
Rasnytsyn, 2008. Теоретичні основи еволюційної біології - 09
1.2.3. ТАКСОНОМІЧНА НОМЕНКЛАТУРА. 1.2.3.1. Таксономічно незалежні принципи. 1.2.3.2. Таксономічно залежні принципи. 1.2.3.3. Таксони, що порушують принципи номенклатури. 1.3. ВИСНОВКИ. Література
1.2.2.2. Фенетика
1.2.2.3. Філетика
А.П.Расніцин. Теоретичні основи еволюційної біології // В.В.Жеріхін, А.Г.Пономаренко, А.П.Расніцин. Вступ до палеоентомології. М.: КМК. 2008. 371 с
1.2.2.3. Філетика (продовження)
1.2.3. ТАКСОНОМІЧНА НОМЕНКЛАТУРА
1.2.3.1. Таксономічно незалежні принципи
1.2.3.2. Таксономічно залежні принципи
1.2.3.2. Таксономічно залежні принципи (продовження)
1.2.3.3. Таксони, що порушують принципи номенклатури
1.2.3. ТАКСОНОМІЧНА НОМЕНКЛАТУРА
Номенклатура — це сукупність домовленостей про те, як створювати та використовувати назви таксонів. Зазвичай її сприймають як область ускладнених деталей і простих до очевидності базових принципів. Це помилково, по‑перше, оскільки ще далеко не всі діючі принципи біологічної номенклатури усвідомлені і сформульовані явно. Деякі з них згадані в кодексах (см. МКЗН, 1988, 2000; МКБН, 2000), але не як базові принципи, а багатьох принципів там просто немає. По‑друге, деякі з базових принципів не є чистими угодами, а відображають певні таксономічні концепції або являють собою наслідки певних еволюційних гіпотез. Ліннеїв принцип біномінальної номенклатури цілком конвенційний і тому легко поєднується з будь‑якою таксономічною концепцією, що визнає ієрархію родів і видів. Інша справа принцип типу, як це докладно обговорюється далі. Розрізнення цих двох типів принципів вважається корисним, і надалі вони будуть називатися таксономічно незалежними та таксономічно залежними принципами відповідно (табл. 4).
Таблиця 4. Принципи номенклатури
Таксономічно незалежні
1
невтручання у прийняття таксономічних рішень поза межами обмежень, накладених таксономічно залежними принципами
2
обмеженої сфери застосування
3
гомонімії
4
пріоритету
5
стандартизованих, ранг‑специфічних назв
6
верховного пріоритету Міжнародної Комісії з номенклатури
Таксономічно залежні
7
типу (немає розумних причин відхилити)
8
ієрархії (немає розумних причин відхилити)
9
сінонімії (застосування до паратаксонів обмежене рамками відповідної приватної системи)
1.2.3.1. Таксономічно незалежні принципи
Таксономічно незалежні принципи зазвичай прості, не вимагають великих коментарів і тут розглядаються першими.
1. У діючому Кодексі зоологічної номенклатури сказано, що він «утримується від втручання у таксономічні судження, які не можуть підлягати регулюванню» (МКЗН, 2000: 24). Це твердження не цілком справедливе через існування таксономічно залежних принципів, які не дозволяють приймати певні таксономічні рішення. Тому формулювання потребує доповнення словами «поза межами обмежень, накладених Кодексом».
2. У зоології область застосування принципів номенклатури обмежена надпопуляційними групами від підвиду до надродини (детальніше см. МКЗН, 2000, ст. 1.2). Виключення старших таксонів зі сфери дії кодексу вважається помилковим (Rasnitsyn, 1982, 1989, 1991, 2002b). Коротко мої міркування можна викласти так.
Стандартизація, забезпечувана кодексами, довела довгою практикою свою благотворну дію на всю біологію. Відмова від її поширення на старші таксони навряд чи пов’язана з їх глибинною специфікою. Серйозних вказівок на таку специфіку не відомо, а численні спроби стандартизувати нетипізовані назви старших таксонів (закінчення -ptera у назв орденів комах, -formes у орденів птахів, -ida у багатьох назв вимерлих орденів різних груп безхребетних) знімають усі сумніви в необхідності такої уніфікації. Справа, звичайно, не в уніфікації, а в типіфікації назв старших таксонів. Під типіфікацією (точніше — сквозною типіфікацією) розуміється процедура, що забезпечує жорстке зв’язування кожної назви кожного таксона з певним підпорядкованим таксоном у його складі, зверху вниз по всій ієрархії до типового екземпляра виду (або підвиду). Типізуються саме назви, а не самі таксони: коли говорять про голотип виду або типовий вид роду, мають на увазі тип назви (імені).
Принцип типу, що відповідає самій природі таксона (см. нижче), є головним досягненням біологічної номенклатури: він забезпечує стійкість вживання і упорядкованість зміни назви таксона при найрізноманітніших змінах його складу (кордонів) і інтерпретації (зміна рангу, діагностичних ознак тощо). Перші спроби типіфікації старших таксонів налічують вже понад два століття (Laicharting, 1781), і розуміння її необхідності також зростає. Така точка зору перемогла в ботаніці (МКБН, 1985) і знаходить певну підтримку в зоології (Rodendorf, 1977; Starobogatov, 1991; Klyuge, 2000). Справа в тому, що нетипізована назва віднесена до таксона як до цілого і практична лише доти, доки інтерпретація самого таксона залишається стійкою. Інакше виникають проблеми з використанням назви, що призводить до безладу і перейменувань. Однак введення типізованих назв старших таксонів і, відповідно, їх перейменування зачіпає інтереси численних користувачів, яким мало цікаві проблеми систематики високих рівнів, але лякає перспектива вчити нові слова. Для тих, хто працює зі старшими таксонами, ситуація виявляється важкою: таксономічні та філогенетичні помилки і неточності консервуются нетипізованими назвами і накопичуються. Чим довше це триває, тим важче буде переходити до типіфікації старших таксонів. Але переходити все одно доведеться, і нащадки не скажуть нам спасибі за наш егоїстичний консерватизм.
3. Ідентичні назви різних таксонів не повинні використовуватись як дійсні назви (МКЗН, 2000, ст. 52). Область застосування принципу гомонімії довільно обмежена так, щоб не зачіпати випадки гомонімії, по‑перше, між назвами видів у різних родах, по‑друге, між таксонами рослин і тварин, по‑третє, з таксонами рангу вище надродини і між такими таксонами.
4. Вибір між конкуруючими назвами (сінонімами або гомонімами) має здійснюватись на користь тієї назви, що була запропонована раніше (принцип пріоритету; МКЗН, 2000, ст. 23).
5. Принцип стандартизованої та ранг‑специфічної форми назв таксонів. Цей принцип становить основу для численних правил мови, граматики і синтаксису назв, і включає принцип біномінальної номенклатури (МКЗН, 2000, ст. 4‑6, 11, 25‑34).
6. Останній серед таксономічно незалежних принципів — принцип верховенства Міжнародної Комісії з номенклатури, яка може приймати рішення на порушення будь‑яких положень Кодексу, крім тих, що стосуються самої Комісії (МКЗН, 2000, ст. 78).
1.2.3.2. Таксономічно залежні принципи
Таксономічно залежних принципів три:
7. Принцип типу. Це найважливіший із таксономічно залежних принципів, і на його прикладі добре видно ті обмеження, які таксономічна концепція може накладати на номенклатуру. У Кодексі 1988 р. цей принцип сформульовано так: «Номенклатурний тип служить об’єктивним еталоном, за допомогою якого застосування назви визначається незалежно від можливих змін кордонів таксона» (МКЗН, 1988, ст. 61а; формулювання МКЗН 2000, ст. 61.1, здається менш вдалим: «Фіксація номенклатурного типу номінального таксона забезпечує об’єктивний еталон для застосування назви цього таксона»). Іншими словами, таксон не може бути введений у систему іншим чином, ніж, в кінцевому підсумку, посиланням на тип, що несе його назву. Щоб оцінити сенс цього твердження, розглянемо альтернативні можливості.
