n\n+++\n\n\n+++\n\n\n++\n\n\n+++\n\n\n+\n\n\n+\n\n\n\n\nСамки\n\n\n+++\n\n\n++\n\n\n+\n\n\n++\n\n\nЙмовірно, не доживають до половозрелості\n\n\n\n\nСамці\n\n\n+++\n\n\n+++\n\n\n+\n\n\n++\n\n\n\n\nСтатеві клітини самців\n\n\nR\n\n\nL\n\n\nR\n\n\nL та R одночасно*\n\n\nL\n\n\nR\n\n\n\n\nСтатеві клітини самок**\n\n\nR\n\n\nL\n\n\nR\n\n\nL та R одночасно\n\n\nLR\n\n\nLR\n\n\n\n\n*Кількість плюсів показує обмеження різних форм.\n* У рідкісних випадках спостерігалися й гамети зі змішаними (гібридними) геномами.\n* Статеві клітини самок наведено на основі припущень, адже їхнє вивчення — набагато складніша проблема, ніж вивчення статевих клітин самців; інші результати зафіксовано за допомогою проточної ДНК-цитометрії та підтверджено комплексом інших методів.\n\n
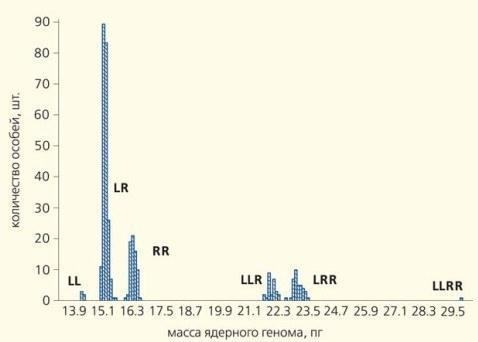
\nРис.6. Результати проточної ДНК-цитометрії, що дозволяє визначити масу ДНК буквально в кожній клітині. На підставі очевидних відмінностей у масі геномів ставкової (LL) та озерної (RR) жаб, а також чотирьох форм їхніх гібридів (LR, LLR, LRR та LLRR), отримані, зокрема, дані, наведені в таблиці\n\nХоча всі перераховані форми (див. табл.) були відомі науці й раніше, системи такого складного складу раніше ніхто не знаходив. Найбільш різноманітні ГПС знайдено у Харківській області України, але зона поширення триплоїдів простягає вниз за течією Північно-Донця аж до Ростовської області Росії [6].\n\nУ самому жі, нормальна життєздатність триплоїдних гібридів погано узгоджується з традиційними уявленнями про генетику тварин. Роздумайте: геноми як ставкової, так і озерної жаби — результат тривалої еволюції диплоїдних організмів з ідентичними хромосомними наборами. Здатність трьох таких геномів, що зустрілися в одній особині у співвідношенні 2:1, забезпечувати її нормальний розвиток та розмноження — парадокс! До речі, дані, отримані під час визначення віку жаб за допомогою врахування «річних кілець» — ліній, що формуються під час зимівки в їхніх трубчастих кістках (рис. 7), свідчать, що триплоїди, які дожили до половозрелості, живуть не менше, а навіть трохи швидше ростуть, ніж диплоїдні гібриди.\n\n
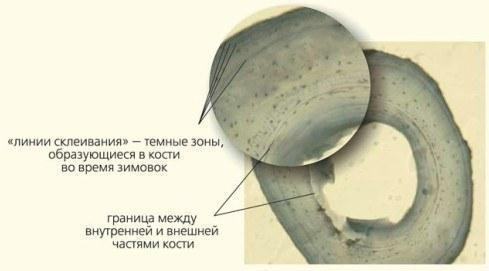
\nРис. 7. Зріз фаланги жабачого пальця. Так визначають вік та динаміку росту жаб. Фото Є.Є.Усової\n\nЯкщо вас це не здивовано, врахуйте, що триплоїди «побудовані» з більших клітин. Справа в тому, що розмір ядра клітини визначається кількістю генетичного матеріалу, що в ньому міститься, а співвідношення розмірів ядра та цитоплазми клітини — приблизно сталою величиною для кожної тканини. У півтори рази більше хромосом — у півтори рази більший об'єм ядра клітини — у півтори рази більша та маса самої клітини. А от розміри та маса самих жаб не зростають! Триплоїди, якщо й обганяють диплоїдів у зростанні, то ненабагато й лише в дуже «зрілому» віці. Є цілий ряд процесів, для яких важливі розміри клітин.\n\nПоведінка триплоїдів, принаймні на перший погляд, нічим особливо не відрізняється від поведінки диплоїдів. Проте з усього сказаного випливає, що функціонування нервної системи триплоїдів має відрізнятися від норми. Або клітин у мозку триплоїдів дійсно менше, або вони функціонують інакше, ніж диплоїдні. Отже, трьом геномам, що еволюціонували в різних видах, доводиться керувати жабачим розвитком із суттєвими відмінностями від норми. І нічого — впоруються!\n\nЩе непривичніше для традиційних уявлень існування гібридних особин, що виробляють статеві клітини, що належать або одному, або іншому батьківському виду. Можливо, це наслідок мозаїчності їхніх тканин, боротьби двох геномів, кожен з яких пристосувався знищувати суперника, або якогось іншого механізму. Якою б не була причина існування таких особин, у будь-якому випадку це — інтелектуальний виклик для сучасної науки.