
Kravchenko ta ін. (2011) Дослідження стійкості геміклональних популяційних систем...
Спільна з математиками стаття. Вивчення моделі ГПС дозволило зробити цікаві, на нашу думку, умозаключення, що стосуються стійкості біосистем. У статті пропонується трактування поняття "стійкість", що відповідає його застосуванню як у математиці, так і в екології. Kravchenko M. A., Shabanov ...
Кравченко М. А., Шабанов Д. А., Владимирова М. В., Жолткевич Г. Н. Дослідження стійкості геміклональних популяційних систем гібридного комплексу зелених жаб за допомогою імітаційного моделювання // Вісник Дніпропетровського університету. Біологія. Екологія. – 2011. – Вип. 19, т. 1. – С. 51–64.
УДК: 004.942:597.851
М. А. Кравченко, Д. А. Шабанов, М. В. Владимирова, Г. Н. Жолткевич
Харківський національний університет імені В. Н. Каразіна
ДОСЛІДЖЕННЯ СТІЙКОСТІ ГЕМІКЛОНАЛЬНИХ ПОПУЛЯЦІЙНИХ СИСТЕМ ГІБРИДНОГО КОМПЛЕКСУ ЗЕЛЕНИХ ЖАБ ЗА ДОПОМОГОЮ ІМІТАЦІЙНОГО МОДЕЛЮВАННЯ
Проаналізовано поняття стійкості в екології та математиці. Обґрунтовується трактування стійкості як здатності системи залишатися в обмеженій зоні фазового простору, що відповідає певному типу систем, що розглядаються. Цей підхід застосовано для опису змін складу геміклональних популяційних систем гібридного комплексу зелених жаб (Pelophylax esculentus complex). Використано імітаційну модель, побудовану на рекурентних різницевих рівняннях у MS Excel. Описано динаміку переходів у частині фазового простору геміклональних популяційних систем зелених жаб. Проаналізовано типологію станів стійкості біосистем.
INVESTIGATION OF THE STABILITY OF HEMICLONAL POPULATION SYSTEMS OF WATER FROGS HYBRIDOGENEOUS COMPLEX BY THE MEANS OF SIMULATION MODELING
M. O. Kravchenko, D. A. Shabanov, M. V. Vladimirova, G. M. Zholtkevych
V. N. Karazin Kharkiv National University
The concept of stability used in ecology and mathematics is analyzed. Stability is interpreted as the ability of the system to remain in a restricted zone of a phase space that corresponds to a certain type of systems. This approach is applied to describe changes in the structure of hemiclonal population systems of hybridogenous complex of water frogs (Pelophylax esculentus complex). Simulation model of these population systems built on a recursive difference equations in MS Excel is used. The dynamics of transitions in the part of phase space of systems hemiclonal populations of water frogs is described. Typology of stabile states of biosystems is analyzed.
Вступ
Для більшості організмів із статевим розмноженням характерне утворення видових популяцій, які входять до складу біотичних спільнот. Особи всередині популяції об’єднані спільністю генофонду і спільно беруть участь у популяційному відтворенні. Популяції у спільноті перебувають у різноманітних взаємовідносинах, від конкуренції до експлуатації та мутуалізму. Частина спільноти, що об’єднує конкуренційні популяції, які використовують один і той самий ресурс, може утворювати гільдії. Перераховані рівні організації біосистем (популяція, гільдія, спільнота) можна вважати типічними. Однак для деяких груп організмів характерні й інші, незвичні рівні організації біосистем. До їх числа належать європейські зелені жаби, Pelophylax esculentus complex.
Назва Pelophylax esculentus (Linnaeus, 1758) належить не виду, а гибриду, який утворюється при схрещуванні двох батьківських видів: Pelophylax lessonae (Camerano, 1882) і Pelophylax ridibundus (Pallas, 1771). Для P. esculentus характерне геміклональне успадкування, при якому в гамети (статеві клітини) переходить або геном (хромосомний набір) P. lessonae, або геном P. ridibundus, а не рекомбінантні геноми, що складаються з суміші батьківських геномів, як це відбувається у більшості організмів із статевим розмноженням [17, 20]. Ми позначаємо геном P. lessonae літерою L, а геном P. ridibundus — R, причому клональність позначаємо дужками: (L) або (R). При схрещуванні з особинами батьківських видів тих гибридів, які передають гетероспецифічний геном (тобто геном іншого батьківського виду), все потомство складається з гибридів: RR × R(L) → R(L). Клональний геном (L) у обох поколіннях однаковий, а рекомбінантні геноми — R — «перебудовуються» заново при передачі з покоління в покоління у особин батьківських видів.
Для зелених жаб характерне утворення популяційних систем, у яких спільно живуть і розмножуються як представники батьківських видів, так і різноманітні геміклональні гибриди. Наприклад, у Північному Донечому центрі різноманіття зелених жаб [17] поряд з особинами з генотипом RR (тобто представниками P. ridibundus) зустрічаються диплоїдні гибриди (L)R, L(R), (L)(R), а також триплоїдні гибриди LLR і LRR. Серед неполовозрілих особин зустрічаються одиночні тетраплоїди LLRR і особини LL (тобто P. lessonae). Важливо, що особини P. lessonae, що виникають від схрещування особин (L)R, виявляються нежиттєздатними. Нежиттєздатність особин батьківських видів, які отримали обидва свої геноми від батьків‑гибридів, реєструвалася і в інших регіонах [20].
