
Коли відбір стає неефективним? Колонка в «КомпьютерреOnline» № 47
Коли на пристосованість особини впливає мінливість багатьох генів, а особливо якщо дія одних генів залежить від стану інших, швидкість еволюції за моделлю СТЕ різко знижується.
←
Дмитро Шабанов
→
Парадокс прискорення еволюції
Коли природний відбір стає неефективним?
Чудеса статевого розмноження
Стаття в «КомпьютерреOnline» № 46
Стаття в «КомпьютерреOnline» № 47
Стаття в «КомпьютерреOnline» № 48
Так-так, судячи з відгуків на попередню колонку, вона не переконала багатьох читачів. Я маю на увазі не креаціоністів та їм подібних, які, наче заклинання, повторюють, що «факти й логіка спростовують еволюцію», і демонструють незнання фактів та невміння користуватися логікою. Треба б якось навчитися відповідати таким читачам, щоб вони втратили інтерес до повторення своїх безхитрісних аргументів, але справа не в них.
Одна з проблем розуміння проявилася в тому, що частина читачів вирішила, що якщо я кажу про те, що слони повинні еволюціонувати повільніше за форамініфер згідно з СТЕ (синтетичною теорією еволюції), то швидка еволюція слонів виявляється чимось незрозумілим або дивовижним. Будь ласка, не ототожнюйте СТЕ та еволюційну біологію взагалі! СТЕ з її сімдесятип'ятирічною історією – лише одна з заслужених теорій, хоч і вельми авторитетна. Повірте: сучасна біологія – одна з найдинамічніших наук.
Більш серйозна проблема пов'язана з дуже конкретним мисленням багатьох читачів. Я навів умоглядний приклад із трьома близнюками, які росли в різних умовах, і підштовхнув коментаторів до обговорення життєвих перипетій цих віртуальних людей. А як вплине на еволюцію слонів те, що вони можуть ходити з місця на місце? А як еволюція видів може враховувати те, що чекає на них попереду?..
Зосередженість на тих чи інших життєвих історіях (буває так, а буває й інакше...) заважає аналізувати вплив окремих факторів на еволюцію. Як зробити цей зв'язок наочнішим? Зв'язок вихідних положень і висновків, що випливають з них, у будь-якій теорії не випадковий. Не всі можуть умоглядно уявити, як пов'язана зміна життєздатності якихось категорій особин з їхньою еволюцією. Як зробити для таких читачів мої міркування наочнішими?
Побудувати модель! Моделі, навіть прості, дозволяють зрозуміти, які наслідки випливають із початкового набору властивостей системи, які були враховані при її моделюванні.
Цим я й займуся. Сила СТЕ (одна з причин, завдяки якій ця теорія ще залишається на плаву) у тому, що вона дуже формалізована й спирається на математичний апарат генетики популяцій. Процеси, що протікають відповідно до її передбачень, легко моделювати! Як обґрунтувати, що моя модель відображає передбачення, які мають робитися відповідно до логіки СТЕ? Тут не обмежишся коротким визначенням СТЕ, від одного з її супротивників, як у минулій колонці. А що вважати каноном СТЕ? Оскільки у цієї теорії не було єдиного автора, її творці якось не зібрали воєдино основні положення свого дітища. Найбільш формально описав нову теорію Микола Володимирович Тимофєєв-Ресовський (1900-1981), який працював у той час у Німеччині. У 1939-1940 роках він виділив елементарні структури, матеріал і фактори еволюції. Першим «скелет» (набір постулатів) СТЕ запропонував з полемічною метою її запеклий противник, Олександр Олександрович Любищев (1890-1972). З прихильників СТЕ, здається, це вперше зробив Микола Миколайович Воронцов (1934–2000), один із учнів Тимофєєва-Ресовського.
Воронцов зібрав перелік постулатів СТЕ у 1978–1980 роках. Але й розвиток біології, і власні дослідження (наприклад, ролі хромосомних перебудов у видоутворенні) з часом переконали Воронцова в обмеженості такого канону (і відповідно обмеженості СТЕ). Наприкінці життя він описав, у чому його розуміння біології кінця минулого століття розходилося зі СТЕ. Я мав нахабство викласти на своєму сайті таблицю, де зіставлені постулати СТЕ за Воронцовим і його уточнення до них, зроблені через 20 років. Не забувайте, що й правий стовпець таблиці за Воронцовим – ще не кінець розвитку еволюційної біології...