\n\nДумаєте, що проблема зелених жаб — лише деталь, що стосується єдиної групи тварин? Ні. Справа в тому, що у цьому та подібних випадках виявляються принципи роботи найзагальніших механізмів. Одна з таких проблем — проблема мейозу. І ось чому.\n\nТварини, що існують у змінній середовищі, змушені поєднувати консерватизм свого розвитку з пошуком новаторств. Справжнім проривом у еволюції стало оплідотворення — злиття двох клітин, що поєднують свій еволюційний досвід, записаний у їхніх геномах. Проте нова клітина матиме вдвічі більший набір спадкової інформації! Отже, у життєвих циклах організмів, де є оплідотворення, має бути не лише мітоз (клітинне ділення, під час якого комплекс спадкової інформації передається клітинам-нащадкам без змін), а й мейоз — ділення, що скорочує спадкову інформацію вдвічі (рис. 8)! Переважна більшість дійсно складних організмів на нашій планеті використовує мейоз у своєму життєвому циклі. Мейоз у форамініфер (морських «амеб», чиї раковини утворили шари крейди), яблуні, білому грибі та людині має свої особливості, але загалом подібний.\n\n
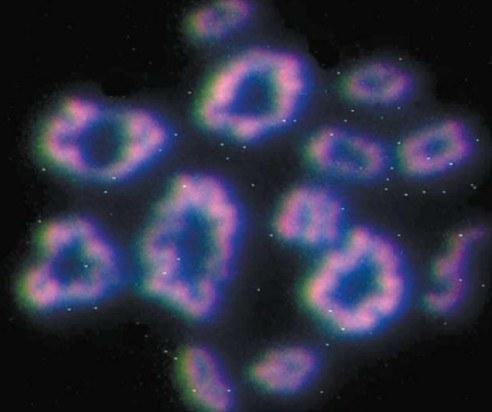
\nРис.8. Так виглядають хромосоми, що утворилися під час мейозу в клітинах яєчників жаб (флуоресцентне забарвлення). Сталася зміна ділянок у хромосомах, і вони відходять одна від одної, але все ще з'єднані кінцями. Фото Н.А.Агафоновой\n\nГеміклональне спадкування у гібридних зелених жаб — наслідок порушення мейозу. А різноманітність складу гамет, що виробляють різні гібриди, означає, що порушено він у них по-різному! Геноми ставкової та озерної жаб, що зустрічаються в одній особині, породжують таку нестійкість мейозу, яка може призводити не до однієї його аномалії, а до кількох. Стійкість таких ГПС, які ми вивчаємо у північно-донецькому центрі різноманіття зелених жаб, пов'язана саме з тим, що в них співіснують та відтворюються особини з різними аномаліями мейозу, що виробляють різні гамети.\n\nОдна з цікавих особливостей Північно-Донецького центру різноманіття жаб полягає в тому, що він розташований за межами ареалу ставкової жаби (див. табл.) [7]. Це означає, що геноми L у цьому центрі передаються від гібридів до гібридів протягом тривалого часу. Згідно з усіма канонами популяційної генетики, такі «відірвані від коріння» геноми мали б втратити генетичне різноманіття та просто дегенерувати. Чомусь цього не відбувається. Можливо, у Північно-Донецькому центрі важливу роль у підтримці життєздатності геномів ставкової жаби відіграють триплоїди LLR, які у чомусь грають роль особин батьківського виду [8]. Проте в інших регіонах гібриди можуть виходити за межі ареалу одного з батьківських видів і не утворювати при цьому триплоїдів. Ще одна загадка...\n\nЩе одна незвичайна особливість гібридних жаб полягає в тому, що їхні геноми виконують різні функції. Низька життєздатність особин із двома клоновими геномами одного виду свідчить, що виживання особини в більшій мірі визначає рекомбінантний геном. Проте важлива задача клонового геному — перейти у статеві клітини самому та не пустити туди інший геном. У гібридів першого покоління, отриманих від схрещування особин батьківських видів, виробництво статевих клітин стикається з великими проблемами. Багато з гібридів практично безплідні. У наступні покоління передаються лише ті клонові геноми, чиї володарі змогли забезпечити формування життєздатних статевих клітин. У результаті гібриди, чиї клонові геноми пройшли через цілий ряд поколінь, відтворюються набагато надійніше. Це свідчення особливої форми еволюції геномів — еволюції, в якій відбувається накопичення пристосувань до клонової передачі.\n\n

\nРис.9. Самці триплоїдних гібридів.Фото С.Н.Литвинчука