Очевидно, що подібні системи, у яких з покоління в покоління передаються як клональні, так і рекомбінантні геноми різних видів, належать до особливої групи біосистем і заслуговують окремої назви. Ми запропонували [16, 17] називати їх геміклональними популяційними системами — ГПС.
ГПС не є популяціями, оскільки вони включають особини різних видів. ГПС не є гільдіями і спільнотами, оскільки об’єднані процесом спільного відтворення. Динаміка ГПС відрізняється від динаміки популяцій, гільдій і спільнот. Одним із наслідків того, що ГПС є особливим рівнем організації біосистем, існування якого було усвідомлене лише нещодавно, є їх значно менша вивченість порівняно з типічними рівнями біосистем.
Метою даної роботи є аналіз різних станів ГПС з точки зору їх стійкості. Попередньою умовою такого аналізу є розгляд сенсу, який вкладається у поняття «стійкість».
Аналіз поняття «стійкість»
Поняття стійкості стало одним із найважливіших екологічних понять з ранніх етапів розвитку цієї науки. Воно було важливим для теорії клімаксу рослинних спільнот Ф. Клементса, що з’явилася на початку ХХ століття [19, цит. за 13]. В. Н. Сукачев [14] визначав стійкість рослинної спільноти як здатність «зберігати свій склад і структуру протягом тривалого часу. Ця стійкість обумовлена здатністю спільноти до самовідновлення».
Незважаючи на інтерес до поняття стійкості в екології, вважати його ясним, на жаль, передчасно. Наведемо два вислови авторитетних авторів. «На жаль, термін «стійкість» надто часто залишався розмитим і невизначеним» [11, с. 324]. «Екологи надзвичайно невігідні у всьому, що стосується стійкості природних систем: які внутрішні механізми спільнот у цьому беруть участь і як вони діють?» [12, с. 394].
Р. Ріклефс проводить аналіз поняття «стійкість». «Стійкість, притаманна даній системі, можна визначити як відношення між змінністю в середовищі і змінністю в самій системі» [12, с. 378], підкреслюючи, що це визначення (фактично говоріння про буферні властивості системи) важко застосувати до популяції і спільноти. Простішим зрозуміти цей підхід на прикладі, що стосується організменного рівня: Ріклефс наводить дані про динаміку кліматичних параметрів і динаміку товщини річних приростів дерев. Виявляється, що з цієї точки зору дерева можуть бути більш чи менш стійкими до зміни зовнішніх умов. Затухання збурень (зниження кількості опадів на 50 % призводить до зниження рослинної продукції на 25 % і до падіння чисельності травоїдних тварин на 10 %) дозволяє говорити про роботу внутрішніх механізмів, що підтримують стійкість екосистеми.
Ріклефс виділяє три типи станів, пов’язаних зі стійкістю: стійка рівновага (збурення компенсуються), нестійка рівновага (збурення посилюються) і нейтральна рівновага (збурення викликають у системі незначні для її функціонування зміни). В кінцевому підсумку, Ріклефс формулює визначення, що характеризує двоїстість розглянутого поняття: «Стійкість – це внутрішньо притаманна системі здатність витримувати зміни, спричинені зовні, або відновлюватися після них» [12, с. 379].
Р. Уіттекер трактує стійкість передусім як буферність. «Ми будемо називати процеси, які зменшують втрати популяції, коли середовище стає менш сприятливим, буферністю популяцій» [15, с. 58].
Понятійне вираження двоїстості феномена стійкості, на яке вказував Р. Ріклефс, дає Ю. Одум [10, с. 66], розглядаючи резистентну стійкість (здатність протистояти збуренням) і еластичну стійкість (здатність відновлюватися після порушення) як прояви стабільності.
М. Бігон і співавтори [3, с. 325‑327, а також 19, p. 576‑577], аналізуючи поняття стійкості, виділяють три пари понять:
— еластичність (resilience) як міра здатності повертатися у вихідний стан і опір (resistance) як міра здатності уникати змін;
— локальна (local) і загальна стійкість (global stability), що різняться за рівнем збурень, які система може компенсувати;
— динамічна крихкість (dynamic fragility) або міцність (robustness) відображають відношення системи до зовнішнього середовища: здатність системи зберігатися в обмеженому наборі зовнішніх умов або в їх широкому діапазоні.
Автори наводять графічні аналогії для пояснення використаних термінів. Як зазначають Бігон і співавтори, вивчення стійкості спільнот здійснюється значною мірою з використанням математичного моделювання.
В. Г. Стороженко [13] надає обширний перелік параметрів, збереження яких є проявом стійкості біогеоценозів. Серед них кількість видів і самі ці види, взаємозв’язки всередині системи, чисельність популяцій, особливості функціонування, придушення шкідливих факторів тощо. Головною задачею аналізу, який проводить В. Г. Стороженко, є пошук критеріїв стійкого лісового спільноти. Автор не розглядає і не класифікує ті стани системи, які далекі від клімаксу.