Перше, що спадає на думку, — це можливість введення таксона в систему посиланням на його ознаки. Це означає, що ознаки таксона виконують визначальну, а не лише діагностичну функцію, а сам таксон відповідає поняттю «клас» (Ghiselin, 1974, 1987). Існують різні можливості такого введення таксона в систему, найпростіший з яких — комбінаторна система, тобто багатовимірна матриця, кожна клітинка якої відповідає певній комбінації ознак і є місцем таксона, що має таку комбінацію. Версію такої матриці представляє політомічна таблиця, іноді використана систематиками, тобто прямокутна матриця з рядками, що представляють таксони, і колонками, що відповідають ознакам. У результаті кожна клітинка відображає стан однієї конкретної ознаки в певному таксоні. Ця форма системи дійсно проста і іноді дуже корисна, але не відповідає завданням загальної системи організмів. Вона не має гнучкості: ми не можемо виправити її локально, щоб, наприклад, краще узгодити її з ознаками конкретного таксона, оскільки введення нової, переозначення або видалення старої ознаки змушує переглядати характеристики всіх таксонів. Тому навіть як засіб ідентифікації комбінативна система використовується рідко.
Більш гнучка система, у якій таксони визначаються ранжованими ознаками. У звичайній визначальній таблиці перша використана ознака має найвищий ранг, оскільки ділить визначену сукупність на найбільші групи. Подальші ознаки забезпечують її подальше дроблення і мають нижчий ранг. Така система дійсно зручна, хоча й не настільки хороша, як загальна система організмів, через очевидно довільне ранжування ознак. Проте в дещо модифікованій формі (з виділенням невеликої кількості високорангованих ознак, достатніх для формування системи) цей підхід був дуже популярний у систематиків. У К. Ліннея (Linnaeus, 1751), наприклад, близьку роль відігравали ознаки фруктіфікацій рослин. У найчіткішій формі цей підхід сформулював A.A. Любищев (1923, 1966). Любищев закликав до пошуку небагатьох найвищих рангованих ознак (параметрів), що детермінують розподіл усіх інших ознак, — так само, як ядерний заряд визначає властивості атомів і, відповідно, розташування елементів у таблиці Менделєєва. У результаті очікувалося створення параметричної системи організмів, яка дозволить прогнозувати всі важливі властивості відповідних таксонів за цими ключовими ознаками (параметрами). Завдання Любищева здавалося безнадійним, і йому самому не вдалося його розв’язати. Проте рішення в певному сенсі було знайдено, але зовсім не там, де він його шукав. Іронічно, це сталося в галузі, яку сам Любищев як переконаний антиселекціоніст вважав хибною.
Рішенням виявилася кладистична система з її центральним твердженням, що ознаки організму і, відповідно, його положення в системі найкраще визначаються єдиною ознакою — родством. Родство виявилося любищевським параметром за визначенням. Суть і можливості кладистичної системи обговорені вище, тут я лише хочу звернути увагу на той факт, що ця система не потребує типіфікації своїх таксонів, оскільки посилання на значення параметра (характер родинних відносин) достатнє для введення таксона в систему.
Дійсно, «назви синонімічні, якщо вони вказують на клад [гілку], що відходить від одного і того ж предка» (de Queiroz, Gauthier, 1990: 307). Логічний висновок з цього твердження, саме відмова від принципу типу (не кажучи вже про ліннеївську систему стандартних таксономічних рангів і інші дрібниці), був зроблений Ф. Плейелем з співавторами (Sundberg, Pleijel, 1994; Plejel, Rouse, 2002), і відповідний кодекс філогенетичної номенклатури (Філокод) знаходиться у процесі розробки. Передбачається, що згідно з цим кодексом роль типу, що фіксує застосування назви таксона, виконуватиме або вузол кладограми, або її міжвузол, або апоморфія. Автори заявляють про великі переваги філогенетичної номенклатури щодо як простоти застосування, так і її стабільності (Plejel, Rouse, 2002). Мені важко в це повірити через високу ступінь гіпотетичності кладограми, яка є системою гіпотез про ненаблюдану історію спостережуваних ознак (см. вище). Проте на цьому місці розумніше зупинитися і дати часу працювати. Воно покаже, наскільки життєздатна таксономічна номенклатура, побудована на кладистичних принципах.
Логічною альтернативою класу як групи, визначеної її ознаками, є індивід, і пропозиція вважати таксон індивідом (Ghiselin, 1974, 1987 і наведена там бібліографія) цілком природна. Маючи властивості індивіда, таксон може вводитися в систему остенсивно, тобто прямим вказанням (в тому числі і посиланням на його назву). Для цього його цілісність у просторі і часі повинна бути достатньо високою, щоб дозволяти таксону народжуватись і вмирати, але не розпадатися на частини, кожна з яких могла б претендувати на успадкування імені таксона. Припустимо, людина на шляху від немовляти до старості не зберегла жодного спільного атома речовини і жодної спільної ознаки, але при усіх її метаморфозах завжди можна простежити, що вона є однією і тією ж особою.
Індивід настільки цілісний, що використання його імені не викликає проблем. Можна просто вказати пальцем — ось воно, і не деталізувати, до чого саме відноситься ім’я — до голови, серця, вуха чи чогось іншого. До всього одразу і до кожної деталі окремо. Саме тут проходить межа між остенсивним методом і принципом типу: якщо будь‑яка частина індивіда рівно є носієм його назви, спеціальні правила вибору і використання номенклатурного типу, що слугує для введення таксона в систему, виявляються зайвими.
Якщо метод типу не застосовний до індивіда, то таксон не є індивідом. Принаймні він не типічний індивід (см., наприклад, коментарі до статті Ghiselin, 1981), тому проблема потребує подальшого обговорення. Парадигматичний приклад індивіда, організм, цілісний завдяки безперервній взаємодії його частин. Те саме стверджувалося щодо виду, цілісність якого забезпечується обміном генами (концепція біологічного виду Майрова). Однак ця модель має обмежене застосування, хоча б тому, що «Концепція біологічного виду застосовна лише до того, що я назвав ‘непротяжною ситуацією’, коли популяції виду перебувають у реальному [репродуктивному] контакті» (Mayr, 1988: 301‑302). Насправді проблем тут, як ми вже бачили, ще більше (см. розділ про онтологію еволюції).
Проте дихотомія клас — індивід не вичерпує всіх можливостей. Таксон володіє рисами обох цих крайностей. У ролі класу він має ознаки, про що свідчить наявність діагнозу, і членами (на противагу Гіселіну, я є не лише частиною виду Homo sapiens, а й його екземпляром, тобто прикладом, представником). У ролі індивіда таксон має частини (популяції щодо виду) і, що важливіше, здатність розвиватися (еволюціонувати), не втрачаючи своєї індивідуальності. Розвиваючись, таксон зберігає свою цілісність у часі, у багатовимірному просторі ознак і, в тій чи іншій мірі, у географічному просторі. Така цілісність дозволяє ідентифікувати таксон як індивіда, тобто за його іменем. У результаті таксон, маючи ознаки і класу, і індивіда, заповнює логічний простір між цими поняттями і перетворює клас і індивіда у два полюси єдиного спектра.
Отже, таксон — це щось проміжне між класом і індивідом. Він достатньо цілісний, щоб, змінюючись, зберігати ім’я як засіб розпізнавання, але недостатньо цілісний, щоб просте «ось воно» було достатньо для його розпізнавання. Недостатньо цілісний, бо таксон — це континуум, згущення точок, хмара у багатовимірному просторі ознак. Хмара, яка може еволюціонувати, тобто змінювати свої розміри і положення в просторі ознак, може зникнути (вимерти) або, навпаки, розділитися навпіл чи відростити нову хмару тощо. Континуум характеризується як цілісністю, виявленою за наявністю гіатуса (порушення неперервності) між ним і іншими хмарами‑континуумами, так і ознаками його діагнозу. Однак обидва ці критерії не виявляють жорсткості і легко змінюються як у ході еволюції, так і з накопиченням наших знань про таксон. У будь‑який момент може виявитися їхня нездатність чітко ідентифікувати і розмежувати таксони.
Можливість ідентифікації континууму, на відміну від індивіда, визначається схожістю, але на відміну від класу мова йде про відношення (загальне схожість), а не про схожість за конкретними ознаками. Тому вказання пальцем не вдається адресувати ні хмару в цілому, ні її конкретні ознаки. Воно має бути адресовано певній точці цієї хмари — її номенклатурному типу. Що б не сталося з таксоном‑хмарою — у ході її еволюції чи при зміні наших уявлень про неї, ім’я, закріплене за номенклатурним типом, як етикетка за музейним екземпляром, завжди покаже нам, про що йдеться. Якщо, звичайно, тип у збереженні — або хоча б описаний з прийнятною повнотою.