}{
"translated_text": "Петербурзькі герпетологи Лев Якович Боркін та Ілля Сергійович Даревський близько 30 років тому припустили, що триплойдні гібриди можуть відігравати важливу роль у виникненні нових гібридних видів. Загалом запропонована ними схема така: при схрещуванні диплоїдних видів виникають клонально розмножуючіся триплойдні гібриди. У результаті розмноження триплойдів та їх схрещування з іншими формами виникають тетраплойди, які переходять до нормального розмноження з рекомбінацією геномів [9]. Може, у Північно-Донецькому центрі відбуваються саме такі процеси? Геміклональні триплойди напевні, одиничні тетраплойди — теж. Але чомусь тетраплойди не доживають до статевої зрілості... Еволюція клональних геномів має призвести до якогось нового якості? Чи тетраплойдам для їх розвитку потрібні інші умови, ніж жабам із меншою кількістю хромосомних наборів?\n\nЩе один «пласт» еволюційних змін зелених жаб пов’язаний із порушенням клональної передачі геномів у гібридів. Іноді в клональні геноми потрапляють фрагменти з рекомбінантних геномів іншого виду, і навпаки. Передаючись від гібрида до гібрида у складі генопарадигматичних систем (ГПС), такий частково рекомбінований клональний геном може потрапити до озерної жаби. Так відбувається перенесення спадкової інформації через видові бар’єри!\n\nТакий перенос не обмежується лише ядерною ДНК, а стосується й мітохондріальної (мтДНК). Як відомо, вона знаходиться не в клітинному ядрі, а в мітохондріях та успадковується інакше — виключно за материнською лінією, з цитоплазмою яйцеклітин. Виявилося, що у більшості озерних жаб у Західній Європі взагалі немає власної мтДНК [10]. Вона повністю замінена на мтДНК прудової жаби, отриманою від гібридів. І справа не лише в тому, що мітохондрії прудової жаби виявилися «кращими» за мітохондрії озерної. Щоб отримати їх, західноєвропейські озерні жаби мали мати досить дивну генеалологію. Уявіть: у кожної озерної жаби одна з «бабусь» за материнською лінією мала бути прудовою жабою — представником іншого виду. А ядерні гени у неї — лише свого виду. Отже, ця «бабуся» із прудових жаб схрещувалася з самцем озерної. У результаті мав з’явитися гібрид із мітохондріями та материнським геномом прудової жаби й відцовським геномом — озерної. Стать у жаб успадковується майже так само, як у людей: у самиць — два жіночих геноми, а у самців — один жіночий, другий — чоловічий.\n\nОтже, «дочері» тієї самої «бабусі» прудової жаби мали передати нащадку жіночий геном озерної жаби та схрещуватися з самцем озерної жаби. Самиці, що утворилися від такого схрещування, стали предками за материнською лінією усіх озерних жаб у Західній Європі.\n\nЩо ж у «сухому залишку»? Група тварин із нетипово діючим спадковим апаратом, який еволюціонує нестандартним шляхом. Ця група утворює не популяції, а особливі біосистеми — генопарадигматичні системи (ГПС), що розвиваються за своїми законами. У межах цієї групи спадкова інформація подолає видові бар’єри, і, дуже ймовірно, на наших очах народжується новий тетраплойдний вид.\n\nЯк і навіщо все це вивчати?\n\nВузли такої складності неможливо розплутати за допомогою одного лише інструменту. Просту деталь можна зобразити на кресленні з однієї точки зору, складну вимагає декількох проекцій та перерізів. Шлях еволюції зелених жаб можна зрозуміти, лише об’єднавши зусилля фахівців різних галузей. Ось неповний перелік методів, які використовували ми з колегами* для опису процесів, що відбуваються із зеленою жабою: польові збори та обліки, опис динаміки росту за перерізами кістки, порівняння форми тіла та особливостей розвитку, математичне моделювання розмноження різних форм жаб у ГПС, визначення форм жаб за розміром їх геному, вивчення білокових маркерів, порівняння послідовностей ядерної та мтДНК, визначення характерних особливостей хромосом (імуннофлуоресцентне забарвлення, аналіз «лампових щіток») тощо. Під час роботи доводилося порівнювати жаб різних форм, із різних типів середовищ, ГПС та регіонів. Природно, для розуміння того, що відбувається із жабами, важливими є результати вивчення інших груп тварин, у яких зареєстровано подібні феномени: риб, зелених жаб, хвостатих земноводних, ящірок.