Огіельска, 1994. Ядроподібні тільця в гоніальних клітинах головастиків Rana esculenta (Amphibia, Anura)
Неофіційний робочий переклад надзвичайно цікавої статті. Maria Ogielska. Nucleus-like bodies in gonial cells of Rana esculenta (Amphibia, Anura) tadpoles - a putative way of chromosome elimination. Zoologica Poloniae (1994) 39/3-4: 461-474. За переклад – подяка Насті Бондаревій!
Ядроподібні тельця в гоніальних клітинах головастиків Rana esculenta (Amphibia, Anura) – можливі шляхи елімінації хромосомMaria OgielskaNucleus-like bodies in gonial cells of Rana esculenta (Amphibia, Anura) tadpoles - a putative way of chromosome elimination. Zoologica Poloniae (1994) 39/3-4: 461-474.Department of General Zoology. Zoological Institute, University of Wroclaw, Sienkiewicza 21, 50-335 Wroclaw, PolandРезюмеRana esculenta є природним гибридогенетичним гибридом двох батьківських видів R. lessonae і R. ridibunda. Через особливе гибридогенетичне розмноження обидві статі виробляють гамети, що несуть гаплоїдний набір хромосом одного з батьківських геномів, тоді як інший набір елімується перед мейозом. Щоб виявити шляхи елімінації хромосом, гонади головастиків (і гибридних, і батьківських форм), тобто ті стадії, коли клітини найбільш активно проліферують, вивчали за допомогою електронної, світлової та флуоресцентної мікроскопії. З двох шляхів елімінації хромосом: поступової втрати в процесі мітозів і відпускання від інтерфазного ядра з утворенням ядероподібних тіл (NLB), останній здається більш імовірним.ВступСмачна жаба, Rana esculenta, є гибридом, геном якого складається з гаплоїдних наборів батьківських видів R. lessonae і R. ridibunda (BERGER 1983, GRAF and POLLS-PELAZ 1989). R. esculenta зазвичай зустрічається в популяціях з одним з батьківських видів. Змішані популяції esculenta-lessonae є найпоширенішими і стійкими; їхня генетична система називається E‑L‑системою (UZZELL and BERGER 1975). Індивіди R. esculenta такої системи зазвичай передають у гаметах лише гаплоїдний набір хромосом озерної жаби, тоді як набір ставковий видаляється в процесі гаметогенезу. Гібриди передають з покоління в покоління геном того батьківського виду, який відсутній у популяції. Такий тип розмноження називається гибридогенетичним (SCHULTZ 1969, TUNNER 1974).Наші недавні результати (OGIELSKA and WAGNER 1993, WAGNER and OGIELSKA 1993) показали, що розвиток і диференціація яєчників у R. esculenta є більш тривалим і пізнім процесом порівняно з таковим у батьківських видів (OGIELSKA arid WAGNER 1990, WAGNER and OGIELSKA 1990). Зокрема, відтягування початку мейозу і пролонгована стадія ділення оогониїв є найпомітнішими рисами розвитку яєчників гибридів. Більше того, у цитоплазмі оогониїв гибридів були описані характерні структури, названі ядероподібними тільцями (NLB). NLB не були виявлені ні у батьківських видів, ні у Xenopus laevis (AL‑MUKHTAR and WEBB 1971, COGGINS 1972) і Rana pipiens (MERCHANT‑LAROIS and VILLALPANDO, 1981), єдиних безхвостих амфібій, для яких проводилися ультраструктурні дослідження диференціації гонад головастиків. Метою роботи було виявлення морфологічних маркерів елімінованих хромосом у зароджевій лінії клітин під час ранніх стадій диференціації гонад у головастиків гибридів.Матеріали і методиВикористані в дослідженні головастики були отримані в результаті схрещувань, проведених у лабораторних умовах. Самки були стимульовані шляхом введення гомогенізованого свіжого або замороженого жаб'ячого гіпофізу. Яйцеклітини були штучно запліднені за RUGH (1965). Самці і самки R. esculenta і R. lessonae були спіймані в природних змішаних E‑L‑популяціях поблизу Вроцлава, Польща (Kotowice, Zakrzow, Paniowice). Головастики R. ridibunda і батьківські особини були спіймані поблизу Познани, Польща і люб'язно надані LESZEK BERGER and MARIUSZ RYBACKI (ПОЛЬСЬКА АКАДЕМІЯ НАУК В ПОЗНАНІ).Вивчали потомство наступних схрещувань:7 схрещувань esculenta × lessonae (EL)2 схрещування ridibunda × lessonae (RL)1 схрещування lessonae × ridibunda (LR)і контрольних схрещувань:5 схрещувань lessonae × lessonae (LL)3 схрещування ridibunda × ridibunda (RR).Головастиків вивчали, починаючи зі стадії 25 (голодуючий головастик) до стадії 46 (завершення метаморфози), згідно GOSNER (1960). Головастиків тримали у пластикових контейнерах і годували вареною капустою і риб'ячим кормом. На кожній 2‑5‑й стадії індивідів від кожного схрещування піддавали мікроскопічному аналізу. Також були протестовані кілька ювенільних особин (1 і 1,5 року).Для електронної мікроскопії у головастиків вирізали гонади і фіксували в 2,5 % глутарному альдегіді в 0,1 M фосфатному буфері, pH 7,4, а потім у 1 % OsO₄ або суміші (1:1) 2 % OsO₄ з 1 % ферроціанідом натрію (MCDONALD 1984).