Як видно з огляду, більшість робіт, присвячених проблемі екологічної стійкості, характеризують умови, за яких ті чи інші спільноти (передусім лісові) виявляються стійкими. Однак, крім стану справжньої стійкості (стійкої рівноваги), як найціннішого стану біосистеми, слід досліджувати й інші її можливі стани. Чи можна розділити сукупність можливих станів системи на типи, що відрізняються з точки зору їх динаміки та стійкості?
Для вирішення цієї проблеми необхідно залучити апарат аналізу стійкості, розвинений у математиці. Розглянутому поняттю «не вдається» і тут: «стійкість – термін, що не має чіткого визначеного змісту» [9, с. 604]. Класичним визначенням поняття «стійкість» є те, що було дано у 1892 р. творцем теорії стійкості руху Олександром Михайловичем Ляпуновим (доцентом, а згодом професором Харківського університету, нині імені В. Н. Каразіна). Стійкість визначався А. М. Ляпуновим з точки зору впливу збурень на рух з певними початковими умовами [8]. Застосування такого підходу до опису динаміки біосистем стикається зі значними складнощами і не зовсім відповідає склалі традиції. На нашу думку, з математичних підходів до розглянутого поняття для екології найкорисніше представлення про стійкість за Ж. Л. Лагранжем, яке є частковим випадком стійкості за А. М. Ляпуновим [5]. З цієї точки зору стійкість є здатністю системи залишатися в обмеженій області свого фазового простору.
Фазовим простором у цьому випадку називається сукупність можливих станів системи, описаних за допомогою набору її змінних стану. Вибір змінних, що задають фазовий простір, дозволяє, наприклад, розглядати стійкість спільноти і з точки зору кількості її видів, і з точки зору постійності її структури. Суть феномена стійкості залишається тією ж, змінюється лише набір координат, у якому досліджується траєкторія змін досліджуваної системи.
Таке поняття стійкості цілком відповідає практиці його застосування в екологічних дослідженнях. Наприклад, хоча Ф. Клементс і не формулював свої погляди саме так, стійкість клімаксної спільноти за Клементсом можна визначити як здатність спільноти зберігати з часом постійний свій тип (на якій би типології не базувалося виділення цього типу).
Подібному підходу близька трактовка стійкості геосистем, що належить А. Д. Арманду [1]. Арманд виділяє стійкість I, тобто незмінність, інертність, а також стійкість II, тобто здатність протистояти збуренням. З стійкістю II пов’язане поняття басейну стійкості. Це «область у абстрактному багатовимірному просторі, побудованому на осях змінних стану, з усіх точок якої перехідний процес приводить систему до одного і того ж рівноважного або гомеостатичного стану».
Арманд виділяє кілька типів станів рівноваги, при яких не відбувається саморозвиток розглянутої системи. Це рівновага стійка, нестійка, байдужа, циклічна, динамічна і підвижна. Перші три типи рівноваги відповідають тим, що виділяє Р. Ріклефс. У випадку циклічної рівноваги система регулярно повертається у один і той же стан. Найскладніше зрозуміти два останні типи. «Динамічна рівновага — стан системи, у якому протилежно спрямовані процеси (привнесення і виведення речовин і енергії, організація і дезорганізація) взаємно компенсуються. Підвижна рівновага — траєкторія системи у просторі станів, за якою система слідує за зміною вхідних змінних (факторів), зберігаючи в кожен момент часу стан рівноваги стійкої» [1]. Коли мова йде про біосистеми, будь‑яке їх рівноважне становище є динамічним. Альтернативою динамічній рівновазі є стійкість I, тобто статичність. На нашу думку, щоб описати різницю між стійкою і підвижною рівновагою, треба розділити впливові на систему фактори на зовнішні і внутрішні. Стійка рівновага — це результат зрівноваження внутрішніх факторів, а підвижна рівновага є наслідком її зміщення зовнішніми факторами.
Отже, поняття стійкості системи виявилося тісно пов’язаним з типологією станів біосистеми. Розділивши простір станів системи на басейни стійкості (між якими розташовані межі стійкості [2]), можна отримати динамічну типологію її станів. При розв’язанні цього завдання для ГПС Pelophylax esculentus complex буде корисною імітаційна модель розглянутих систем.
Імітаційне моделювання ГПС Pelophylax esculentus complex
Для моделювання динаміки ГПС зелених жаб ми розробили сукупність вимог користувача до моделі (URD, User Requirements Definition). Вивчення динаміки ГПС зелених жаб здійснювалося методом імітаційного моделювання. Для верифікації результатів моделювання ми розробили дві диверсні імітаційні моделі. Спільно з А. А. Луциком ми створили мультиагентну імітаційну модель ГПС [4]. Зручнішою у роботі через легкість внесення змін виявилась модель ГПС, основана на рекурентних різницевих рівняннях, реалізована у середовищі Microsoft Office Excel[6]. Одна з версій цієї моделі (названа Batrachometrics-2010) доступна у вільному інтернет‑доступі на сайті Центральної наукової бібліотеки Харківського національного університету імені В. Н. Каразіна за адресою https://dspace.univer.kharkov.ua/handle/123456789/2037. Важливо підкреслити, що обидві реалізації моделі, і мультиагентна, і основана на різницевих рівняннях у MS Excel, генерують якісно схожу динаміку моделюваних систем.