Сказане робить зрозумілим, що номенклатурний тип є природним атрибутом традиційної, тобто філетичної, а також фенетичної системи, і, відповідно, випливає з епігенетичної теорії еволюції. Що стосується кладизму, то кладистичний таксон визначається, як ми бачили, посиланням на подію дивергенції або на синапоморфію, набуту цим таксоном (його першим членом) у момент виникнення. Саме дивергенція або/і набуття синапоморфії розглядаються як подія, що створює таксон. Ця подія є ознакою, єдиною і однозначною, що визначає кладистичний таксон. Таким чином, кладистичний таксон — це клас за визначенням, і для його ідентифікації не потрібен жоден номенклатурний тип: все, що необхідно і достатньо, це, як вже зазначалося, акт дивергенції і/або синапоморфії.
К тому ж результату можна прийти, використовуючи інший і коротший шлях міркувань.Мета будь‑якої класифікації полягає в тому, щоб охопити таксонами весь об’єкт класифікації різноманіття без залишку і, за можливості, без перекриття меж таксонів. Якщо ми приймаємо, що і самі таксони як природні групування, і утворена ними система існують у природі, а не вигадані систематиками (є й така точка зору, див., наприклад, С. Расницин, 2000), то головна мета класифікації двоєдина. По‑перше, простежування природних перерв неперервності в класифікованому просторі (користуючись платонівською метафорою, цитованою Hull, 1983: 186, ми розрізняємо природу за її суглобами). По‑друге, ми фіксуємо і ранжуємо виявлені гіатуси як таксономічні межі (при штучній класифікації ми накладаємо ці межі замість того, щоб простежувати їх).
Класи не піддаються описаній процедурі, оскільки вони визначаються ознаками, для яких природно перекриватися з іншими ознаками (якщо лише не визначати клас так витончено, як я зробив вище, аналізуючи природу кладистичного таксона). Що стосується індивідів, то нам невідомі природні сили і закономірності, які могли б упакувати індивіди щільно, не залишаючи вільних проміжків (знову ж, якщо не розглядати в кладистичному таксоні індивіда на підставі того, що він народжується з дивергенцією свого предка, щоб потім зникнути внаслідок вимирання або дивергенції). Таксон‑континуум, навпаки, ідеально відповідає зазначеній процедурі класифікації, оскільки єдиний законний спосіб визначення континууму — простежування і ранжування його меж.
Не треба думати, що це унікальна властивість загальної системи організмів. З роздумів С. В. Мейєна (1989) очевидно випливає, що такі ж, наприклад, мерон, тобто таксон мерономії (морфологічної системи частин організму), і стратон — таксон стратиграфічної класифікації.
Окрім принципу типу, існують ще два таксономічно залежних принципи номенклатури, які, як і принцип типу, випливають з континуальної природи таксона.
8. Принцип ієрархії. Будучи континуумами, таксони не можуть бути об’єднані іншим чином, ніж у наступні ширші континууми (старші таксони), які повинні охоплювати включені континууми цілком. Так само таксон‑континуум не може одночасно входити в два і більше старших таксона ні цілком, ні частинами, бо інакше ці старші таксони будуть перекриватися, порушуючи тим самим визначення континууму. Все це означає, що система має суворо ієрархічну форму.
9. Принцип синонімії. Якщо таксон може бути членом єдиного старшого таксона, він має єдине місце в системі і, відповідно, йому достатньо одного імені. Усі інші назви повинні бути визнані недійсними, тобто оголошені синонімами.
1.2.3.3. Таксони, що порушують принципи номенклатури
Життя складне, і наука теж. Іноді нам доводиться порушувати принципи номенклатури — не через невігластво чи недбалість, а через необхідність, коли виникають серйозні причини. Розглянемо ці можливі причини.
Таксономічно незалежні принципи являють собою угоди між систематиками і в цьому сенсі суб’єктивні. Причини відмови слідувати їм також завжди суб’єктивні в тому сенсі, що вони не обґрунтовані якимись таксономічними міркуваннями. Тому вони тут не розглядаються. Проблема таксономічно залежних принципів важливіша і заслуговує аналізу. Детальніший такий аналіз був проведений раніше (Расницин, 1986а) і тут подається в скороченій формі.
Порушувати таксономічно залежні принципи номенклатури нам доводиться, коли нестача інформації, необхідної для класифікації певного таксономічного матеріалу, вступає в конфлікт з необхідністю його класифікувати — через наукову чи прикладну його значимість. Неповнота інформації в цих випадках зазвичай пов’язана зі специфічною дефектністю матеріалу, як це добре відомо в палеонтології та в систематиці груп зі складним онтогенезом (наприклад, у паразитичних черв’ях і грибах).
Міжнародні кодекси номенклатури визнають деякі форми таксонів особливого статусу, але не вирішують проблему в цілому, а регламентації в кодексах часто суперечливі. Незадовільність ситуації з таксонами особливого статусу важко назвати несподіваною, принаймні в зоології, де такі таксони традиційно вважаються рідкісними і вкрай небажаними. В цілому (як у зоології, так і в ботаніці) вони дійсно не надто численні, проте в деяких специфічних галузях (палеонтологія, паразитологія, мікологія) відмова від їх використання загрожує великою втратою інформації про об’єкти, що мають важливе стратиграфічне, медичне, агро‑ або лісогосподарське значення. Використання «неповноцінних» таксонів неминуче, але при їх неповній регламентації кодексами це веде до номенклатурного розбрату і самодіяльності. У палеонтології, наприклад, принципово однорідні групування в окремих випадках позначають як спеціальні таксономічні категорії (іхнотактсони в їхнології, зокрема «індузироди» і «індузиподроди» в систематиці викопних будинків, побудованих личинками комах‑ручейників (В’ялов, Сукачева, 1976), турми, антитурми тощо в палеопалінології, формальні роди і нещодавно скасовані орган‑роди в систематиці макроструктур рослин тощо). В інших випадках беруть у лапки назву таксона (Белокрис, 1984) або таксономічної категорії (Пономаренко, 1985), у третіх — просто уточнюють статус таксона в описі чи преамбулі. Іноді справа доходить до розробки спеціальних кодексів номенклатури (Sarjeant, Kennedy, 1973).
Цей розбрат незручний, хоча й не небезпечний. Більше того, він навіть корисний як спосіб пошуку оптимальних форм роботи з «неповноцінними» таксонами. Небезпечно породжене негативним ставленням до таких таксонів бажання представити їх нормальними, повноцінними. Дивно, наприклад, прагнення багатьох фахівців з конодонтів (ізольованих мікроскопічних елементів скелета древніх тварин) показати їх таксони повноцінними (Melville, 1981a,b). Ще більш демонстративна ситуація з сучасним родом Laberius Kieffer (перепончатокрилі комахи родини Dryinidae). Цей род був описаний (під назвою Labeo Hal., nom. praeocc.) як звичайний таксон, але потім використаний як збірна група для самців триб Dryinini і Gonatopodini (Kiefer, 1914). Ці триби характеризуються сильним статевим диморфізмом, причому їх система побудована за ознаками самок, а самців, якщо вони не асоційовані з самками експериментально, до недавнього часу не вдавалось визначити навіть до триби. Однак у ревізії родини (Olmi, 1984) її автор відмовився від такої інтерпретації роду Laberius і зводив його в синоніми роду Dicondylus Haliday (не обговорюючи при цьому ознаки типового виду Laberius, тобто не обґрунтовуючи свого рішення). При цьому вимерлий вид, описаний у складі роду Laberius, що розуміється як збірна група (Н. Пономаренко, 1981), був переведений у род Dryinus Latreille з вказанням, що єдиний екземпляр цього виду збережений «настільки погано, що це не дозволяє віднести його до якогось роду родини Dryinidae» (Olmi, 1984). Таким чином Dryinus (типовий род родини) був фактично перетворений у збірну групу, тобто найнеповноціннішу форму таксона (див. нижче), хоча мета автора була явно протилежною — позбутись неповноцінного таксона.
Отже, проблема таксонів, що не задовольняють тим чи іншим правилам номенклатури, залишається актуальною. Проаналізуємо детальніше природу і форми таких таксонів (табл. 5).