Для цитохімічного дослідження ДНК на парафінових зрізах, зафіксованих у глутарному альдегіді, формальдегіді або параформальдегіді, використовували метод фарбування за Фьольгеном і флуоресцентне DAPI‑фарбування. Серії зрізів у парафіні, товщиною 5‑7 мкм, фарбували Fe‑гематоксиліном після фіксації за Ценкером або Хелле і використовували для аналізу мітозу. Рис. 1. Ядроподібні тельця в клітинах гонад головастиків Rana esculenta. NLB – Ядроподібне тельце, N – ядро. А‑С 11000х. D 6600 xРезультатиI. Ядроподібні тельця (NLB)У процесі аналізу ультраструктури клітин зароджевого шляху гонад головастиків у цитоплазмі гоніальних клітин були виявлені сферичні тельця діаметром 1,5‑3,0 мкм. Ці тельця (NLB) були оточені подвійною мембраною і містили матеріал, схожий на ядерний хроматин (рис. 1 A‑D). NLB були виявлені на різній відстані від «основного» ядра. Під електронним мікроскопом ці тельця ніколи не виявлялися у батьківських видів: R. ridibunda і R. lessonae. NLB спостерігалися в інтерфазних гоніальних клітинах (як оогониях, так і сперматогоніях), починаючи зі стадії 28 розвитку головастика, коли починається статева диференціація. NLB не виявлялися в недиференційованих гонадах (стадії 25‑27).Найвища частота NLB зареєстрована в гонадах (як у яєчниках, так і в яєчниках) на стадіях 28‑41, тобто між 30‑м і 80‑м днем після запліднення. У деяких особин кількість NLB була високою на момент фіксації, тоді як у інших – низькою, або ж NLB взагалі відсутні. Під час метаморфози (стадії 42‑46) NLB також виявлялися, але не так часто, як на ранніх стадіях. Вони спостерігалися лише в інтерфазних клітинах як окремі сферичні тельця, розташовані на різних відстанях між ядром і клітинною мембраною. Деякі фотографії, зроблені під електронним мікроскопом, дозволяють реконструювати передбачуваний шлях їх утворення: ядерна мембрана формує випинання, і спочатку внутрішня, а потім і зовнішня мембрани зливаються. Наступним кроком є відокремлення NLB від «основного» ядра (рис. 2 A‑D і II). У кількох випадках атипічні NLB спостерігалися у головастиків на стадіях 35‑36.Фарбування DAPI і за Фьольгеном серії зрізів у парафіні показали наявність ДНК у невеликих сферичних тельцях у цитоплазмі гоніальних клітин (рис. 3 і 4). Кількість таких тіл складала 1‑3 на клітину. У гонадах гибридів зароджеві клітини з тільцями були численними. Неочікувано, такі клітини іноді виявлялися і в гонадах батьківських видів, хоча їх кількість була мала.
Рис. 1. Ядроподібні тельця в клітинах гонад головастиків Rana esculenta. NLB – Ядроподібне тельце, N – ядро. А‑С 11000х. D 6600 xРезультатиI. Ядроподібні тельця (NLB)У процесі аналізу ультраструктури клітин зароджевого шляху гонад головастиків у цитоплазмі гоніальних клітин були виявлені сферичні тельця діаметром 1,5‑3,0 мкм. Ці тельця (NLB) були оточені подвійною мембраною і містили матеріал, схожий на ядерний хроматин (рис. 1 A‑D). NLB були виявлені на різній відстані від «основного» ядра. Під електронним мікроскопом ці тельця ніколи не виявлялися у батьківських видів: R. ridibunda і R. lessonae. NLB спостерігалися в інтерфазних гоніальних клітинах (як оогониях, так і сперматогоніях), починаючи зі стадії 28 розвитку головастика, коли починається статева диференціація. NLB не виявлялися в недиференційованих гонадах (стадії 25‑27).Найвища частота NLB зареєстрована в гонадах (як у яєчниках, так і в яєчниках) на стадіях 28‑41, тобто між 30‑м і 80‑м днем після запліднення. У деяких особин кількість NLB була високою на момент фіксації, тоді як у інших – низькою, або ж NLB взагалі відсутні. Під час метаморфози (стадії 42‑46) NLB також виявлялися, але не так часто, як на ранніх стадіях. Вони спостерігалися лише в інтерфазних клітинах як окремі сферичні тельця, розташовані на різних відстанях між ядром і клітинною мембраною. Деякі фотографії, зроблені під електронним мікроскопом, дозволяють реконструювати передбачуваний шлях їх утворення: ядерна мембрана формує випинання, і спочатку внутрішня, а потім і зовнішня мембрани зливаються. Наступним кроком є відокремлення NLB від «основного» ядра (рис. 2 A‑D і II). У кількох випадках атипічні NLB спостерігалися у головастиків на стадіях 35‑36.Фарбування DAPI і за Фьольгеном серії зрізів у парафіні показали наявність ДНК у невеликих сферичних тельцях у цитоплазмі гоніальних клітин (рис. 3 і 4). Кількість таких тіл складала 1‑3 на клітину. У гонадах гибридів зароджеві клітини з тільцями були численними. Неочікувано, такі клітини іноді виявлялися і в гонадах батьківських видів, хоча їх кількість була мала.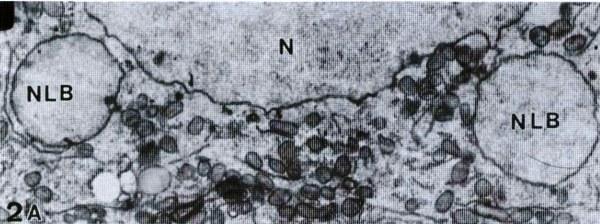
 Рис. 2. Реконструкція передбачуваного шляху формування NLB в інтерфазних ядрах головастиків Rana esculenta на стадіях 29‑31. А – Малий збільшення (2300х) оогонія з двома NLB. В – Формування випинання «основного» ядра (N). Зовнішня мембрана вже злилася. 19200х. С – Збільшений фрагмент одного з NLB з рис. 2 A. Зверніть увагу на стан «основного» ядра (N), позначений стрілкою. 16000х. D – NLB, відокремлене від ядра. Зверніть увагу на подвійну мембрану між ядром і тілом (стрілка). 22400х. Е – Нестандартне NLB, можливо, руйнівне. 16000х.