Отже, формалізований характер СТЕ дозволяє стверджувати, що створена мною модель відображає положення цієї теорії. Модель створена в Excel, якщо хочете, можете завантажити її тут; втім, сподіваюся, що все буде зрозуміло і з самої таблиці. Взагалі, Excel надає чудові можливості для моделювання, роблячи його доступним для людей без математичної та програмістської освіти.
У моделі розглядається популяція, що складається з організмів, які вільно схрещуються. Як це і прийнято в СТЕ, фенотипи (і пристосованість організмів) визначаються їхніми генами. Розглянемо два гени, A і B, кожен з яких представлений двома алелями: A і a для гена A, B і b для гена B. Отже, у популяції можуть існувати дев'ять генотипів: AABB, AABb, AAbb, AaBB, AaBb, Aabb, aaBB, aaBb, aabb.
Занадто просто? І цього мінімуму вистачить, щоб комбінаторика генів виявилася не цілком тривіальною.
Отже, задамо чисельність популяції (позначимо її K – ємність середовища – максимальна кількість особин, яка може мешкати в даних умовах). Задамо початкові частки генотипів (PAAbb; PAaBB і так далі). Що далі?
За СТЕ, джерело нових ознак – нові гени, що виникають у результаті мутацій; поки що не будемо з цим сперечатися. Задамо в моделі частоти, з якими відбуваються переходи одних алелей в інші: Pa→A, PA→a, Pb→B і PB→b.
Залишилося передбачити ту силу, яка повинна змінювати співвідношення алелей у певному напрямку, – добір. Для кожного генотипу вкажемо його пристосованість (fitness): FAAbb; FAaBB та інші.
Модель працює так. Виходячи з розподілу генотипів у популяції, обчислюється склад гамет, які вони будуть продукувати. Для спрощення ми розглядаємо гермафродитні організми, які виробляють і яйцеклітини, і сперматозоїди (один раз у своєму житті). Імовірність зустрічі гамет з будь-якими генотипами однакова (наприклад, так може бути, якщо гамети викидаються на волю випадку у воду, як це роблять багато морських донних тварин).
При обчисленні складу гамет враховується ймовірність мутацій. Комбінації гамет визначають генотипи потомства. Шанси на виживання нащадків залежать від їх пристосованості.
Все, цикл роботи моделі закінчено. Залишилося визначити склад наступного покоління. Округлення частки генотипів у популяції до одиниць особин має ймовірнісний характер. Наприклад, величина 1,4 округлиться до 2 з імовірністю 0,4, і до 1 — з імовірністю 0,6.
Залишилося повторити описаний цикл багато разів (модель побудована для 500 поколінь). Виведемо на графік найцікавіший, з точки зору СТЕ, показник – динаміку співвідношення альтернативних алелей. Компоненти моделі я розмістив на аркуші Excel так, щоб на один екран потрапляли і графік, і віконця для введення вхідних значень (позначення перераховані вище). Скріншоти цього екрана ілюструють подальший виклад.
Ну що, почнемо.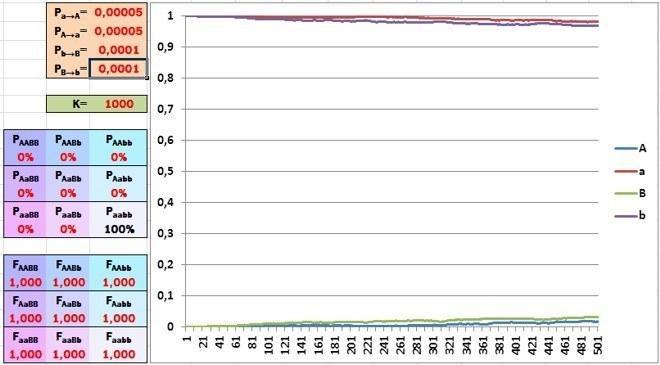
Початковий склад популяції повністю складається з особин aabb. Пристосованість усіх генотипів однакова, склад популяції поступово змінюється внаслідок мутацій (за геном A дещо швидше, ніж за геном B, через різну частоту мутацій).