У моделі постулюється існування ГПС, що складається з різних форм жаб. У ГПС передаються геноми, що відрізняються за видовою приналежністю (P. lessonae і P. ridibundus), характером передачі (клональні і рекомбінантні) і статтю (жіночі і чоловічі). Від геному (набору генотипів) і віку особини залежать параметри її життєздатності: максимальна тривалість життя, вік настання статевої зрілості, виживаність за відсутності конкуренції, конкурентоспроможність, привабливість для партнерів, плодовитість і, у випадку самців, ймовірність повторних амплексусів (поєднань пар).
Крок моделі відповідає одному року. Протягом кожного року роботи моделі імітується зниження чисельності кожного вікового класу кожного генотипу під час зими. Ємність середовища обмежує сумарну чисельність або біомасу всіх особин у складі ГПС. Якщо чисельність популяції перевищує ємність середовища, відбувається її конкурентне скорочення. Чисельність кожної групи особин зменшується пропорційно конкурентоспроможності її представників. Парами, що вступають у амплексус, утворюються залежно від привабливості особин кожної групи. У розмноженні беруть участь усі представники статі, яка при спостережуваній у ГПС чисельності здатна вступити в меншу кількість амплексусів, а також відповідна кількість представників іншої статі.Для всіх можливих комбінацій генотипів батьків задаються генотипи виникаючих нащадків. Після імітації виживання в зимівлі, конкурентного скорочення, утворення пар їх розмноження та утворення нащадків модель переходить до наступного кроку (у наступному році), і вся описана послідовність розрахунків повторюється.
Для роботи моделі необхідно задати певні значення параметрів життєздатності для всіх розглянутих форм жаб (детальніше — див. [6]). Далі в даній статті, крім оговорених випадків, використовуються значення, встановлені в Batrachometrics-2010 за замовчуванням.
Фазовий простір і типологія ГПС *Pelophylax esculentus* complex
Той факт, що зелені жаби утворюють популяційні системи, що відрізняються за своїм складом, відомий вже кілька десятиліть. Найпоширеніша типологія популяційних систем жаб була запропонована у 1975 р. за участі відкривача гібридної природи *P. esculentus* Л. Бергера [21]. У цій типології літера L означає присутність у популяційній системі *P. lessonae*, R — присутність *P. ridibundus*, а E — присутність *P. esculentus*. Наявність серед гібридів триплоїдних особин позначається літерою t, присутність виключно самок — літерою f, і виключно самців — літерою m [7]. Наприклад, можна стверджувати, що в заплаві річки Сіверський Донець в околицях біостанції Харківського університету (с. Гайдари Зміївського району Харківської області) мешкає популяційна система REt‑типу [17].
Описаний підхід широко використовується при вивченні поширення різних форм зелених жаб, проте він не враховує того, що особи *P. esculentus* можуть продукувати гамети з різними клональними геномами. Склад гамети визначає і склад майбутнього потомства, і особливості динаміки розглянутої ГПС.
Ми пропонуємо використовувати для характеристики ГПС склад передаваних у ній геномів. У даній роботі ми розглянемо ГПС RE‑типу, де з покоління в покоління передаються рекомбінантні геноми R, і можуть передаватися клональні геноми (L) і (R). Побудуємо двовимірний фазовий простір, осями якого є частки жіночих клональних геномів (XL) і (XR) у загальній кількості геномів (рис. 1). Початку координат у такій системі відповідає популяція R‑типу, що складається з особин *P. ridibundus*. На прямій, що з’єднує точки на осях координат, які відповідають 100 % (R) і 100 % (L), розташовані ГПС E‑типу. Точки вище цієї прямої не визначені, оскільки сума передаваних клональних геномів для них перевищувала б 100 %. Інші точки описаної площини відповідають ГПС RE‑типу.
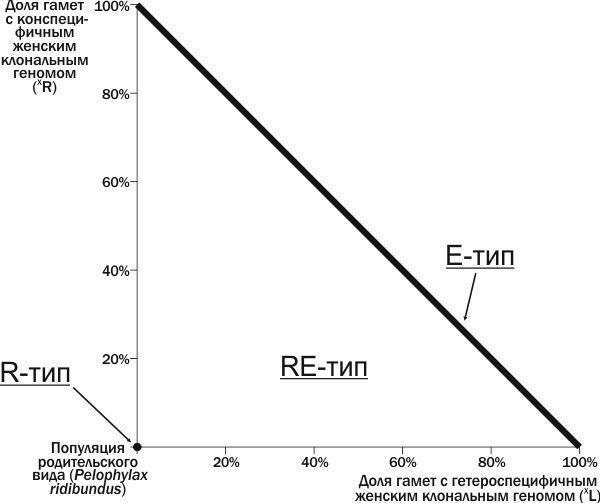
Набір станів ГПС, показаний на рис. 1, — це лише частина можливого різноманіття ГПС зелених жаб. У ГПС з одним батьківським видом можуть передаватися і жіночі, і чоловічі клональні геноми. Окрім RE‑типу ГПС можна розглядати також LE‑тип (набір фазових станів якого симетричний набору станів RE‑типу), а також LER‑тип. Нарешті, у ГПС можуть присутні різні триплоїдні гібриди.