\n\n*Ми щиро вдячні Л. Я. Боркіну, Г. А. Ладе, Ю. М. Розанову, М. В. Владимировій, Д. В. Дедуху, А. І. Зіненку, В. В. Клименку, А. В. Коршунову, М. А. Кравченку, Г. А. Мазепі, Є. Є. Усовій, С. Ю. Морозову-Леонову, Й. Плетнеру та багатьом іншим колегам за допомогу та підтримку під час проведення наших досліджень.\n\nДля вирішення настільки складної проблеми не вистачить сили ні одного автора, ні одного наукового колективу. Проблеми, подібні до тієї, яку ми розглядаємо, якраз і здатні об’єднувати наукове співтовариство, пов’язуючи вчених із різних країн.\n\nА навіщо вивчати зелених жаб? Головна для авторів цієї статті відповідь — тому що це дуже цікаво. Цікавість, недовіра, дослідницький азарт, які викликають описані у статті особливості жаб, належать до найприродніших двигунів науки. Але справа не лише в інтересі. Здогадуватися, які «бонуси» можна отримати від вивчення тієї чи іншої проблеми — неблагодарне заняття, але деякі спроби ми все ж зробимо.\n\nГеміклональне успадкування — спосіб прискорення еволюції; вивчення механізмів геміклональності відкриє нові можливості для селекції.\n\nПри геміклональному успадкуванні усі нащадки від схрещування двох особин (кожна з яких передає клональний геном) можуть бути генетично ідентичними. Можливість отримання таких тварин буде корисною для біотехнології та сільського господарства.\n\nОволодіння механізмами, які забезпечують збереження життєздатності гібридів із районів, де неможливе схрещування батьківських видів, допоможе підтримувати життєздатність порід та чистих ліній сільськогосподарських організмів.\n\nПри геміклональному успадкуванні певний механізм викликає вибіркову елімінацію певного геному. Розуміння цього механізму може бути цікавим для генної медицини вже зараз: він дозволитиме за потреби вимикати небажані гени.\n\nА чим глибше ми зрозуміємо шлях еволюції жаб, тим цікавішим буде — переконайтеся!\n\nРоботу виконано за спільної підтримки Російського фонду фундаментальних досліджень (проєкт 08-04-01184) та Державного фонду фундаментальних досліджень України (проєкт 09-04-90475).\n\nЛітература\n1. Berger L.// Acta Zoologica Cracoviensia. 1967. №12. Р.123—160.\n2. Tunner H.G.// Zeitschrift fur Zoologische Systematik und Evolutionsforschung. 1974. №12. P.309—314.\n3. Borkin L.J., Korshunov A.V., Lada G.A. et al.// Russian J. of Herpetol. 2004. V.11. №3. P. 194—213.\n4. Боркін Л.Я., Зіненко А.И., Коршунов А.В. та ін. Масова поліплоїдія в гібридогенному комплексі Rana esculenta (Ranidae, Anura, Amphibia) на сході України // Матеріали 1-ї конференції Українського герпетологічного товариства. Київ, 2005. С.23 — 26.\n5. Шабанов Д.А, Зіненко А.И., Коршунов А.В. та ін.// Вісник Харківського національного університету імені В.Н.Каразіна. Сер. бюл. 2006. Вип.3. №729. С.208 — 220.\n6. Borkin L.J., Lada G. A., Litvinchuk S.N. et al // Russian J. of Herpetol. 2006. V.13. №1. P.77—82.\n7. Шабанов Д. А., Коршунов О.В., Кравченко М.О.// Біологія та валеологія. Вип.11. Харків, 2009. С.125 — 164.\n8. Plotner J.Die Westpalaarktichen Wasserfrosche. Bielefeld, 2005.\n9. Боркін Л. Я. , Даревський И.С. // Журнал загальної біології. 1980. Т.41. №4. С.485 — 506.\n10. Plotner J., Uzzell T, Beerli P. et al. // J. of Evol. Biol. 2008. V.21. №3. P.668—681.\n"