Якщо в початковому стані популяції по кожному гену представлений лише один алель, мутації поступово підвищують частку альтернативних алелів. А що буде, якщо співвідношення алелів виявиться рівноважним?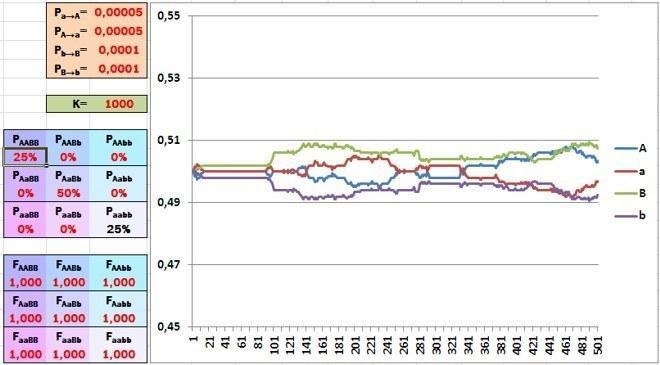
Початкове співвідношення алелей є рівним і коливається під впливом випадкових факторів (мутацій та зсувів під час дискретизації). Зверніть увагу на зміну масштабу по осі ординат.
За відсутності добору частоти алелей почнуть «коливатися» навколо середнього значення. До речі, іноді такі випадкові зсуви можуть призвести до втрати однієї з алелей.
Час «включати» відбір. Нехай особини, які мають «вдалий» генотип, мають на 1 відсоток більші шанси на виживання, ніж усі інші. Вам здається, що така невелика перевага не може зіграти свою роль в еволюції?
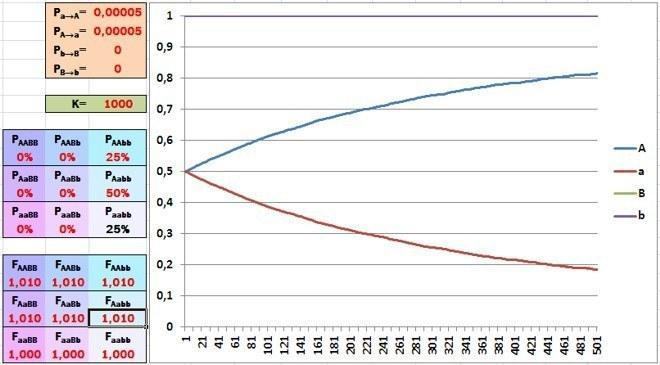
Якщо пристосованість особини визначається єдиним геном, переваги в 1% для носіїв домінантного гена достатньо для досить швидкої зміни співвідношення алелів. А як позначиться на швидкості еволюції рецесивність алеля, якій сприяє відбір?
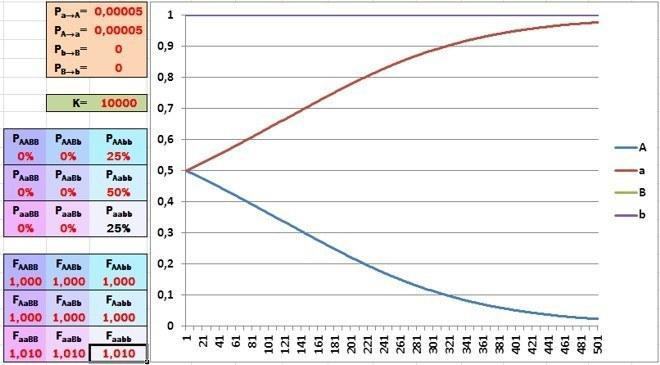
Прискорюється (за умови початкової рівності частот алелів). Дивіться: за час, розглянутий у моделі, алель, якому сприяє відбір, майже витіснив свою альтернативу!
Насправді і популяції, і види відрізняються не за одним, а за безліччю генів. У нашій моделі ми можемо розглянути лише одночасний відбір за двома з них. Як позначиться на швидкості еволюції залучення другого гена з ще однією парою алелей? Щодо гена B теж підтримаємо відбором рецесивний алель (b).
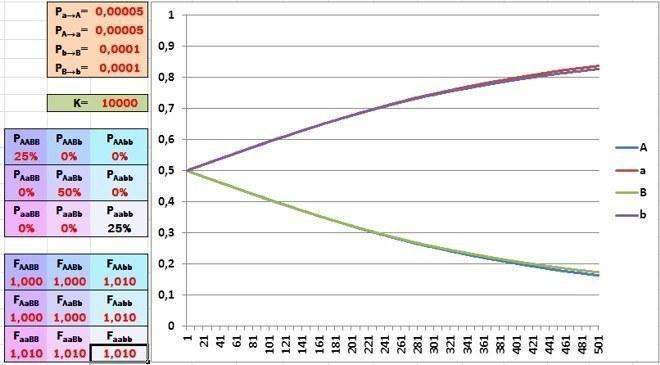
Подивіться, як сповільнився відбір! Пристосованість фенотипу особини залежить одночасно від алелей двох незалежних генів; комбінаторика кожного з них заважає відбору за іншим геном.