Рис. 2. Реконструкція передбачуваного шляху формування NLB в інтерфазних ядрах головастиків Rana esculenta на стадіях 29‑31. А – Малий збільшення (2300х) оогонія з двома NLB. В – Формування випинання «основного» ядра (N). Зовнішня мембрана вже злилася. 19200х. С – Збільшений фрагмент одного з NLB з рис. 2 A. Зверніть увагу на стан «основного» ядра (N), позначений стрілкою. 16000х. D – NLB, відокремлене від ядра. Зверніть увагу на подвійну мембрану між ядром і тілом (стрілка). 22400х. Е – Нестандартне NLB, можливо, руйнівне. 16000х. Рис. 3 і 4. Фарбування DAPI яєчника (рис. 3) і яєчка (рис. 4) на стадії 31 головастиків Rana esculenta. У гоніальних клітинах є кругле ядро і більш диспергований хроматин, ніж у соматичних клітинах. Стрілки вказують на маленькі сферичні тельця, що містять ДНК. 700х.2. Мітотичне ділення гоніальних клітин.Одним із можливих шляхів видалення хромосом може бути їх поступова елімінація з клітини в процесі послідовних мітотичних поділів. Така можливість була врахована, і на серіях зрізів у парафіні проаналізовано розподіл хромосом під час ана‑ і телофаз. Інший шлях – елімінація за один крок під час одного мітотичного поділу шляхом формування уніполярного веретена ділення. Ця можливість була підтверджена аналізом форми веретена ділення. Перше мітотичне ділення спостерігали в примордіальних зароджевих клітинах (PGC) зачатків гонад головастиків на пізній стадії 25 (рис. 5). Найвища інтенсивність мітозів у батьківських видів спостерігалася на стадіях 26‑29 (рис. 6, 7, 8), а у гибридів – на стадіях 29‑41. Усі мітотичні веретена, як у батьківських видів, так і у гибридів, були біполярними, уніполярних веретен не виявлено.До стадії 28 (недиференційовані гонади) мітози у гибридів регулярні і жодних відмінностей між батьками і гибридами не виявлено. З початком статевої диференціації гонад гибридів (стадії 28‑29) іноді спостерігаються анафази і телофази з одиничними хромосомами, неправильно розподіленими у веретені (рис. 9 A‑D, 10 A, B).
Рис. 3 і 4. Фарбування DAPI яєчника (рис. 3) і яєчка (рис. 4) на стадії 31 головастиків Rana esculenta. У гоніальних клітинах є кругле ядро і більш диспергований хроматин, ніж у соматичних клітинах. Стрілки вказують на маленькі сферичні тельця, що містять ДНК. 700х.2. Мітотичне ділення гоніальних клітин.Одним із можливих шляхів видалення хромосом може бути їх поступова елімінація з клітини в процесі послідовних мітотичних поділів. Така можливість була врахована, і на серіях зрізів у парафіні проаналізовано розподіл хромосом під час ана‑ і телофаз. Інший шлях – елімінація за один крок під час одного мітотичного поділу шляхом формування уніполярного веретена ділення. Ця можливість була підтверджена аналізом форми веретена ділення. Перше мітотичне ділення спостерігали в примордіальних зароджевих клітинах (PGC) зачатків гонад головастиків на пізній стадії 25 (рис. 5). Найвища інтенсивність мітозів у батьківських видів спостерігалася на стадіях 26‑29 (рис. 6, 7, 8), а у гибридів – на стадіях 29‑41. Усі мітотичні веретена, як у батьківських видів, так і у гибридів, були біполярними, уніполярних веретен не виявлено.До стадії 28 (недиференційовані гонади) мітози у гибридів регулярні і жодних відмінностей між батьками і гибридами не виявлено. З початком статевої диференціації гонад гибридів (стадії 28‑29) іноді спостерігаються анафази і телофази з одиничними хромосомами, неправильно розподіленими у веретені (рис. 9 A‑D, 10 A, B). Рис. 5‑8. 5. Профаза примордіальної зароджевої клітини (PGC) зачатка гонади головастика пізньої стадії 25. Y – жовткова пластинка. 1150х. 6, 7 і 8. Мітози сперматогоній на стадіях 29 і 33 головастиків Rana lessonae. 1150х. Чим пізніше стадія, тим більш численні ненормальні мітози, хоча нормальні завжди становлять більшість. У гибридних самок мітози оогоніїв також спостерігаються після метаморфози, що нетипово для батьківських видів. Після стадії 35 у R. ridibunda і стадії 33 R. lessonae кількість оогоніїв швидко скорочується, і яєчники складаються з усе більшої кількості клітин на стадії диплотени. У гибридів ооцити на фазі диплотени відсутні або їх мало, і більша частина кори яєчника складається з оогоніїв. Мітотичну активність у яєчках після метаморфози не аналізували.
Рис. 5‑8. 5. Профаза примордіальної зароджевої клітини (PGC) зачатка гонади головастика пізньої стадії 25. Y – жовткова пластинка. 1150х. 6, 7 і 8. Мітози сперматогоній на стадіях 29 і 33 головастиків Rana lessonae. 1150х. Чим пізніше стадія, тим більш численні ненормальні мітози, хоча нормальні завжди становлять більшість. У гибридних самок мітози оогоніїв також спостерігаються після метаморфози, що нетипово для батьківських видів. Після стадії 35 у R. ridibunda і стадії 33 R. lessonae кількість оогоніїв швидко скорочується, і яєчники складаються з усе більшої кількості клітин на стадії диплотени. У гибридів ооцити на фазі диплотени відсутні або їх мало, і більша частина кори яєчника складається з оогоніїв. Мітотичну активність у яєчках після метаморфози не аналізували. Рис. 9‑10. 9. A‑D. Неправильна біполярна телофаза оогонія на стадії 20 головастика Rana esculenta, видима на трьох послідовних зрізах (В і С – однакові зрізи при різних фокусуваннях). 1150х; 10. A і B. Біполярний мітоз оогонія на стадії 30 головастика Rana esculenta з однією хромосомою поза метафазною пластинкою. 1150х.