Таблиця 5. Форми таксонів
| Таксон | Належить системі | Тип і діагноз | При синонімії з ортотаксоном |
|--------|------------------|---------------|------------------------------|
| ортотаксон | загальній | власний | підлягає синонімізації |
| таксон неясного положення | загальній | власний | підлягає синонімізації |
| морфотаксон (паратаксон) | спеціальній | власний | не підлягає синонімізації |
| збірний таксон (паратаксон) | спеціальній | старшого таксона | не підлягає синонімізації |Найлегший випадок — це таксон невизначеного положення (taxon incertae sedis). Він відрізняється від звичайного таксона (optotaxon) тим, що неповнота його характеристики не дозволяє уточнити його розташування лише на певному рівні ієрархії. Наприклад, рід невизначеного положення (genus incertae sedis) наразі може бути з достатньою впевненістю віднесений до певного відряду, але не до сімейства. Вимерлий рід Baissobius Rasn., описаний серед перетинчастокрилих комах підроду Apocrita, «володіє своєрідним типом редукованого жилкування, якого, наскільки відомо, не спостерігається у інших Hymenoptera» (Расніцин, 1975, с. 128). При цьому його жилкування було досить своєрідним, щоб ускладнити визначення систематичного положення комахи (інші важливі ознаки на викопному не видно), але недостатньо своєрідним, щоб виправдати виділення окремої родини. І справді, нові знахідки дозволили розпрощатися з цим таксоном невизначеного положення, перетворивши його на звичайний рід, що належить родині Embolemidae (Rasnitsyn, 1996b).
Використання таксонів невизначеного положення можна зрозуміти як локальне відхилення від принципу синонімії. Рід, підпорядкований відряду, минаючи сімейство, фактично виступає як самостійне, але неназване сімейство, ймовірно синонімне якомусь із існуючих сімей, лише поки невідомо, якому саме. Як видно на прикладі Baissobius, з часом ця синонімія дійсно може стати відомою.
Більш складні випадки, об’єднані поняттям паратаксонії. Визначення, що трактує паратаксон як таксон у часткових системах для ізольованих органів і частин тіла тварин (Melville, 1979), вже практичного застосування цього поняття. Дійсно, специфіка і проблеми, пов’язані з введенням і використанням таксономічної назви, мало відрізняються у часткових системах, створених для ізольованих органів і частин тіла організмів, для певної статі або стадії життєвого циклу, або для слідів життєдіяльності. Тому тут прийнято інше визначення: паратаксон, на відміну від ортотаксона і таксона невизначеного положення, належить самостійній системі, пов’язаній з основною системою принципом гомонімії, а не принципом синонімії, і тому не підлягає синонімізації навіть у тих випадках, коли вказана його синонімія з якимось ортотаксоном. Таким чином, паратаксон — це таксон, на який не поширюється вимога унікальності таксономічного положення, тобто вимога входження в єдину для всіх організмів ієрархію. Саме до цього, зокрема, зводяться всі п’ять ознак своєрідності викопного матеріалу, сформульовані С.В. Мейеном (Meyen, Traverse, 1979; Мейен, 1983). Важливі міркування про природу і функції паратаксона наведені в іншій роботі (Мейен, 1990).
Розрізняють кілька видів паратаксонів. Термін формальний таксон був запропонований (Расніцин, 1986а) як ширша інтерпретація ботанічного поняття форм‑рід. Нещодавно (МКБН, 2000) ботаніки замінили цей термін більш вдалим поняттям морфотаксон, яким ми і скористаємося. Морфотаксон — це еквівалент звичайного таксона (ортотаксона), але використовується лише в рамках спеціальної системи. Ця система паралельна як основній системі, так і іншим подібним частковим системам, але незалежна від них щодо принципу синонімії. Такі системи існують для ізольованих викопних листків, насіння або відбитків крила жука, викопних слідів переміщення організмів і інших форм їхньої життєдіяльності (наприклад, будиночків личинок струмків), системи личинкових стадій сучасних паразитичних черв’їв або не асоційованих із самицями самців у деяких групах комах, де система орієнтована на ознаки самок.
Морфотаксон відрізняється від ортотаксона лише тим, що може бути, а іноді й точно є синонімом якогось ортотаксона, і все ж не підлягає синонімізації. Добре відомо, наприклад, що у ряді випадків для розрізнених частин стародавніх рослин (листя, стовбури, корені, чоловічі та жіночі фруктіфікації, насіння і пилок), описаних кожна у своїй спеціальній системі, відомі також знахідки у живій зв’язці, що дозволяють «зібрати» ціле рослина. Проте кожна з таких частин зберігає свою назву, отриману як морфотаксон. Так, добре збережені вуглецеві стовбури Lepidodendron, стовбури Aspidiaria, Bergeria, Aspidiopsis, Knorria з корою, що опала на різну глибину, корені Stigmaria, жіночі стробіли Lepidocarpon, чоловічі стробіли Lepidostrobus і мікроспори Lycospora (рис. 7) могли належати одній і тій самій рослині, оскільки живе поєднання різних пар із цього списку більш‑менш надійно встановлено (Мейен, 1987б). І ніхто при цьому не намагається спростити валідність усіх цих родових назв!
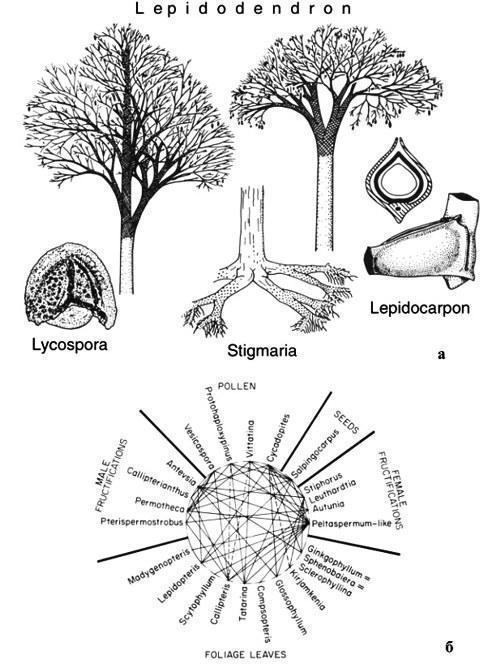
Рис. 7. Формальні таксони, запропоновані для класифікації ізольованих частин лепідофітів (за Мейеном, 1987б, 1988): а — Lepidodendron для стовбурів із збереженою корою (Aspidiaria, Bergeria, Aspidiopsis, Krorria для стовбурів, у різній мірі втратили кору), Stigmaria для коренів, Lepidocarpon для жіночих стробілів (показано загальний вигляд і розріз мегаспорангія), і мікроспора Lycospora, виділена з мікроспорангія Lepidostrobus; б — усі ці назви в деяких випадках означають одне й те саме рослина, частини якої в різних поєднаннях неодноразово знаходилися у живій зв’язці, як це показано лініями (пунктир — менш надійні свідчення живої зв’язки).
Щодо деяких випадків цього роду стверджується, що вони торкаються і принципу типу. Іноді резонність таких тверджень важко зрозуміти (як у випадку відмови типізувати їхнотаксони, тобто таксони системи викопних слідів, у версії МКЗН, 1988, ст. 66), в інших випадках причини докладно обговорюються. А.Г. Пономаренко (1985) встановив нетипізовані формальні таксони, визначені лише ознаками їх діагнозу («Рід [Flichea Handlirsch.—А.Р.] розуміється як формальне об’єднання ізольованих надкрилий жукiв без виразних борозен на верхній стороні і з помітною вирізкою посередині зовнішнього краю», с. 76]). Як пояснив мені автор, причиною стало вперте прагнення деяких систематиків синонімізувати і таким чином ліквідувати паратаксони, якою б довільною не була така синонімізація, і зокрема згаданий вище приклад з родом Laberius Kieffer. Нетипізовані таксони Пономаренка — це, по суті, ті ж морфотаксоні, але штучно детипізовані і тим самим виведені з‑під юрисдикції кодексу номенклатури (оскільки кодекс регламентує застосування назви посиланням на тип, див.‑ вище). Детипізація дійсно запобігає необґрунтованій синонімізації, але проблема в тому, що вона виключає будь‑яку синонімізацію і тому виявляється ліками гіршими, ніж сама хвороба. Мені здається, що розуміння цілей і функцій паратаксонії буде кращим захистом від довільної синонімізації.
Останній і найбідніший за змістом вид паратаксона — це збірний таксон (collective taxon). Це паратаксон, зазвичай у ранзі роду, який віднесений до певного старшого таксона, але на відміну від морфотаксона не може бути виділений у спеціальну систему, паралельну основній. По суті, види у збірних групах — це просто види невизначеного положення (species incertae sedis). Наприклад, назва «Cercaria O.F. Muller, 1773 встановлена для роду черв’їв, і багато авторів у XIX ст. зверталися з нею як з назвою номінального роду, ніби його типовим видом був C. lemna O.F. Muller, 1773. Наразі Cercaria використовується як назва збірної групи для личинок триматод, які не можуть бути з упевненістю віднесені до звичайних родів» (МКЗН, 2000, ст. 67.14). Різниця між таксонами невизначеного положення і видами збірного роду полягає лише в тому, що видовий епітет не має права з’являтися самостійно, без назви роду. Тому для таких видів доводиться вводити назву збірної групи, яка має вигляд родової назви, але виконує роль назви того старшого таксона, до якого автор відносить види, включені до даного збірного роду. Так, Cercaria у сучасному розумінні — це еквівалент відряду триматод, Laberius у інтерпретації Й. Kieffer (Kieffer, 1914) — еквівалент Dryinini + Gonatopodini, а Dryinus у інтерпретації М. Olmi (Olmi, 1984) — еквівалент Dryinidae.