Характеристика поняття стійкості вимагає конкретизації збурень, що впливають на розглянуту систему. Для складу ГПС збуренням є викликане зовнішніми обставинами зміна чисельності представлених у ній форм особин. У розглянутому фазовому просторі такими збуреннями є додавання в ГПС особин *P. esculentus*, що передають жіночі клональні геноми: ♀♀(XL)XR, ♂♂(XL)YR, ♀♀XL(XR) і ♂♂YL(XR).
Трансформації ГПС RE‑типу з жіночими клональними геномами
Популяція R‑типу може перебувати у стаціонарному стані необмежений час. При додаванні ♀♀XL(XR) або ♂♂YL(XR) система через кілька років повертається до попереднього стану. Щодо цього збурення ГПС перебуває у стані стійкої рівноваги.
При потраплянні в популяцію *P. ridibundus* хоча б однієї особини, що передає геном (XL), у ній починаються незворотні перетворення. Усе потомство від схрещування особин батьківського виду і гібридів виявляється гібридним; внаслідок цього частка гібридів від покоління до покоління зростає. По мірі накопичення в ГПС гібридів, що переносять геном (XL), все більша частка схрещувань відбувається між ними і призводить до появи нежиттєздатних особин (XL)(XL). Описаний процес завершується загибеллю ГПС (рис. 2).
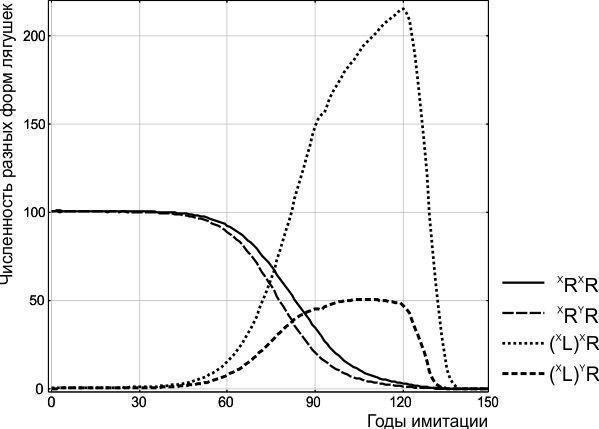
На рис. 2 ілюструється перехід системи з стану нестійкого до додавання геному (XL) рівноваги у стан цілеспрямованої зміни, що характеризується зростанням частки особин (XL)XR і (XL)YR. Результатом цього процесу є загибель усієї системи.
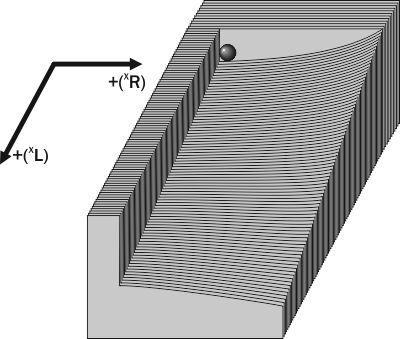
По мірі зростання в ГПС частки геномів (XL) система втрачає стійкість до потрапляння в неї геномів (XR), оскільки за прийнятими в моделі Batrachometrics-2010 умовчаннями особини (XL)(XR) є життєздатними. Щоб пояснити властивості системи на розглянутому ділянці фазового простору, можна, слідом за M. Bigon та співавторами [3], скористатися фізичною аналогією (рис. 3). Початковий стан системи на рис. 3 є стійкою рівновагою щодо додавання геномів (XR) і нестійкою — щодо додавання геномів (XL). В цілому таке становище слід характеризувати як нестійку рівновагу. При потраплянні геномів (XL) система «скочується» у бік збільшення їх частки і втрачає стійкість до потрапляння в неї геномів (XR).
На 90‑му році імітації внесення в показану на рис. 2 ГПС як ♀ XL(XR), так і ♂ YL(XR), не призводить до жодних суттєвих змін. На 100‑му і 110‑му роках і те, і інше впливи призводять до того, що ГПС переходить до іншого стану, у якому передається значна кількість клональних геномів обох видів (рис. 4).
Як видно з рис. 4, ГПС, у якій передаються клональні геноми обох видів, зазнає затухаючих циклічних змін свого складу. При аналізі 500 років імітації можна було б припустити, що ГПС у кінцевому підсумку переходить у стан циклічної рівноваги, при якому вона рухається по певній замкненій траєкторії. На основі аналогій з динаміки інших біосистем можна зазначити, що стану циклічної рівноваги відповідають циклічні сукцесії біогеоценозів або незатухаючі циклічні коливання двох видів у системі «хижак‑жертва» з запізненням реакції на зміни чисельності один одного [11, с. 231].