ОбговоренняМодель гибридогенезу, запропонована TUNNER (1974), хоча й добре пояснює генотипи потомства R. esculenta, не була підтверджена цитологічними даними про елімінацію одного з хромосомних наборів. У 1973 р. GONTHER описав сперматогенез у дорослих самців R. esculenta, але не виявив жодних ознак елімінації хромосом. З іншого боку, HEPPICH et al. (1982) and BUCCI et al. (1990) показали, що проліферуючі сперматогонії дорослих самців R. esculenta містять лише хромосоми R. ridibunda. Однак VINOGRADOV et al. (1990) виявили кілька дорослих самців R. esculenta, у чиїх сперматогоніях були геноми обох батьківських видів. Подібні результати отримали для дорослих самок. Експериментальний гіногенез (розвиток яйцеклітини без чоловічого пронуклеуса) ооцитів R. esculenta привів до отримання ridibunda потомства (GRAF and MULLER, 1979), а цитоплазма ооцитів R. esculenta на стадії диплотени містила продукти трансляції генів R. ridibunda (CHEN and STUM‑ZOLLINGER, 1986). Нещодавно BUCCI et al. (1990) описали біваленти ооцитів R. esculenta на стадії диплотени, що складаються з двох ridibunda хромосом.Вищенаведені дані показують, що елімінація хромосом R. lessonae має відбуватися на ранніх стадіях розвитку і до мейозу, як було вперше запропоновано UZZELL et al. (1980). TIJNNER and HEPPICH (1981) і TUNNFR and HEPPICH‑TUNNER (1991) розробили техніку подвійного фарбування fluorescence lioechst‑Actinomycin D, яка дозволяє розрізняти ridibunda і lessonae хромосоми. Вони досліджували самок R. esculenta незадовго до метаморфози, коли ще є ділячі оогонії, і описали три класи оогоніїв: диплоїдні ridibunda, гаплоїдні ridibunda і анеуплоїдні з домінуючими ridibunda хромосомами. Схоже, що елімінація lessonae хромосом була поступовою і, можливо, розпочалася на ранніх стадіях розвитку. Детальні дослідження розвитку яєчника у R. ridibunda, R. lessonae і R. esculenta показали, що інтенсивна проліферація оогоніїв у батьківських видів закінчується перед метаморфозою (стадія 42), тоді як у гибридів цей процес спостерігався і після метаморфози (OGIELSKA and WAGNER, 1990, 1993; WAGNER and OGIELSKA 1990, 1993). Більше того, трансформація оогоніїв в ооцити затримувалася і у більшості ювенільних самок не було ооцитів на стадії диплотени. Ці дані відповідають частотності NLB: вони виявлялися на стадіях 29‑41, але також до і після метаморфози, тобто на тих стадіях, для яких TUNNER and HEPPICH (1981) і TUNNER and HEPPICH‑TUNNER (1991) описали поступову втрату lessonae хромосом клітинами зароджевої лінії. Ці дані дозволяють припустити, що NLB можуть бути носіями видалених хромосом.NLB також виявляли в сперматогоніях під час ранніх стадій розвитку яєчка у головастиків. Дослідження сперматогенезу проводили HEPPICH et al. (1982) and Bucci et al. (1990), але вони стосувалися лише дорослих самців. Генотип сперматогоній дорослих особин був виключно ridibunda. Це дозволяє припустити, що lessonae хромосоми під час сперматогенезу видаляються лише до або одразу після метаморфози. Однак, немає докладних робіт щодо сперматогенезу і розвитку яєчок.Під час аналізу електронною мікроскопією NLB виявляли лише у головастиків R. esculenta і ніколи у батьківських видів. З іншого боку, цитохімічний аналіз за допомогою світлової мікроскопії показав наявність невеликих сферичних тіл, що містять ДНК, у гоніальних клітинах батьків. Якщо NLB і DAPI‑місткі тельця є одними і тими ж структурами, ми повинні розглядати можливість того, що батьківські види також здатні викидати хромосоми з зароджевих клітин, хоча цей феномен є рідкісним. Це може розглядатися як еволюційна основа для гибридогенезу у гибридів: гибридогенез можливий, оскільки батьківські види мають цитологічні механізми видалення хромосом. Головне питання в цьому дослідженні, що потребує відповіді, – яким чином хромосоми видаляються з зароджевої лінії. Для обговорення прийнято два можливих шляхи, схематично показані на рис. 12:1.Елімінація хромосом відбувається під час інтерфази. Можливий механізм – різновид ферментативного деграду хроматину, який потім видаляється з інтерфазного ядра у вигляді NLB. Ця гіпотеза підтверджується схожістю NLB і структур, спостережуваних під час апоптозу (програмованої клітинної смерті) (см. CLARKE, 1990). Такий ймовірний шлях також був згаданий у роботах VINOGRADOV et al. (1990).
Рис. 9‑10. 9. A‑D. Неправильна біполярна телофаза оогонія на стадії 20 головастика Rana esculenta, видима на трьох послідовних зрізах (В і С – однакові зрізи при різних фокусуваннях). 1150х; 10. A і B. Біполярний мітоз оогонія на стадії 30 головастика Rana esculenta з однією хромосомою поза метафазною пластинкою. 1150х.ОбговоренняМодель гибридогенезу, запропонована TUNNER (1974), хоча й добре пояснює генотипи потомства R. esculenta, не була підтверджена цитологічними даними про елімінацію одного з хромосомних наборів. У 1973 р. GONTHER описав сперматогенез у дорослих самців R. esculenta, але не виявив жодних ознак елімінації хромосом. З іншого боку, HEPPICH et al. (1982) and BUCCI et al. (1990) показали, що проліферуючі сперматогонії дорослих самців R. esculenta містять лише хромосоми R. ridibunda. Однак VINOGRADOV et al. (1990) виявили кілька дорослих самців R. esculenta, у чиїх сперматогоніях були геноми обох батьківських видів. Подібні результати отримали для дорослих самок. Експериментальний гіногенез (розвиток яйцеклітини без чоловічого пронуклеуса) ооцитів R. esculenta привів до отримання ridibunda потомства (GRAF and MULLER, 1979), а цитоплазма ооцитів R. esculenta на стадії диплотени містила продукти трансляції генів R. ridibunda (CHEN and STUM‑ZOLLINGER, 1986). Нещодавно BUCCI et al. (1990) описали біваленти ооцитів R. esculenta на стадії диплотени, що складаються з двох ridibunda хромосом.