}
}{
"translated_text": "Петербурзькі герпетологи Лев Якович Боркін та Ілля Сергійович Даревський близько 30 років тому припустили, що триплойдні гібриди можуть відігравати важливу роль у виникненні нових гібридних видів. Загалом запропонована ними схема така: при схрещуванні диплоїдних видів виникають клонально розмножуючіся триплойдні гібриди. У результаті розмноження триплойдів та їх схрещування з іншими формами виникають тетраплойди, які переходять до нормального розмноження з рекомбінацією геномів [9]. Може, у Північно-Донецькому центрі відбуваються саме такі процеси? Геміклональні триплойди напевні, одиничні тетраплойди — теж. Але чомусь тетраплойди не доживають до статевої зрілості... Еволюція клональних геномів має призвести до якогось нового якості? Чи тетраплойдам для їх розвитку потрібні інші умови, ніж жабам із меншою кількістю хромосомних наборів?\n\nЩе один «пласт» еволюційних змін зелених жаб пов’язаний із порушенням клональної передачі геномів у гібридів. Іноді в клональні геноми потрапляють фрагменти з рекомбінантних геномів іншого виду, і навпаки. Передаючись від гібрида до гібрида у складі генопарадигматичних систем (ГПС), такий частково рекомбінований клональний геном може потрапити до озерної жаби. Так відбувається перенесення спадкової інформації через видові бар’єри!\n\nТакий перенос не обмежується лише ядерною ДНК, а стосується й мітохондріальної (мтДНК). Як відомо, вона знаходиться не в клітинному ядрі, а в мітохондріях та успадковується інакше — виключно за материнською лінією, з цитоплазмою яйцеклітин. Виявилося, що у більшості озерних жаб у Західній Європі взагалі немає власної мтДНК [10]. Вона повністю замінена на мтДНК прудової жаби, отриманою від гібридів. І справа не лише в тому, що мітохондрії прудової жаби виявилися «кращими» за мітохондрії озерної. Щоб отримати їх, західноєвропейські озерні жаби мали мати досить дивну генеалологію. Уявіть: у кожної озерної жаби одна з «бабусь» за материнською лінією мала бути прудовою жабою — представником іншого виду. А ядерні гени у неї — лише свого виду. Отже, ця «бабуся» із прудових жаб схрещувалася з самцем озерної. У результаті мав з’явитися гібрид із мітохондріями та материнським геномом прудової жаби й відцовським геномом — озерної. Стать у жаб успадковується майже так само, як у людей: у самиць — два жіночих геноми, а у самців — один жіночий, другий — чоловічий.\n\nОтже, «дочері» тієї самої «бабусі» прудової жаби мали передати нащадку жіночий геном озерної жаби та схрещуватися з самцем озерної жаби. Самиці, що утворилися від такого схрещування, стали предками за материнською лінією усіх озерних жаб у Західній Європі.\n\nЩо ж у «сухому залишку»? Група тварин із нетипово діючим спадковим апаратом, який еволюціонує нестандартним шляхом. Ця група утворює не популяції, а особливі біосистеми — генопарадигматичні системи (ГПС), що розвиваються за своїми законами. У межах цієї групи спадкова інформація подолає видові бар’єри, і, дуже ймовірно, на наших очах народжується новий тетраплойдний вид.\n\nЯк і навіщо все це вивчати?\n\nВузли такої складності неможливо розплутати за допомогою одного лише інструменту. Просту деталь можна зобразити на кресленні з однієї точки зору, складну вимагає декількох проекцій та перерізів. Шлях еволюції зелених жаб можна зрозуміти, лише об’єднавши зусилля фахівців різних галузей. Ось неповний перелік методів, які використовували ми з колегами* для опису процесів, що відбуваються із зеленою жабою: польові збори та обліки, опис динаміки росту за перерізами кістки, порівняння форми тіла та особливостей розвитку, математичне моделювання розмноження різних форм жаб у ГПС, визначення форм жаб за розміром їх геному, вивчення білокових маркерів, порівняння послідовностей ядерної та мтДНК, визначення характерних особливостей хромосом (імуннофлуоресцентне забарвлення, аналіз «лампових щіток») тощо. Під час роботи доводилося порівнювати жаб різних форм, із різних типів середовищ, ГПС та регіонів. Природно, для розуміння того, що відбувається із жабами, важливими є результати вивчення інших груп тварин, у яких зареєстровано подібні феномени: риб, зелених жаб, хвостатих земноводних, ящірок.