Проте у випадку розглянутої моделі ГПС рівновага не циклічна, і приблизно до 1000‑го року ГПС досягає точки стійкої рівноваги. Склад нерестового стада в цій точці такий: 10,5 % ♀♀ XRXR; 6,9 % ♂♂ XRYR; 53,9 % ♀♀ (XL)XR; 16,7 % ♂♂ (XL)YR; 8,6 % ♀♀ XL(XR); 3,4 % ♀♀ (XL)(XR). Цей склад ГПС дуже далек від рівності статей: у ньому лише 23,6 % самців і 76,4 % самок. Тим не менш, усі ГПС, що знаходяться всередині петлеподібної частини траєкторії, показаної на рис. 4, саме до цього стану і приходять.
А що відбувається з ГПС, що знаходяться поза показаною на рис. 4 «петлею»? Розглянемо випадок, коли геном (XR) додається лише на 120‑му році (рис. 5).
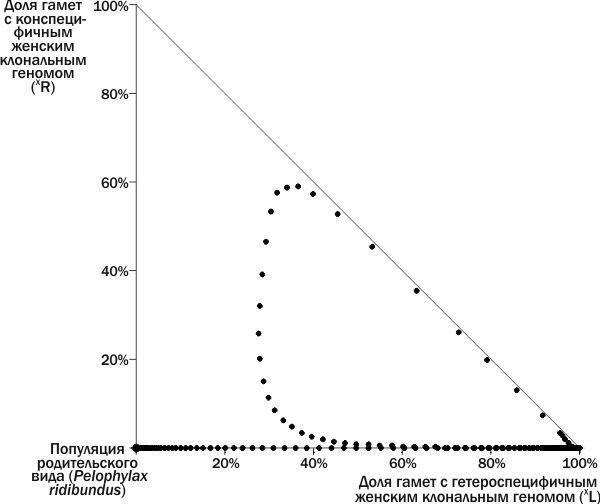
Різноманітність станів ГПС зелених жаб з точки зору їх стійкості
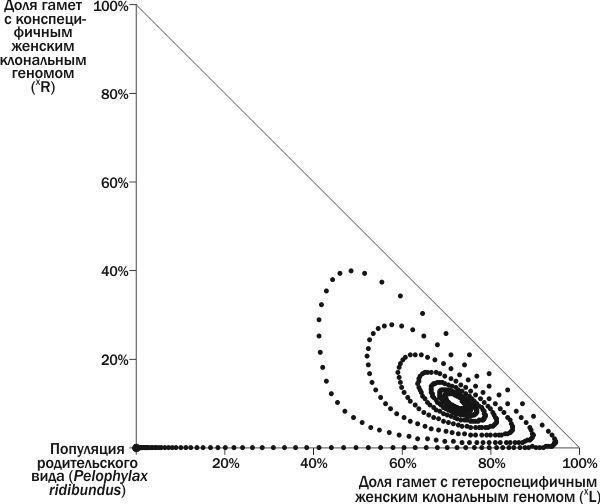
Виходячи з викладеного, ми можемо виділити на вивченому фазовому просторі станів ГПС положення рівноваги та їх басейни (рис. 6). Межа між басейном стійкої рівноваги і басейном загибелі ГПС проходить між петлею, описаною ГПС на рис. 4, і ширшою петлею, описаною ГПС на рис. 5. Розмістивши на невивченій частині фазового простору точки, що відповідають початковим станам ГПС, ми можемо дізнатися, до яких басейнів вони належать. ГПС, що лежать на осі ординат, переходять у початок координат, до популяції *P. ridibundus*. Розвиток усіх інших ГПС, як це ні дивно, закінчується їх загибеллю у точці, що відповідає 100 % (XL).
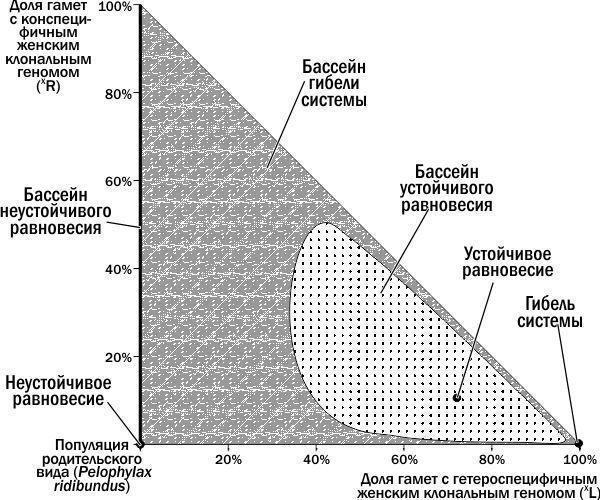
На рис. 6 видно три басейни стійкості (один з них відповідає осі ординат) і три положення рівноваги: нестійке, стійке та загибель системи. Як би парадоксально це не звучало, стан нестійкої рівноваги також має свій басейн. Це пов’язано з тим, що популяція *P. ridibundus*, будучи нестійкою до одних збурень, демонструє стійкість до інших.