Вищенаведені дані показують, що елімінація хромосом R. lessonae має відбуватися на ранніх стадіях розвитку і до мейозу, як було вперше запропоновано UZZELL et al. (1980). TIJNNER and HEPPICH (1981) і TUNNFR and HEPPICH‑TUNNER (1991) розробили техніку подвійного фарбування fluorescence lioechst‑Actinomycin D, яка дозволяє розрізняти ridibunda і lessonae хромосоми. Вони досліджували самок R. esculenta незадовго до метаморфози, коли ще є ділячі оогонії, і описали три класи оогоніїв: диплоїдні ridibunda, гаплоїдні ridibunda і анеуплоїдні з домінуючими ridibunda хромосомами. Схоже, що елімінація lessonae хромосом була поступовою і, можливо, розпочалася на ранніх стадіях розвитку. Детальні дослідження розвитку яєчника у R. ridibunda, R. lessonae і R. esculenta показали, що інтенсивна проліферація оогоніїв у батьківських видів закінчується перед метаморфозою (стадія 42), тоді як у гибридів цей процес спостерігався і після метаморфози (OGIELSKA and WAGNER, 1990, 1993; WAGNER and OGIELSKA 1990, 1993). Більше того, трансформація оогоніїв в ооцити затримувалася і у більшості ювенільних самок не було ооцитів на стадії диплотени. Ці дані відповідають частотності NLB: вони виявлялися на стадіях 29‑41, але також до і після метаморфози, тобто на тих стадіях, для яких TUNNER and HEPPICH (1981) і TUNNER and HEPPICH‑TUNNER (1991) описали поступову втрату lessonae хромосом клітинами зароджевої лінії. Ці дані дозволяють припустити, що NLB можуть бути носіями видалених хромосом.NLB також виявляли в сперматогоніях під час ранніх стадій розвитку яєчка у головастиків. Дослідження сперматогенезу проводили HEPPICH et al. (1982) and Bucci et al. (1990), але вони стосувалися лише дорослих самців. Генотип сперматогоній дорослих особин був виключно ridibunda. Це дозволяє припустити, що lessonae хромосоми під час сперматогенезу видаляються лише до або одразу після метаморфози. Однак, немає докладних робіт щодо сперматогенезу і розвитку яєчок.Під час аналізу електронною мікроскопією NLB виявляли лише у головастиків R. esculenta і ніколи у батьківських видів. З іншого боку, цитохімічний аналіз за допомогою світлової мікроскопії показав наявність невеликих сферичних тіл, що містять ДНК, у гоніальних клітинах батьків. Якщо NLB і DAPI‑місткі тельця є одними і тими ж структурами, ми повинні розглядати можливість того, що батьківські види також здатні викидати хромосоми з зароджевих клітин, хоча цей феномен є рідкісним. Це може розглядатися як еволюційна основа для гибридогенезу у гибридів: гибридогенез можливий, оскільки батьківські види мають цитологічні механізми видалення хромосом. Головне питання в цьому дослідженні, що потребує відповіді, – яким чином хромосоми видаляються з зароджевої лінії. Для обговорення прийнято два можливих шляхи, схематично показані на рис. 12:1.Елімінація хромосом відбувається під час інтерфази. Можливий механізм – різновид ферментативного деграду хроматину, який потім видаляється з інтерфазного ядра у вигляді NLB. Ця гіпотеза підтверджується схожістю NLB і структур, спостережуваних під час апоптозу (програмованої клітинної смерті) (см. CLARKE, 1990). Такий ймовірний шлях також був згаданий у роботах VINOGRADOV et al. (1990). Рис. 11. Формування NLB шляхом відпускання від інтерфазного ядра гоніальної клітини. См. також рис. 2 A‑D2.Елімінація хромосом відбувається під час мітозу. Ця гіпотеза підтверджується численними мета‑, ана‑ і телофазами, у яких окремі хромосоми неправильно розподілялися у веретені ділення. Такі одиничні хромосоми (або хромосомні фрагменти) можуть формувати власну ядерну оболонку, що характерно для хроматину, що з'являється в цитоплазмі, наприклад, у випадку ДНК вірусів, введеної в цитоплазму яйцеклітини (FORBES et al., 1983, SHIOKAWA et al. 1987), що проникає в яйцеклітку пронуклеусом сперматозоїда, який реконструює свою оболонку перед злиттям (LONGO and ANDERSON 1968) або у випадку кариомерів, коли окремі хромосоми формують власну ядерну оболонку під час телофази (ITO ET AL., 1981). Те саме відбувається і у риб Hydrolagus colliei, для яких описано зменшення кількості хромосом у сперматогоніях (STANLEY et al. 1984). У гибридних R. esculenta це може бути, однак, вторинним ефектом зменшення кількості інтерфазних хромосом при подальших клітинних діленнях. Складно сказати, яка з двох гіпотез вірна. Однак, згідно морфологічних особливостей, перша здається більш імовірною.
Рис. 11. Формування NLB шляхом відпускання від інтерфазного ядра гоніальної клітини. См. також рис. 2 A‑D2.Елімінація хромосом відбувається під час мітозу. Ця гіпотеза підтверджується численними мета‑, ана‑ і телофазами, у яких окремі хромосоми неправильно розподілялися у веретені ділення. Такі одиничні хромосоми (або хромосомні фрагменти) можуть формувати власну ядерну оболонку, що характерно для хроматину, що з'являється в цитоплазмі, наприклад, у випадку ДНК вірусів, введеної в цитоплазму яйцеклітини (FORBES et al., 1983, SHIOKAWA et al. 1987), що проникає в яйцеклітку пронуклеусом сперматозоїда, який реконструює свою оболонку перед злиттям (LONGO and ANDERSON 1968) або у випадку кариомерів, коли окремі хромосоми формують власну ядерну оболонку під час телофази (ITO ET AL., 1981). Те саме відбувається і у риб Hydrolagus colliei, для яких описано зменшення кількості хромосом у сперматогоніях (STANLEY et al. 1984). У гибридних R. esculenta це може бути, однак, вторинним ефектом зменшення кількості інтерфазних хромосом при подальших клітинних діленнях. Складно сказати, яка з двох гіпотез вірна. Однак, згідно морфологічних особливостей, перша здається більш імовірною. Рис. 12. Два передбачувані шляхи формування NLBЯким би не був шлях видалення хромосом (зменшення кількості), цей феномен загалом рідкісний для клітин зароджевої лінії. Серед хребетних він відомий у риб Poeciliopsis