Збірні таксони вважаються детипізованими, але це твердження помилкове. Наведене вище визначення означає, що збірний таксон за характеристикою і обсягом збігається з відповідним старшим таксоном, і його члени, подібно до ортотаксонів, вводяться в систему в кінцевому підсумку посиланням на номенклатурний тип останнього. Види Cercaria визначаються як такі просто тому, що достатньо схожі з личинками типового виду триматод. А те, що типовий вид триматод поки формально не зафіксований, не суттєво: я не сумніваюся, що старші таксони будуть типізовані і в зоології. Інший приклад — збірний рід Carabilarva Ponomarenko, запропонований для мезозойських личинок жукiв надродини Caraboidea, не визначуваних до сімейства (А.Г. Пономаренко, 1985). Тому типовим видом роду є тип надродини Carabus granulatus Linnaeus, 1758. Отже, Carabilarva є молодшим об’єктивним синонімом роду Carabus, але не підлягає синонімізації з ним, доки залишається паратаксоном.
Останній приклад означає існування ще однієї форми множинного підпорядкування в номенклатурі, окрім вже обговорюваного випадку, коли різні форми збереженості одного природного таксона описуються як різні морфотаксоні. Тут вже один і той самий вид виявляється типовим для безлічі збірних груп. Дійсно, рід Carabus одночасно є типовим для родини Carabidae (жуки‑жужеліці), підродини Сагаbинае, триби Сагаbіпі тощо, а для далі невизначених членів кожного з цих таксонів теоретично можуть знадобитися свої збірні групи, ще різні для личинок і різних ізольованих частин тіла дорослого жука. Легко уявити, скільки збірних родів у підсумку можуть виявитися заснованими на одній і тій самій номенклатурній типі. При всій незвичності ситуації, вона навряд чи може викликати серйозні номенклатурні чи таксономічні проблеми, якщо ми будемо розуміти, навіщо потрібні збірні таксони. Адже нікому не заважають роди і старші таксони невизначеного положення, чому ж ставлення до видів невизначеного положення має бути іншим?
Існує ще один важливий аспект проблеми неповноцінного таксона (таксона особливого статусу). Таксон належить цій категорії просто тому, що він виділений за «поганими» ознаками — поганими у порівнянні з іншими, звичайними таксонами. Якщо ж інших таксонів немає, той самий неповноцінний таксон виявляється цілком повноцінним. Якби сучасних струмків не існувало, викопні будиночки, ймовірно, вивчалися б поза будь‑яким зв’язком з викопними струмками, і їхня система, заснована на тих самих «поганих» ознаках, вважалася б звичайною. У багатьох групах комах сучасні види нині розрізняються практично лише за ознаками чоловічих геніталій, а їхні вимерлі представники — головно за жилкуванням крил. Поки викопні відносять до особливих, вимерлих родів, номенклатурних проблем не виникає, і з викопними можна працювати як з ортотаксонами. Там, де вимерлий вид потрапляє в один рід (підрід) із сучасними, він часто опиняється в категорії неповноцінного. Слабким розраюванням тут служить те обставина, що те саме заслуговують і давно описані сучасні види, для яких ще не вдалося описати будову геніталій.
Окрім умовності кваліфікації таксона як орто‑ або паратаксона, існують складнощі іншого роду. Хороший ознака чи погана — це, по‑перше, міра (наскільки велика різниця), по‑друге, смак. Звична справа, коли суперечка між систематиками зводиться, в кінцевому підсумку, до того, на яку систему ознак слід спиратися, розробляючи систему конкретної групи. Можна лише здивуватись, що у суперечці ще не виникають взаємні звинувачення, що, так би мовити, система опонента, заснована на таких‑то ознаках, корисна лише як часткова система паратаксонів, а не загальна (основна) система групи.
Усі ці міркування дозволяють зробити висновок, що ортотаксон і паратаксон формально чітко різні, але змістової обособленості тут немає. Залежно від точки зору, а тим більше, як ми бачили, від рівня наших знань (приклад Baissobius), таксон може переходити з однієї категорії в іншу. Схоже, до паратаксонії треба ставитися прагматично: якщо використання паратаксонів реально полегшує пізнання і інвентаризацію біорізноманіття, їх треба вводити і застосовувати. Якщо ж вигода невелика, не треба ускладнювати ситуацію введенням паралельних, перекриваючих систем організмів.
1.3. ВИСНОВОК{
"translated_text": "Проблеми еволюційного крила сучасної біології, куди можна віднести ті її розділи, що досліджують історію та сучасний стан біологічного різноманіття, включаючи їх теорію і методологію, пов’язані передусім з пошуками парадигми. Уже більше століття триває суперечка двох підходів — «елементаристської» («редукціоністської») і «голістичної» парадигм. Дивовижно, але «розмежування» цих так різних підходів не відбувається, а продукти кожного з них співіснують у головах і друкованій продукції майже у більшості вчених, про що вони зазвичай і не підозрюють.\nЕлементаристській парадигмі, що редукує вивчувану систему до її елементів, відповідає популяційно‑генетична (синтетична) теорія еволюції, що представляє еволюційний процес як динаміку частот дискретних спадкових детермінант (генів) у популяціях за умов відбору. Відображенням цієї динаміки є спостережуваний процес еволюції, і її специфіка визначається, по‑перше, специфікою генетичного рівня організації, по‑друге, специфікою відбору. Відповідно, еволюційний процес виступає як суттєво рівномірний, а єдиним регулярним джерелом дискретності, що робить можливим розбиття продукту еволюції на таксони, виявляються дивергенція і вимирання. Цій парадигмі адекватна таксономічна концепція кладизму, де таксон виникає в ході еволюції як продукт дивергенції і вводиться в систему саме на цьому підґрунті, а послідовність дивергенцій визначається за допомогою синапоморфій.\nАльтернативна голістична парадигма редукує еволюційну систему до цілого і, зокрема, до епігенотипу, тобто до системи побудови живого організму в онтогенезі. У дзеркалі епігенетичної теорії еволюції складна, пронизана переплетеними взаємозв’язками і взаємодіями жива система являє собою напружений компроміс протилежних вимог оптимізації різних адаптивних функцій. Зміна такої системи виявляється суттєво ускладненою, малопередбачуваною і більш‑менш стрибкоподібною, а специфіка еволюційного процесу визначається передусім специфікою епігенотипу і лише в суттєво меншій мірі — специфікою відбору. Через стрибкоподібність еволюційних змін біологічне різноманіття, що виникає в ході такої еволюції, виявляється дискретним, розбитим гіатусами на природні окремості, які і заслуговують використання в якості таксонів загальної системи. Відповідно, сенс таксономії, адекватної епігенетичній концепції і голістичній парадигмі, полягає у виявленні об’єктивно існуючої дискретності і простежуванні гіатусів.\nАналіз доступного матеріалу свідчить на користь його відповідності радше епігенетичній, ніж синтетичній концепції. Більш важливою, однак, здається сама спроба розрізнення концепцій і методологій, що використовуються в аналізі еволюційного процесу і його результатів, як походять від тієї чи іншої з двох конкуруючих парадигм. Схоже, що суворе розрізнення двох парадигм і їх наслідків дозволить не лише і навіть не стільки зробити осмислений вибір між ними, скільки позбутися одночасного використання взаємно несумісних принципів і підходів, тобто від свого роду плюралізму в одній голові.