Чи може ГПС перебувати в інших станах стійкості, крім показаних на рис. 6? Вище було сказано, що А. Д. Арманд [1] виділяє в окремі категорії стани байдужого та рухомого рівноваги. Як зазначено, рисунки 2, 4, 5 і 6 були отримані при параметрах життєздатності, прийнятих у моделі Batrachometrics-2010 за замовчуванням. При зміні відносної життєздатності різних форм жаб змінюються і властивості фазового простору. Так, при зниженні відносної життєздатності гібридних жаб перевага у їх розмноженні компенсується перевагою особин батьківського виду у життєздатності. При додаванні в популяції *P. ridibundus* ♀(XL)XR (тобто в умовах, аналогічних тим, наслідки яких показані на рис. 2), ГПС переходить у стан рухомого рівноваги. Нагадаємо, що з нашої точки зору про рухоме рівновагу можна говорити за наявності зовнішнього фактора, що протистоїть внутрішнім процесам розвитку системи. У цьому випадку ми розглядаємо знижену життєздатність гібридів як середовий, тобто зовнішній вплив на динаміку ГПС.
Модель Batrachometrics-2010 є детерміністичною. При однакових початкових умовах вона приводить до одного і того ж стану. Стан байдужого рівноваги досяжний при стохастичній, випадковій динаміці ГПС. Для моделювання таких процесів необхідно ввести в модель випадковий компонент.
У наступній версії моделі, Batrachometrics-2011, чисельність кожної групи жаб раз на рік округлюється до цілого числа, причому округлення має ймовірнісний характер (наприклад, величина 1,7 буде округлена до 1 з ймовірністю 0,3, а до 2 — з ймовірністю 0,7). Внаслідок ймовірнісного округлення при кожній новій імітації з одними і тими ж початковими умовами модель може приводити до різних станів.
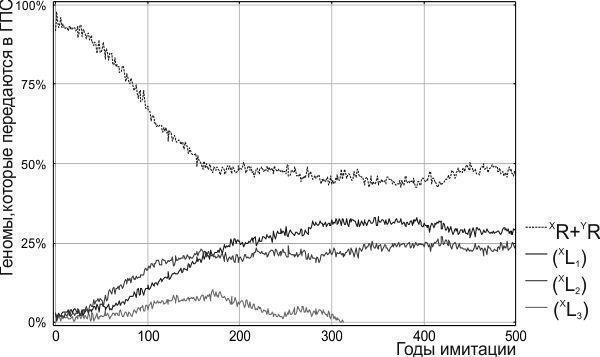
На рис. 7 показана динаміка моделі Batrachometrics-2011 за умов зниженої життєздатності гібридних жаб. З точки зору відношення між представниками батьківського виду і гібридів система переходить у стан рухомого рівноваги: частка рекомбінантних батьківських геномів коливається, кілька разів перевищуючи 50 %. Клональні геноми (XL) на рис. 7 представлені трьома ідентичними за своєю життєздатністю та початковою чисельністю геміклонами. При різних імітаціях з однаковими початковими умовами доля цих геміклонів виявляється різною. На рисунку показаний випадок, коли чисельність одного з них у зв’язку з випадкових причин знижується до нуля, і він зникає з ГПС.
Типи стійкості біосистем
Підводячи підсумки вивченню динаміки ГПС зелених жаб за допомогою імітаційних моделей, можна запропонувати класифікацію типів стійкості біосистем. Ця класифікація в основному відповідає поглядам А. Д. Арманда [1]. Типи стійкості можна розділити на дві групи (рис. 8). До першої належать ті стани, які можуть бути «центрами» (точніше — аттракторами) басейнів стійкості. Це стійка рівновага, циклічна рівновага (приклади якої в даній роботі детально не розглядалися), рухома рівновага та стан загибелі системи. Може здаватися дивним, що загибель системи («негативний» стан) стоїть в одному ряду зі стійкою рівновагою (яка сприймається як «позитивний» стан). Поєднує ці стани те, що, потрапивши в них, система залишається в них назавжди (принаймні, доки її не виведе зовнішній вплив).
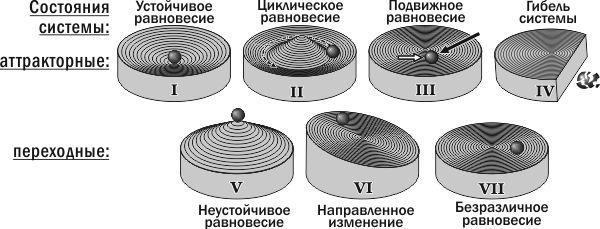
Перехідні стани утворюють басейни аттракторних станів. Це нестійка рівновага, цілеспрямована зміна і байдужа рівновага. Межі між басейнами (межі стійкості) представлені станами нестійкої рівноваги.
Як показують розглянуті в даній статті приклади, описані типи стійкості не абсолютні. Їх виділення залежить від того, які параметри стану системи відстежуються (у якій системі координат будується фазовий простір її станів). Так, досягнуте на рис. 7 ГПС можна трактувати двозначно. З точки зору відношення між рекомбінантними і клональними геномами воно є станом рухомого рівноваги, а з точки зору відношення між різними геміклонами його слід вважати байдужим рівновагою.