lucida‑moncicha (CIMINO, 1972) і Hydrolagus colliei (STANLEY et al. 1984). P. lucida‑monacha формують диплоїдні популяції самок, для яких описано гибридогенез (SCHULTZ, 1969). Під час мітозів оогоніїв спостерігали одноразову елімінацію батьківських хромосом. Її цитологічний механізм включає формування уніполярного веретена ділення, до якого приєднані лише материнські хромосоми, тоді як батьківський набір хромосом залишається в цитоплазмі. Під час сперматогенезу H. colliei частина хромосом видаляється в метафазі I. Компактна маса видалених хромосом не включається в ядро, а формує окремі тільця (CDC – chromatin diminution body), оточені подвійною клітинною мембраною. CDC залишається в цитоплазмі однієї з дочірніх клітин до стадії сперматиди, а потім переміщується в цитоплазму клітини Сертолі, де і руйнується.R. esculenta третій відомий вид хребетних, для яких описано зменшення кількості хромосом. І R. esculenta, і P. lucida‑monacha розмножуються шляхом гибридогенезу, а H. colliei не має специфічного типу розмноження, і роль CDC залишається невідомою.У безхребетних також описано зменшення кількості хромосом у зароджевій лінії. Під час сперматогенезу Sciara coprofila батьківські хромосоми елімуються в цитоплазму в метафазі I. Вони залишаються гетерохроматизованими, а потім видаляються з клітини. Тільки материнські хромосоми залишаються прикріпленими до мітотичного уніполярного веретена (METZ, 1933, ABBOT et al. 1981). Інший приклад – Metaseiulus occidentalis (Acarina), у яких видаляються під час сперматогенезу батьківські хромосоми спочатку гетехроматинізуються всередині ядра, а потім елімуються з клітини (NELSON‑REES et al., 1980).Нещодавно гібридогенез і елімінація геному були описані для деяких популяцій палочників Bacillus rossius‑grandii henazzi з Сицилії (TINTI and SCALI, 1992). У цьому випадку один із батьківських наборів хромосом піддається гетерохроматинізації перед метафазою I, потім переміщується в полярне тіло і елімінуються. Ні формування уніполярного веретена, ні гетерохроматинізація хромосом не спостерігалися у досліджуваних R. esculenta. Найймовірніший шлях видалення одного набору хромосом – формування NLB під час інтерфази. Однак механізм цього феномена залишається не розкритим.REFERENCESABBOT, A.G., HEES, J.E. and GERBI. S.A., 1981: Spermatogenesis in Sciara coprophilq. I. Chromosome orientation on the monopolar spindle of meiosis I. Chromosoma, 83: 1-18.AL-MUKHTAR, K.K. and WLBB, A.C., 1971: An ultrastructural study of primordial germ cells, oogonia and early oocytes in Xenopus laevis. J. Embryol. exp. Morph., 26: 195-217.BERGER, L., 1983: Western palearctic frogs (Amphibia, Ranidac): systematics, genetics and population composition. Experientia, 39: 127-130.BUCCU S., RAGGHIANTI, M., MANCINO, G., BERGBR, L., HOTZ, H. and UZZELL, Т., 1990: Lampbrush and mitotic chromosomes of the hemiclonally reproducing hybrid Rana esculenta and its parental species. J. exp. Zool., 255: 37-56. CHEN, P.S. and STUMM‑ZOLLINGER, E., 1986: Patterns of protein synthesis in oocytes and early embryos of Rana esculenta complex. Roux's Arch. Dev. Biol., 195: 1-9.CIMINO, M.C., 1972: Egg‑production, polyploidization and evolution in a diploid all‑female fish of the genus Poeciliopsis. Evolution, 26: 294-306. CLARKE. P.G.H., 1990: Developmental cell death: morphological diversity and multiple mechanisms. Anat. Embryol., 181:195-213.COGGINS, L.W., 1973: An ultrastruciural and autoradiographic study of early oogenesis in the toad Xenopus laevis. J. Cell Science, 12: 71-93.FORBES, D.J., KIRSCHNER, M.W. and NEWPORT, J.W., 1983: Spontaneous formation of nucleus‑like structures around bacteriophage DNA microinjected into Xenopus eggs. Cell, 34: 12-23.GOSNER, K.L., I960: A siplified table for staging anuran embryos with notes of their identification. Herpetologia, 16: 183-190.GRAF, J‑D. and MULLER, W.P., 1979: Experimental gynogenesis provides evidence of hybridogenetic reproduction in the Rana esculenta complex. Experientia, 35: 1574-1576.GRAF, J‑D. and POLLS‑PELAZ, M., 1989: Evolutionary genetics of the Rana esculenta complex. In: Evolution ane ecology of unisexual vertebrates. Ed. R.M. Dawley and J.P. Bogart, Bulletin 466, New York State Museum, Albany, N. York, USA, pp. 289-301.GUNTHER, R., 1973: IJber die verwandtschaftlichen Beziehungen zwischcn den europaisschen Grunfroschen und dem bastardcharakter von Rana esculenta L. (Anura). Zool. Anz., Leipzig, 190: 250-285.HEPPICH, S., TUNNER, H.G. and GRFJLHUBFR, J., 1982: Premeiotic chromosome doubling after genome elimination during spermatogenesis of the species hybrid Rana esculenta. Theor. Appl. Genet., 61: 101-104.ITO, S., DAN, K. and GOODENOUGH, D., 1981: Ultrastructure and lP‑thymidyne incorporation into chromosome vesicles in sea urchin embryos. Chromosoma, 83: 441- 453.LONGO, F.J. and ANDERSON, E., 1968: The fine structure of pronuclear development and fusion in the sea urchin Arbacia punctata. J. Cell Biol., 39: 339-368.MCDONALD, K., 1984: Osmium ferrocyanide fixation improves microfilament preservation and membrane visualization in a variety of animal cell types. J. Ultr. Res., 86: 107-108.MERCUAND‑LAROIS, П., VILLALPANDO, I., 1981: Ultrastructural events during early gonadal development in Rana pipiens and Xenopus laevis. Anat. Rec., 199: 349- 360.METZ, C.W., 1933: Monocentric mitosis with segregation of chromosomes in Sciara and its bearing on the mechanisms of mitosis. Biol. Bull., 64: 333-347.NELSON‑REES, W.A., HOY. M.A. and ROUSH, R.T., 1980: Heterochroma‑tinization, chromatin elimination and haploidization in the parahaploid mite Metaseiulus occidentalis (Nesbitt) (Acarina: Phytoseiidae). Chromosoma, 77: 263-276.OGIELSKA, M. and WAGNER, E., 1990: Oogenesis and development of the ovary in European green frog, Rana ridibunda (Pallas). I. Tadpole stages until metamorphosis. Zool. Jb. Anat., 120: 211-221.OGIELSKA, M. and WAGNER, E., 1993: Oogenesis and ovary development in the natural hybridogenetic water frog, Rana esculenta L. 1. Tadpole stages until metamorphosis. Zool. Jb. Physiola., 97: 349-368.SHIOKAWA, K.. TASHIRO, K., YAMANA, K. and SAMESHIMA, M., 1987: Electron microscopic studies of giant nucleus‑like structures formed by DNA introduced into the cytoplasm of Xenopus laevis fertilized eggs and embryos. Cell Diff., 20: 253-261.STANLEY, H.P., KASINSKY, H.E. and BOLS, N.C., 1984: Meiotic chromatine diminution in a vertebrate, the holocephalan fish Hydrolagus colliei (Chondrichthies, Holocephali). Tissue and Cell, 16: 203-215.RUGH, R., 1965: Experimental Embryology. Burges, Minneapolis.SCHULTZ, JR., 1969: Hybridization, unisexuality and polyploidy in the teleost Poeciliopsis (Poeciliidae) and other vertebrates. Amer. Natur., 103: 605-619.TINTI, F. and SCALI, V., 1992: Genome exclusion and gametic DAPI‑DNA content in the hybridogenetic Bacillus rossius‑grandii benazzi complex (Insecta Phasmatodea). Mol. Rep. Dev., 33: 235-242.TUNNER, H.G., 1974: Die klonale Struktur einer Wasserfroschpopulation. Z. Zool. Evol. Forsch., 12: 309-314.TUNNER, H.G. and HEPPICH, S., 1981: Premeiotic genome exclusion during oogenesis in the common edible frog, Rana esculenta. Naturwissenschaften, 68: 207- 208.TUNNER. H.G. and HEPPICH‑TUNNER, S., 1991: Genome exclusion and two stratergies of chromosome duplication in oogenesis of a hybrid frog. Naturwissenschaften, 78: 32-34.UZZELL, Т., HOTZ, H. and BERGER, L. 1980: Genome exclusion in gametogenesis by an interspecific Rana hybrid: evidence from electrophoresis of individual oocytes. J. exp. Zool., 259: 214-251.VINOGRADOV, A.E., BORKIN, L.J. and ROSANOV, J.M., 1990: Genome elimination in triploid and diploid Rana esculenta males: cytological evidence from DNA flow cytometry. Genome, 33:619-626.WAGNER, E. and OGIELSKA, M., 1990: Oogenesis and development of the ovary in European green frog, Rana ridibunda (Pallas). 2. Juvenile stages until adults. Zool. Jb. Anat., 120: 223-321.WAGNER, E. and OGIELSKA, M., 1993: Oogenesis and ovary development in the natural hybridogenetic water frog, Rana esculenta L. 2. After metamorphosis until adults. Zool. Jb. Physiol., 97: 349-368.UZZELL, T.M. and BERGER, L., 1975: Electrophoretic phenotypes of Rana ridibunda, Rana lessonae and their hybridogenetic associate, Rana esculenta. Proc. Acad. Nat. Sci. Philadelphia, 127: 13-24.
Рис. 12. Два передбачувані шляхи формування NLBЯким би не був шлях видалення хромосом (зменшення кількості), цей феномен загалом рідкісний для клітин зароджевої лінії. Серед хребетних він відомий у риб Poeciliopsis lucida‑moncicha (CIMINO, 1972) і Hydrolagus colliei (STANLEY et al. 1984). P. lucida‑monacha формують диплоїдні популяції самок, для яких описано гибридогенез (SCHULTZ, 1969). Під час мітозів оогоніїв спостерігали одноразову елімінацію батьківських хромосом. Її цитологічний механізм включає формування уніполярного веретена ділення, до якого приєднані лише материнські хромосоми, тоді як батьківський набір хромосом залишається в цитоплазмі. Під час сперматогенезу H. colliei частина хромосом видаляється в метафазі I. Компактна маса видалених хромосом не включається в ядро, а формує окремі тільця (CDC – chromatin diminution body), оточені подвійною клітинною мембраною. CDC залишається в цитоплазмі однієї з дочірніх клітин до стадії сперматиди, а потім переміщується в цитоплазму клітини Сертолі, де і руйнується.R. esculenta третій відомий вид хребетних, для яких описано зменшення кількості хромосом. І R. esculenta, і P. lucida‑monacha розмножуються шляхом гибридогенезу, а H. colliei не має специфічного типу розмноження, і роль CDC залишається невідомою.У безхребетних також описано зменшення кількості хромосом у зароджевій лінії. Під час сперматогенезу Sciara coprofila батьківські хромосоми елімуються в цитоплазму в метафазі I. Вони залишаються гетерохроматизованими, а потім видаляються з клітини. Тільки материнські хромосоми залишаються прикріпленими до мітотичного уніполярного веретена (METZ, 1933, ABBOT et al. 1981). Інший приклад – Metaseiulus occidentalis (Acarina), у яких видаляються під час сперматогенезу батьківські хромосоми спочатку гетехроматинізуються всередині ядра, а потім елімуються з клітини (NELSON‑REES et al., 1980).Нещодавно гібридогенез і елімінація геному були описані для деяких популяцій палочників Bacillus rossius‑grandii henazzi з Сицилії (TINTI and SCALI, 1992). У цьому випадку один із батьківських наборів хромосом піддається гетерохроматинізації перед метафазою I, потім переміщується в полярне тіло і елімінуються. Ні формування уніполярного веретена, ні гетерохроматинізація хромосом не спостерігалися у досліджуваних R. esculenta. Найймовірніший шлях видалення одного набору хромосом – формування NLB під час інтерфази. Однак механізм цього феномена залишається не розкритим.REFERENCESABBOT, A.G., HEES, J.E. and GERBI. S.A., 1981: Spermatogenesis in Sciara coprophilq. I. Chromosome orientation on the monopolar spindle of meiosis I. Chromosoma, 83: 1-18.AL-MUKHTAR, K.K. and WLBB, A.C., 1971: An ultrastructural study of primordial germ cells, oogonia and early oocytes in Xenopus laevis. J. Embryol. exp. Morph., 26: 195-217.BERGER, L., 1983: Western palearctic frogs (Amphibia, Ranidac): systematics, genetics and population