\nЗроблений висновок дозволяє дещо просунутися у виявленні коренів і у більш чіткому формулюванні методологічних концепцій, що використовуються на полі еволюційного крила біології, зокрема у філогенетиці, таксономії і таксономічній номенклатурі. Однак спочатку нам довелося розглянути деякі найзагальніші проблеми наукового (і не лише) аналізу.\nФілогенетичні висновки, як будь‑яке наукове дослідження, спираються на (1) планування майбутнього дослідження, (2) спостереження (включаючи експеримент), (3) пошук аналогій, (4) створення гіпотез про лежачі в основі видимої картини закономірності і механізми, (5) спроби фальсифікації цих гіпотез (переважно шляхом аналізу їх наслідків), і (6) оцінки результатів цих спроб за допомогою презумпцій, з метою вибору найбільш перспективної гіпотези для подальшої роботи.\nНабір філогенетичних презумпцій включає презумпцію пізнаваності філогенезу і ряд більш конкретних презумпцій, які можна розділити на використані в аналізі груп і в аналізі ознак. Аналіз груп досліджує відносини предків і нащадків і спирається передусім на палеонтологічну презумпцію для груп. Презумпції, використані в аналізі ознак, далі діляться на дві групи відповідно до того, чи включені вони в аналіз різниць або схожостей. Аналіз різниць — це поляризація трансформаційних серій, тобто визначення плезіоморфних (вихідних) і апоморфних (змінених) станів ознаки. Тут працюють, з більш важливих, палеонтологічна презумпція для ознак, біогенетична презумпція, презумпції аналогії, необоротності еволюції, функціональної досконалості, складності, рудиментів і збереженого розповсюдження ознак. Презумпції аналізу схожостей використовуються при вирішенні питання, чи успадковано дану схожість від спільного предка чи вона виникла незалежно. Сюди належать передусім презумпції парсимонії і зваженої схожості.\nМета таксономії полягає у створенні системи, чиї таксони осмислені для максимально широкого кола користувачів і, відповідно, найгомогенніші всередині себе і різні між собою. З трьох головних конкуруючих підходів кладизм спирається на синтетичну теорію еволюції, що ігнорує дискретність біорізноманіття. Тому кладистична система враховує лише дивергенції і визнає лише таксони, охарактеризовані синапоморфіями. Синапоморфія — не ознака, а її інтерпретація, тому кладистична система не лише ігнорує дискретність біорізноманіття, а й надмірно гіпотетична і привередлива у відборі ознак.\nФілетика і фенетика спираються на епігенетичну теорію еволюції і зайняті простежуванням гіатусів між таксонами. Фенетика використовує доступні ознаки як такі і тому надто близькозорка, це суттєво емпіричний підхід. Філетика займає проміжне положення між фенетикою і кладизмом. Вона намагається відобразити повний баланс схожостей і різниць, включаючи і ті, що ще не вивчені. Для цього філетика використовує прогностичні можливості філогенезу, який виступає в ролі евристичного методу контролю системи, побудованої фенетичними методами. Філетика визначає таксон як монофілетичний континуум і використовує гіатуси для розмежування таксонів, а монофілію — як спосіб оцінити, чи успішне було розмежування, чи система потребує доопрацювання.\nУ основі таксономічної номенклатури лежать 9 принципів, з яких шість представляють чисті домовленості для зручності і уніфікації у використанні назв таксонів і незалежні від використаного таксономічного підходу. Інші три принципи залежать від таксономії. З них найважливіший принцип типу, що дозволяє стабілізувати назви таксонів, що відповідають поняттю монофілетичного континууму. Два інші таксономічно залежних принципи — принципи ієрархії і синонімії, з яких другий може бути порушений при роботі з дефектним таксономічним матеріалом, недостатньо охарактеризованим для його повноцінної систематизації. У цьому випадку можуть застосовуватись таксони неясного положення і паратаксон різного роду, що використовуються в рамках спеціальних систем, паралельних основній системі організмів.\nНа відміну від принципів синонімії, очевидно, не існує об’єктивних причин відмовлятись від дотримання принципів типу і ієрархії, а тим більше від таксономічно незалежних принципів номенклатури.\nЛІТЕРАТУРА\nАлексеев А.С., Дмитриев В.Ю., Пономаренко А.Г. 2001. Еволюція таксономічного різноманіття. Екосистемні перестройки і еволюція біосфери. Вип. 5. М.: Геос. 126 с.\nАристотель. Біомедґіз. 1937. 220 с.\nБекетов А.Н. 1860. Гармонія в природі // Руский вестник. № 30. С. 197‑240, 534‑558.\nБелокрис Л.С. 1984. Спірорбіни причерноморського міоцену // Палеонтол. журн. № 2. С. 26‑39.\nБеляев Д.К. 1974. Про деякі питання стабілізуючого і дестабілізуючого відбору. Історія і теорія еволюційного вчення, Вип. 2. Л.: Наука: 76‑84.\nБерман Д.І. 2001. Безхребетні тварини. Заключення // Холодні степи Північно‑Східної Азії / Берман Д.І. (ред.). Магадан: Ін‑т біол. Проблеми Півночі СВНЦ ДВО РАН. С. 96‑167.\nВахрамеев В.А., Котова І.З. 1977. Найдавніші покритосемені і супутні їм рослини з нижньомелових відкладень Забайкалья // Палеонтол. журн. № 4. С. 101‑109.\nВейсман А. 1905. Лекції з еволюційної теорії. М.: Видав‑во М. і С. Сабашникових. Т. 1. 505 с.\nВороб’єва Е.І. 1992. Проблема походження наземних хребетних. М.: Наука. 344 с.\nВялов О.С., Сукачева І.Д. 1976. Викопні будинки личинок ручейників (Insecta, Trichoptera) і їхнє значення для стратиграфії // Палеонтологія і біостратиграфія Монголії. Тр. Совм. Сов‑Монг. палеонтол. експед. Вип. 3. С. 169‑230.\nГрант В. 1980. Еволюція організмів. М.: Мир. 480 с.\nГриценко В.В., Креславський А.Г., Міхеєв А.В., Північне А.С., Соломатин В.М. 1983. Концепції виду і симпатричне видоутворення. М.: Видав‑во МГУ 194 с.\nГродницький Д.Л. 2000. Дві теорії біологічної еволюції. Красноярськ: Ін‑т лісу ім. В.Н. Сукачева СО РАН. 180 с.\nГродницький Д.Л. 2002. Дві теорії біологічної еволюції. 2‑й вид. Саратов: Наукова книга. 160 с.\nДарвін Ч. 1991. Походження видів шляхом природного відбору. СПб.: Наука. 540 с.\nДіяльність міжсекційного семінару з проблем еволюції з жовтня 1965 р. по квітень 1966 р. 1967 // Бюл. Моск. о‑вип. вип. природи. Відд. біол. Т. 72. Вип. 4. С. 136‑138.\nЖегалло В.І., Каландадзе Н.Н., Кузнецова Т.В., Раутиан А.С. 2001. Доля мегафауни Голарктики в пізньому антропогенезі / Мамонт і його оточення: 200 років вивчення. М.: ГЕОС. С. 287‑306.\nЖерихін В.В. 1978. Розвиток і зміна мелових і кайнозойських фауністичних комплексів (трахейні і хеліцерові) // Тр. Палеонтол. ін‑т АН СССР. Т. 165. М.: Наука. 200 с.\nЖерихін В.В. 1987. Біоценотична регуляція еволюції // Палеонтол. журн. № 1. С. 312.\nЖерихін В.В. 1993. Історія біома тропічних дощових лісів // Журн. заг. біол. Т. 54. С. 659‑666.\nЖерихін В.В., Расницин А.П. 1980. Біоценотична регуляція еволюції. Мікро‑ і макроеволюція. Матеріали симпозіуму. Кяеріку. 2‑5 вер. 1980. Тарту. С. 77‑81.\nЗаварзін Г.А. 1987. Особливості еволюції прокаріот // Еволюція і біоценотичні кризи / Л.П. Татаринов, А.П. Расницин (ред.). М.: Наука. С. 144‑158.\nІванова Л.С. 1978. Порівняльне вивчення партеногенетичних довгоносиць (Coleoptera, СитшІо^ае) Сибіру: Автореф. дис ... канд. біол. наук. Новосибірськ: Біол. ін‑т СО АН СССР. 18 с.\nІорданський Н.Н. 1977. Нерівномірність темпів макроеволюції і ключові ароморфози // Природа. № 6. С. 36‑46.\nІорданський Н.Н. 1979. Основи теорії еволюції. М.: Просвітлення. 