Висновки
Геміклональні популяційні системи (ГПС) гібридогенних комплексів є групою біосистем, яка була відкрито зовсім недавно, і їх вивчення лише починається. Це вивчення не лише надає цінний матеріал для пізнання розглянутих груп організмів, а й розширює розуміння загальних властивостей біосистем.
У даній роботі стійкість біосистеми розглядається як здатність залишатися в певній частині її фазового простору. Ця частина фазового простору відповідає певному типу розглянутих систем. Увесь простір станів біосистеми може бути розділений на басейни стійкості, що відповідають певним аттракторним станам. Запропонований підхід придатний для частини фазового простору ГПС зелених жаб RE‑типу, яка вивчалась за допомогою імітаційного моделювання. Побудовані на різних обчислювальних основах імітаційні моделі продемонстрували якісно схожу динаміку досліджуваних систем. Показано вплив на розташування та конфігурацію басейнів стійкості тих умов, у яких розвивається розглянута ГПС.
У даній роботі продемонстровано шість типів станів стійкості біосистеми; сьомий тип відомий з літературних джерел. В цілому ці типи утворюють дві групи: аттракторні стани, «центри» басейнів стійкості, і перехідні стани, які з часом переводять розглянуту систему до тих чи інших аттракторних станів.
Бібліографічні посилання
1. Арманд А. Д. Механізми стійкості геосистем. – М.: Наука, 1992. – 208 с.
2. Арнольд В. І. Теорія катастроф. – М.: Наука, 1990. – 128 с.
3. Бигон М. Екологія. Особини, популяції і спільноти: у 2‑х т. Т. 2./ Бигон М., Харпер Дж., Таунсенд К. // М.: Мир, 1989. – 477 с.
4. Владимирова М. В. Дослідження цілісних властивостей біосистем на прикладі імітаційного моделювання гібридогенних популяційних систем зелених жаб / Владимирова М. В., Жолткевич Г. Н., Луцик А. А., Шабанов Д. А. // Вісник Харківського нац. ун‑ту. Серія «Математичне моделювання. Інформаційні технології. Автоматизовані системи управління» — 2007, № 780, C. 61–70.
5. Калитин Б. С. Якісна теорія стійкості руху динамічних систем. – Мінськ: БГУ, 2002. – 198 с.
6. Кравченко М. А. Моделювання трансформацій геміклональних популяційних систем зелених жаб (*Pelophylax esculentus* complex; Amphibia, Ranidae) за допомогою рекурентних різницевих рівнянь / Кравченко М. А., Шабанов Д. А. // Вісник Харківського національного університету імені В. Н. Каразіна. Серія: біологія. — 2010. – Вип. 12 (№ 920). – С. 70–82.
7. Лада Г. А. Середньоєвропейські зелені жаби (гібридогенний комплекс *Rana esculenta*): введення в проблему // Флора і фауна Чорнозем’я. — Тамбов, 1995. – С. 88—109.
8. Ляпунов А. М. Загальна задача про стійкість руху. М‑Л.: Гостехіздат, 1950. – 472 с.
9. Математичний енциклопедичний словник. – М.: Радянська енциклопедія, 1988. – 847 с.
10. Одум Ю. Екологія: у 2‑х т. Т. 1. – М.: Мир, 1986. – 328 с.
11. Піанка Е. Еволюційна екологія. – М.: Мир, 1981. – 400 с.
12. Ріклефс Р. Основи загальної екології. – М.: Мир, 1979. – 424 с.
13. Стороженко В. Г. Стійкі лісові спільноти. Теорія і експеримент. – Тула: Гриф і К., 2007 – 192 с.
14. Сукачев В. Н. Вибрані праці. – Л. Наука, 1972. – Т. 1, 343 с.
15. Уіттекер Р. Спільноти і екосистеми. – М.: Прогрес, 1980. – 327 с.
16. Шабанов Д. А. Які ж зелені жаби населяють Харківську область? Термінологічний і номенклатурний аспекти проблеми / Шабанов Д. А., Коршунов О. В., Кравченко М. О. // Біологія та валеологія. — Харків: ХДПУ, 2009. — Вип. 11. — С. 164–125.
17. Шабанов Д. А. Зелені жаби: життя без правил або особливий спосіб еволюції? / Шабанов Д. А., Литвинчук С. Н. // Природа. — 2010. — № 3. — С. 29–36.
18. Begon M. Ecology. From individuals to ecosystems. / Begon M., Townsend C. R., Harper J. L. // Blackwell Publishing, 2006. — 738 р.
19. Clements F. E. Plant succession. An analysis of the development of vegetation. – Washington: Carnegie Inst., 1916 – 242 p.
20. Plötner J. Die westpaläarktichen Wasserfrösche. Bielefeld: Laurenti‑Verlag, 2005. – 161 S.
21. Uzzell T. M.{"translated_text":"Електрофоретичні фенотипи Rana ridibunda, Rana lessonae та їх гібридогенний асоційований вид Rana esculenta/ Uzzell T. M., Berger L. // Proc. Acad. nat. Sci. Phila. — 1975. — Vol. 127. — P. 13-24."}