composition. Experientia, 39: 127-130.BUCCU S., RAGGHIANTI, M., MANCINO, G., BERGBR, L., HOTZ, H. and UZZELL, Т., 1990: Lampbrush and mitotic chromosomes of the hemiclonally reproducing hybrid Rana esculenta and its parental species. J. exp. Zool., 255: 37-56. CHEN, P.S. and STUMM‑ZOLLINGER, E., 1986: Patterns of protein synthesis in oocytes and early embryos of Rana esculenta complex. Roux's Arch. Dev. Biol., 195: 1-9.CIMINO, M.C., 1972: Egg‑production, polyploidization and evolution in a diploid all‑female fish of the genus Poeciliopsis. Evolution, 26: 294-306. CLARKE. P.G.H., 1990: Developmental cell death: morphological diversity and multiple mechanisms. Anat. Embryol., 181:195-213.COGGINS, L.W., 1973: An ultrastruciural and autoradiographic study of early oogenesis in the toad Xenopus laevis. J. Cell Science, 12: 71-93.FORBES, D.J., KIRSCHNER, M.W. and NEWPORT, J.W., 1983: Spontaneous formation of nucleus‑like structures around bacteriophage DNA microinjected into Xenopus eggs. Cell, 34: 12-23.GOSNER, K.L., I960: A siplified table for staging anuran embryos with notes of their identification. Herpetologia, 16: 183-190.GRAF, J‑D. and MULLER, W.P., 1979: Experimental gynogenesis provides evidence of hybridogenetic reproduction in the Rana esculenta complex. Experientia, 35: 1574-1576.GRAF, J‑D. and POLLS‑PELAZ, M., 1989: Evolutionary genetics of the Rana esculenta complex. In: Evolution ane ecology of unisexual vertebrates. Ed. R.M. Dawley and J.P. Bogart, Bulletin 466, New York State Museum, Albany, N. York, USA, pp. 289-301.GUNTHER, R., 1973: IJber die verwandtschaftlichen Beziehungen zwischcn den europaisschen Grunfroschen und dem bastardcharakter von Rana esculenta L. (Anura). Zool. Anz., Leipzig, 190: 250-285.HEPPICH, S., TUNNER, H.G. and GRFJLHUBFR, J., 1982: Premeiotic chromosome doubling after genome elimination during spermatogenesis of the species hybrid Rana esculenta. Theor. Appl. Genet., 61: 101-104.ITO, S., DAN, K. and GOODENOUGH, D., 1981: Ultrastructure and lP‑thymidyne incorporation into chromosome vesicles in sea urchin embryos. Chromosoma, 83: 441- 453.LONGO, F.J. and ANDERSON, E., 1968: The fine structure of pronuclear development and fusion in the sea urchin Arbacia punctata. J. Cell Biol., 39: 339-368.MCDONALD, K., 1984: Osmium ferrocyanide fixation improves microfilament preservation and membrane visualization in a variety of animal cell types. J. Ultr. Res., 86: 107-108.MERCUAND‑LAROIS, П., VILLALPANDO, I., 1981: Ultrastructural events during early gonadal development in Rana pipiens and Xenopus laevis. Anat. Rec., 199: 349- 360.METZ, C.W., 1933: Monocentric mitosis with segregation of chromosomes in Sciara and its bearing on the mechanisms of mitosis. Biol. Bull., 64: 333-347.NELSON‑REES, W.A., HOY. M.A. and ROUSH, R.T., 1980: Heterochroma‑tinization, chromatin elimination and haploidization in the parahaploid mite Metaseiulus occidentalis (Nesbitt) (Acarina: Phytoseiidae). Chromosoma, 77: 263-276.OGIELSKA, M. and WAGNER, E., 1990: Oogenesis and development of the ovary in European green frog, Rana ridibunda (Pallas). I. Tadpole stages until metamorphosis. Zool. Jb. Anat., 120: 211-221.OGIELSKA, M. and WAGNER, E., 1993: Oogenesis and ovary development in the natural hybridogenetic water frog, Rana esculenta L. 1. Tadpole stages until metamorphosis. Zool. Jb. Physiola., 97: 349-368.SHIOKAWA, K.. TASHIRO, K., YAMANA, K. and SAMESHIMA, M., 1987: Electron microscopic studies of giant nucleus‑like structures formed by DNA introduced into the cytoplasm of Xenopus laevis fertilized eggs and embryos. Cell Diff., 20: 253-261.STANLEY, H.P., KASINSKY, H.E. and BOLS, N.C., 1984: Meiotic chromatine diminution in a vertebrate, the holocephalan fish Hydrolagus colliei (Chondrichthies, Holocephali). Tissue and Cell, 16: 203-215.RUGH, R., 1965: Experimental Embryology. Burges, Minneapolis.SCHULTZ, JR., 1969: Hybridization, unisexuality and polyploidy in the teleost Poeciliopsis (Poeciliidae) and other vertebrates. Amer. Natur., 103: 605-619.TINTI, F. and SCALI, V., 1992: Genome exclusion and gametic DAPI‑DNA content in the hybridogenetic Bacillus rossius‑grandii benazzi complex (Insecta Phasmatodea). Mol. Rep. Dev., 33: 235-242.TUNNER, H.G., 1974: Die klonale Struktur einer Wasserfroschpopulation. Z. Zool. Evol. Forsch., 12: 309-314.TUNNER, H.G. and HEPPICH, S., 1981: Premeiotic genome exclusion during oogenesis in the common edible frog, Rana esculenta. Naturwissenschaften, 68: 207- 208.TUNNER. H.G. and HEPPICH‑TUNNER, S., 1991: Genome exclusion and two stratergies of chromosome duplication in oogenesis of a hybrid frog. Naturwissenschaften, 78: 32-34.UZZELL, Т., HOTZ, H. and BERGER, L. 1980: Genome exclusion in gametogenesis by an interspecific Rana hybrid: evidence from electrophoresis of individual oocytes. J. exp. Zool., 259: 214-251.VINOGRADOV, A.E., BORKIN, L.J. and ROSANOV, J.M., 1990: Genome elimination in triploid and diploid Rana esculenta males: cytological evidence from DNA flow cytometry. Genome, 33:619-626.WAGNER, E. and OGIELSKA, M., 1990: Oogenesis and development of the ovary in European green frog, Rana ridibunda (Pallas). 2. Juvenile stages until adults. Zool. Jb. Anat., 120: 223-321.WAGNER, E. and OGIELSKA, M., 1993: Oogenesis and ovary development in the natural hybridogenetic water frog, Rana esculenta L. 2. After metamorphosis until adults. Zool. Jb. Physiol., 97: 349-368.UZZELL, T.M. and BERGER, L., 1975: Electrophoretic phenotypes of Rana ridibunda, Rana lessonae and their hybridogenetic associate, Rana esculenta. Proc. Acad. Nat. Sci. Philadelphia, 127: 13-24.