190 с.\nКлюге Н.Ю. 2000. Сучасна систематика комах. Частина I. Принципи систематики живих організмів і загальна система комах з класифікацією первиннобескрилих і древнекрилих. СПб.: Лань. 336 с.\nКрасилов В.А. 1969. Філогенія і систематика // Проблеми філогенії і систематики: Матеріали симпоз. Владивосток. С. 12‑30.\nКурочкін Є.Н. 2006. Паралельна еволюція тероподних динозаврів і птахів // Зоол. журн. Т. 85. С. 283‑297\nЛюбищев А.А. 1923. Про форму природної системи організмів // Ізв. Біол. н‑і. ін‑т при Пермськ. ун‑те. Т. 2. Вип. 3. С. 99‑110. (см. також у кн.: Любищев А.А. Проблеми форми, систематики і еволюції організмів. М.: Наука. 1982. С. 24‑36).\nЛюбищев А.А. 1982. Проблеми форми, систематики і еволюції організмів. М.: Наука. 280 с.\nМайр Е. 1974. Популяції, види і еволюція. М.: Мир. 460 с.\nМамкаєв Ю.В. 1968. Порівняння морфологічних різниць у нижчих і вищих групах одного філогенетичного стовпа // Журн. заг. біол. Т. 29. С. 48‑55.\nМейен С.В. 1983. Порівняння принципів систематики сучасних і викопних організмів // Математика і ЕВМ у палеонтології. Кишинів: Штиинца. С. 10‑25.\nМейен С.В. 1987. Основи палеоботаніки. М.: Недра. 402 с.\nМейен С.В. 1989. Вступ до теорії стратиграфії. М.: Наука. 215 с.\nМейен С.В. 1990. Палеоботанічна таксономія і номенклатура: необхідність нового підходу // Теоретичні проблеми палеоботаніки. М.: Наука. С. 49‑76.\nМКЗН 1988: Міжнародний кодекс зоологічної номенклатури. Вид. 3. Л.: Наука. 1988. 205 с.\nМКЗН 2000: Міжнародний кодекс зоологічної номенклатури. Вид. 4. СПб.: Наука. 2000. 221 с.\nНазаров В.І. 1984. Реконструкція ландшафтів північно‑східної Білорусі в антропогенезі за палеоентомологічними даними // Тр. Палеонтол. ін‑т АН СССР. Т. 205. М.: Наука. 96 с.\nНовокшонов В.Г. 1997. Рання еволюція скорпіонниць. М.: Наука: 140 с.\nПономаренко А.Г., Пацницин А.П. 1971. O фенетичній і філогенетичній системах // Zooл. журн. Т. 50. № 1. С. 5‑14.\nПолянський В.І. 1956. Про вид нижчих водоростей // Комаровські читання. М‑Л.: Видав‑во АН СССР. 73 с.\nПолянський Ю.І. 1957. Про внутрішньовидову диференціацію і структуру виду у простих // Вестн. ЛГУ № 21. С. 45‑64.\nПономаренко А.Г. 1985. Жорсткокрилі з юри Сибіру і західної Монголії // Юрські комахи Сибіру і Монголії. Тр. Палеонтол. ін‑т АН СССР. Т. 211. М.: Наука. С. 478‑7.\nПономаренко Н.Г. 1981. Нові перепончатокрилі комахи родини Dryinidae з пізнього мела Таймиру і Канади // Палеонтол. журн. № 1. С. 139‑143.\nПоппер К. 1983. Логіка і ріст наукового знання. М.: Прогрес. 606 с.\nРазумовський С.М. 1971. Про походження і вік тропічних і лавролістних флор // Бюлл. ГБС АН СССР. Т. 82. С. 43‑51.\nРасницин А.П. 1969. Походження і еволюція нижчих перепончатокрилих // Тр. Палеонтол. ін‑т АН СССР. Т. 123. М.: Наука. 196 с.\nРасницин А.П. 1971. До питання про причини морфофункціонального прогресу // Журн. заг. біол. Т. 32. № 5. С. 549‑556.\nРасницин А.П. 1975. Вищі перепончатокрилі мезозою // Тр. Палеонтол. ін‑т АН СССР. М.: Наука. Т. 147. 134 с.\nРасницин А.П. 1986а. Інадаптація і евадаптація // Палеонтол. журн. № 1. С. 3‑7.\nРасницин А.П. 1987. Темпи еволюції і еволюційна теорія (гіпотеза адаптивного компромісу) // Еволюція і біоценотичні кризи. М.: Наука. С. 46‑64.\nРасницин А.П. 1988а. Філогенетика / Сучасна палеонтологія. Т. 1. М.: Недра. С. 480‑497.\nРасницин А.П. 2002. Процес еволюції і методологія систематики // Тр. Русск. ентомол. о‑в. Т. 73. СПб. 107 с.\nРасницин А.П. 2005. Вибрані праці з еволюційної біології. М.: КМК. 347 с.\nРасницин А.П., Длусський Г.М. 1988. Принципи і методи реконструкції філогенезів // Меловий біоценотичний кризис і еволюція комах. М.: Наука. С. 5‑15.\nРасницин С.П. 2000. Майбутнє ентомологічної систематики // Захоплюючі есе про діяльність і діячів протипухової системи Росії і СРСР. Вип. 10. М.: Інформатика. С. 189‑196.\nРаутиан А.С. 1988. Палеонтологія як джерело відомостей про закономірності і фактори еволюції // Сучасна палеонтологія. Т. 2. М.: Недра. С. 76‑118.\nРодендорф Б.Б. 1977. Про раціоналізацію назв таксонів високого рангу в зоології // Палеонтол. журн. № 2. С. 14‑22.\nРодендорф Б.Б., Жерихін В.В. 1974. Палеонтологія і охорона природи // Природа № 2. С. 82‑91.\nСеверцов С.А. 1941. Динаміка населення і пристосувальна еволюція тварин. М‑Л.: Видав‑во АН СССР. 316 с.\nСімпсон Дж. 1948. Темпи і форми еволюції. М.: Видав‑во інстр. літ. 358 с.\nТатаринов Л.П. 1976. Морфологічна еволюція теріодонтів і загальні питання філогенетики. М.: Наука. 258 с.\nЦвєлєв Н.Н. 1993. Гібридизація як один з факторів збільшення біологічного різноманіття і геномний критерій роду у вищих рослин // Біологічне різноманіття: підходи до вивчення і збереження. СПб.: Зоол. ін‑т РАН. С. 193‑201.\nЧайковський Ю.В. 2003. Еволюція // Ценологічні дослідження. Вип. 22. М.: Центр системних досліджень - ІІЕТ РАН. 472 с.\nШапошников Г.Х. 1961. Специфічність і виникнення адаптації до нових господарів у тлі (Homoptera, Aphidodea) у процесі природного відбору (експериментальне дослідження) // Ентомол. обозр. Т. 40. № 4. С. 739‑762.\nШапошников Г.Х. 1965. Морфологічна дивергенція і конвергенція в експерименті з тлями (Homoptera, Aphidinea) // Ентомол. обозр. Т. 44. № 1. С. 3‑25.\nШапошников Г.Х. 1966. Виникнення і втрата репродуктивної ізоляції і критерій виду // Ентомол. обозр. Т. 45. № 1. С. 3‑35.\nШапошников Г.Х. 1978. Динаміка клонів, популяцій і видів і еволюція // Журн. заг. біол. Т. 39. № 1. С. 15‑33.\nШарп Д. 1910. Комахи. СПб.: Брокгауз‑Ефрон. 1054 с.\nШварц Є.А. 2004. Збереження біорізноманіття: спільноти і екосистеми. М.: КМК. 112 с.\nШварц С.С. 1980. Екологічні закономірності еволюції. М.: Наука. 278 с.\nШер А.В. 1997а. Природна перестройка в Східно‑Сибірській Арктиці на межі плейстоцену і голоцену і її роль у вимиранні ссавців і становленні сучасних екосистем (Повідомлення 1) / Кріосфера Землі. Т. 1. № 1. С. 21‑29.\nШер А.В. 1997б. Природна перестройка в Східно‑Сибірській Арктиці на межі плейстоцену і голоцену і її роль у вимиранні ссавців і становленні сучасних екосистем (Повідомлення 2) // Кріосфера Землі. Т. 1. № 2. С. 3‑11.\nШишкін М.А. 1984. Індивідуальний розвиток і природний відбір // Онтогенез. Т. 15. № 2. С. 115‑136.\nШишкін М.А. 1987. Індивідуальний розвиток і еволюційна теорія // Еволюція і біоценотичні кризи. М.: Hayra. С. 76‑124.\nШишкін М.А. 1988а. Еволюція як епігенетичний процес // Сучасна палеонтологія. Т. 1. М.: Хедра. С. 142‑169.\nШишкін М.А. 1988б. Закономірності еволюції онтогенезу // Сучасна палеонтологія. Т. 1. М.: Недра. С. 169‑209.\nШишкін М.А. 2006. Індивідуальний розвиток і уроки еволюціонізму // Онтогенез. № 37. С. 179‑198.\nЕшбі У.Р. 1964. Конструкція мозку. М.: Мир.\nAlberch P. 1980. Ontogenesis and morphological diversification // Amer. Zool. Vol. 20. P. 653‑667.\nAshlock P.D. 1971. Monophyly and associated terms // Syst. Zool. Vol. 20. P. 63‑69.\nBaer K.E., von. 1828. Uber Entwickelungsgeschichte der Thiere. Beobachtung und Reflexion. Bd. 1. Konigsberg: Gebr. Borntrager. 271 S.\nBonde N. 1984. Primitive features and ontogeny in phylogenetic reconstructions // Vidensk. Meddr. dansk naturh. Foren. Vol. 145. P. 219‑236.\nBrothers D.J. 1975. Phylogeny and classification of the aculeate Hymenoptera, with special reference to Mutillidae // Univ. Kansas Sci. Bull. Vol. 50. P."