На нашій, досить простій моделі це не показати, але якщо на пристосованість особини впливає одночасно 12 генів (по кожному з яких відбувається вибір між двома алелями), відбір стає абсолютно неефективним! Цей феномен називається дилемою Холдейна, і його відкрив не хто-небудь, а один із творців СТЕ Дж.Б.С. Холдейн (1892-1964).
До речі, російська Вікіпедія пише, що дилему Холдейна люблять креаціоністи. Нагадую: це аргумент проти СТЕ, а не проти еволюції як такої!
Як ми переконалися, при рівності частот алелей ефективніше відбувається відбір на користь рецесивного алеля. Відбір на користь рідкісного алеля ефективніший, якщо той домінантний.
Залишилося проілюструвати одну обставину, порушену в минулій колонці. Виставляємо в моделі високу підтримку відбором спочатку рідкісного домінантного алеля – 10%. За 500 поколінь відбору такої сили вистачає, щоб рідкісний алель став переважаючим.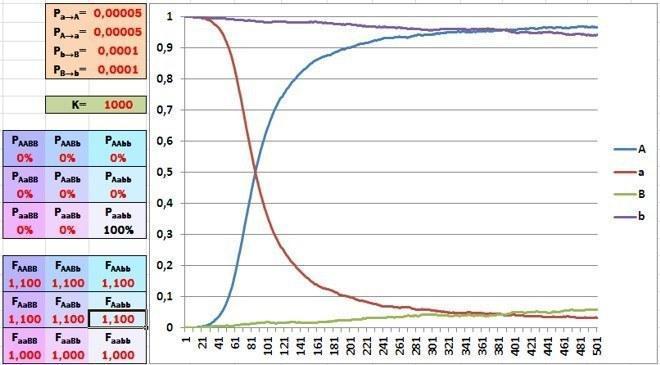
Збільшення частки рідкісного алеля відбувається ефективніше у разі його домінантності.
Але, як ми говорили минулого разу, у складних організмів гени взаємодіють між собою. Розглянемо два домінантні алелі різних генів. Коли вони зустрічаються разом, вони дають значний виграш — аж 20%. Окремо вони дають 1% втрати. Припустимо, у популяції таких генів дуже мало. Чи зможе добір так само ефективно підвищити їхню частку, як у минулому прикладі?
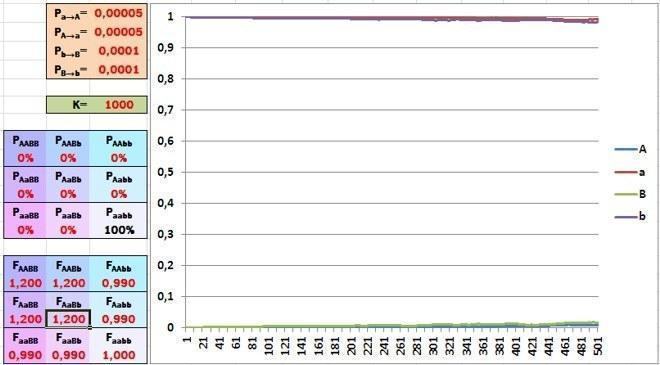
На жаль. Невеликого програшу особин, що мають лише один із взаємодоповнюючих генів, достатньо, щоб цих генів у популяції залишалося небагато. Відбір підтримає щасливчиків із вдалим поєднанням генів, але більшість їхніх нащадків знову отримає ці гени окремо. Популяційна еволюція за таких умов відбувається вкрай повільно.
Чому? Коли фенотип однозначно відображає генотип, виживання більш пристосованих особин ефективно сортує гени. Якщо залежність пристосованості особини від її генотипу ускладнюється, механізм еволюції за СТЕ починає працювати неефективно.
На сьогодні вистачить. Сподіваюся, ви переконалися: у випадках, коли на пристосованість особини впливає мінливість багатьох генів, а тим більше якщо ці гени демонструють складну взаємодію (а не просте підсумовування своїх ефектів), швидкість зміни частот алелей серйозно сповільнюється.
Як узгодити з цим те, що складні організми, які демонструють різноманітні взаємодії своїх генів, еволюціонують швидше за своїх простіших віддалених родичів? З точки зору СТЕ це незрозуміло.
Втім, на СТЕ розвиток науки не зупинився...
←
Дмитро Шабанов
→
Парадокс прискорення еволюції
Коли відбір стає неефективним?
Чудеса статевого розмноження
Колонка в КомпьютерреOnline #46
Колонка в КомпьютерреOnline #47
Колонка в КомпьютерреOnline #48