\n\n*Ми щиро вдячні Л. Я. Боркіну, Г. А. Ладе, Ю. М. Розанову, М. В. Владимировій, Д. В. Дедуху, А. І. Зіненку, В. В. Клименку, А. В. Коршунову, М. А. Кравченку, Г. А. Мазепі, Є. Є. Усовій, С. Ю. Морозову-Леонову, Й. Плетнеру та багатьом іншим колегам за допомогу та підтримку під час проведення наших досліджень.\n\nДля вирішення настільки складної проблеми не вистачить сили ні одного автора, ні одного наукового колективу. Проблеми, подібні до тієї, яку ми розглядаємо, якраз і здатні об’єднувати наукове співтовариство, пов’язуючи вчених із різних країн.\n\nА навіщо вивчати зелених жаб? Головна для авторів цієї статті відповідь — тому що це дуже цікаво. Цікавість, недовіра, дослідницький азарт, які викликають описані у статті особливості жаб, належать до найприродніших двигунів науки. Але справа не лише в інтересі. Здогадуватися, які «бонуси» можна отримати від вивчення тієї чи іншої проблеми — неблагодарне заняття, але деякі спроби ми все ж зробимо.\n\nГеміклональне успадкування — спосіб прискорення еволюції; вивчення механізмів геміклональності відкриє нові можливості для селекції.\n\nПри геміклональному успадкуванні усі нащадки від схрещування двох особин (кожна з яких передає клональний геном) можуть бути генетично ідентичними. Можливість отримання таких тварин буде корисною для біотехнології та сільського господарства.\n\nОволодіння механізмами, які забезпечують збереження життєздатності гібридів із районів, де неможливе схрещування батьківських видів, допоможе підтримувати життєздатність порід та чистих ліній сільськогосподарських організмів.\n\nПри геміклональному успадкуванні певний механізм викликає вибіркову елімінацію певного геному. Розуміння цього механізму може бути цікавим для генної медицини вже зараз: він дозволитиме за потреби вимикати небажані гени.\n\nА чим глибше ми зрозуміємо шлях еволюції жаб, тим цікавішим буде — переконайтеся!\n\nРоботу виконано за спільної підтримки Російського фонду фундаментальних досліджень (проєкт 08-04-01184) та Державного фонду фундаментальних досліджень України (проєкт 09-04-90475).\n\nЛітература\n1. Berger L.// Acta Zoologica Cracoviensia. 1967. №12. Р.123—160.\n2. Tunner H.G.// Zeitschrift fur Zoologische Systematik und Evolutionsforschung. 1974. №12. P.309—314.\n3. Borkin L.J., Korshunov A.V., Lada G.A. et al.// Russian J. of Herpetol. 2004. V.11. №3. P. 194—213.\n4. Боркін Л.Я., Зіненко А.И., Коршунов А.В. та ін. Масова поліплоїдія в гібридогенному комплексі Rana esculenta (Ranidae, Anura, Amphibia) на сході України // Матеріали 1-ї конференції Українського герпетологічного товариства. Київ, 2005. С.23 — 26.\n5. Шабанов Д.А, Зіненко А.И., Коршунов А.В. та ін.// Вісник Харківського національного університету імені В.Н.Каразіна. Сер. бюл. 2006. Вип.3. №729. С.208 — 220.\n6. Borkin L.J., Lada G. A., Litvinchuk S.N. et al // Russian J. of Herpetol. 2006. V.13. №1. P.77—82.\n7. Шабанов Д. А., Коршунов О.В., Кравченко М.О.// Біологія та валеологія. Вип.11. Харків, 2009. С.125 — 164.\n8. Plotner J.Die Westpalaarktichen Wasserfrosche. Bielefeld, 2005.\n9. Боркін Л. Я. , Даревський И.С. // Журнал загальної біології. 1980. Т.41. №4. С.485 — 506.\n10. Plotner J., Uzzell T, Beerli P. et al. // J. of Evol. Biol. 2008. V.21. №3. P.668—681.\n"
}