}483-648.
Carson H.L.T. 1975. Генетика видоутворення на диплоїдному рівні // Amer. Natur. Vol. 109. P. 8392.
Cope E.D. 1904. Первинні фактори органічної еволюції. Чикаго: Open Court Publ. Co. 547 с.
Cracraft J. 1974. Філогенетичні моделі та класифікація // Syst. Zool. Vol. 23. P. 71-90.
Eldredge N., Gould S. J. 1972. Пунктуальні рівноваги: альтернатива філетичному градіалізму. // Models in Paleobiology / Schopf T.J.M. (ed.) Сан‑Франциско: Freeman. P. 82-115.
Eskov K.Yu. 1987. Новий археїдний павук (Chelicerata: Araneae) з Юрського Казахстану, з нотатками про так звані «Гондванські» ареали сучасних таксонів // Neues Jb. Geol. Palaeont. Abh. 175. P. 81-106.
Eskov K.Yu. 1992. Археїдні павуки з еоценового балтійського янтаря (Chelicerata: Araneida: Archaei‑dae) з зауваженнями щодо так званих «Гондванських» ареалів сучасних таксонів // Neues Jb. Geol. Palaeont. Abh. 185. P. 311-328.
Farrer D.R. 1990. Вид і еволюція у безстатевих самостійних гаметофітів папоротей // Syst. Bot. Vol. 15. P. 98-111.
Farris J.S. 1979. Інформаційний вміст філогенетичної системи // Syst. Zool. Vol. 28. P. 483‑519.
Farris J.S. 1983. Логічна основа філогенетичного аналізу // Advances in cladistics / Platnick N.I., Funk V.A. (eds.) Нью‑Йорк: Columbia Univ. Press. Vol. 2. P. 7-36.
Ghiselin M.T. 1974. Радикальне вирішення проблеми видів // Syst. Zool. Vol. 23. P. 536-544. Ghiselin M.T. 1981. Категорії, життя і мислення // Behavioral and Brain Sciences. Vol. 4. P. 269-313.
Ghiselin M.T. 1987. Концепції видів, індивідуальність і об’єктивність // Biology and Philosophy. Vol. 2. P. 127-143.
Gidley J.W. 1912. Lagomorpha, незалежний порядок // Science. Vol. 36. P. 285-286.
Gould S.J., Eldredge N. 1977. Пунктуальні рівноваги: темп і режим еволюції переосмислені // Paleobiology. Vol. 3. P. 115-151.
Gressit J.L. 1978. Еволюція ендемічних гавайських церамбіцидних жукiв // Pacif: Insects. Vol. 18. P. 137-167.
Hennig W. 1966a. Філогенетична систематика. Урбана: University of Illinois Press. 263 с.
Hull D.L. 1983. Карл Поппер і метафора Платона // Advances in cladistics / Platnick N.I., Funk V.A. (eds.). Vol. 2. P. 177-189.
Humphries C.J., Parenti L.R. 1986. Кладистична біогеографія. Oxford Monographs in Biogeography. Vol. 2. Оксфорд: Clarendon Press. 98 с.
Kieffer J. J. Bethylidae. 1914. Das Tierreich. Берлін, R. Fridlandar u. Sohn. Lief. 41. 595 с.
Kukalova‑Peck J. 1978. Походження і еволюція крила комах і їх зв’язок з метаморфозом, задокументовані викопним записом // J. Morphol. Vol. 15. P. 53-126.
Labandeira C.C., Beall B.S., Hueber F.M. 1988. Раннє різноманіття комах: докази з нижньодевонського щетинника з Квебеку // Science. Vol. 242. P. 913-916.
Laicharting J.N., von. 1781. Verzeichniss und Beschreibung der Tyroler Insecten. Bd. 1. Цюрих: Fuessley. xii+248 с.
Linnaeus C. 1751. Philosophia botanica in qua explicatur fundamenta botanica cum definitionibus partium, exemplis terminorum, observationibus rariotum, adjectis figuris aeneis. Стокгольм. P. G. Kiesewetter. viii + 362 с.
Mayr E. Toward a new philosophy of biology: observations of an evolutionist. Cambridge, London: Belknap Press, 1988. xi + 564 с.
McCune A.R. 1990. Еволюційна новизна і атавізм у комплексі Semionotus: розслаблений відбір під час колонізації розширюваного озера // Evolution. Vol. 44. P. 71-85.
Melville R.V. 1981. International code of zoological nomenclature. Results of vote on proposals for substantive amendments (fifth installment) // Bull. Zool. Nomencl. Vol. 38. P. 30-48.
Melville R.V. 1981. International code of zoological nomenclature. Deferment of proposal to introduce provisions to regulate paranomenclature // Bull. Zool. Nomencl. Vol. 38. P. 166-167.
Meyen S.V., Traverse A. 1979. Proposal on Article 3: remove «form‑genus» too // Taxon. Vol. 28. P. 595-598.
Mickevich M.F. 1978. Taxonomic congruence // Syst. zool. Vol. 27. P. 143-158.
Nelson G.J. 1978. Онтогенез, філогенез, палеонтологія і біогенетичний закон // Syst. Zool. Vol. 27. P. 324-345.
Olmi M. 1984. Revision of Dryinidae // Mem. Amer. Ent. Inst, Ann. Arbor. Vol. 37. Pt. 1‑2. 1913 с.
Osborn H.F. 1934. Aristogenesis: the creative principle in the origin of species // Amer. Natur. Vol. 68. P. 193-703.
Oster G., Alberch P. 1982. Evolution and bifurcation of developmental programs // Evolution. Vol. 36. P. 444-459.
Platnick N.I. 1979. Philosophy and the transformation of Cladistics // Syst. Zool. Vol. 28. P. 537‑546.
Plejel F., Rose G.W. 2002. Ceci n'est pas une pipe: names, clades and phylogenetic nomenclature // J. Zool. Syst. Evol. Research. Vol. 41. P. 162-174.
Poljansky G. I. 1977. Some aspects of the species in asexually reproducing Protozoa // Protozoology. Vol. 3. P. 17-23.
Queiroz K., de, Donoghue M.J. 1990a. Phylogenetic systematics or Nelson's version of cladistics? // Cladistics. Vol. 6. P. 61-75.
Queiroz K., de, Donoghue M.J. 1990b. Phylogenetic systematics and species revisited // Cladistics. Vol. 6. P. 83-90.
Queiroz K., de, Gauthier J. 1990. Phylogeny as a central principle in taxomony: phylogenetic definition of taxon names // Syst. Zool. Vol. 39. P. 307-322.
Rachootin S.P., Thomson K.S. 1981. Epigenetics, paleontology and evolution / Scudder G.G.E., Reveal J.E. (eds.) Evolution today. Піттсбург: Hunt Inst. P. 181-194.
Rasnitsyn A.P. 1981. Modified paranotal theory of insect wing origin // J. Morphol. Vol. 168. P. 331-338.
Rasnitsyn A.P. 1982. Proposal to regulate the names of taxa above the family group. Z.N.(S.) 2381 // Bull. Zool. Nom. Vol. 39. P. 200-207.
Rasnitsyn A.P. 1989. Vespida vs. Hymenoptera // Sphecos. Vol. 18. 8 с.
Rasnitsyn A.P. 1991. Typified names of higher taxa once again // Sphecos. Vol. 22. P. 5-7.
Rasnitsyn A.P. 1996a. Conceptual issues in phylogeny, taxonomy, and nomenclature // Contributions to Zoology. Vol. 66. P. 3-41.
Rasnitsyn A.P. 1996b. New Early Cretaceous Embolemidae (Vespida = Hymenopterea: Chrysidoidea) // Norden B.B., Menke A.S. (eds.) Contributions on Hymenoptera and associated insects dedicated to Carl V. Krombein. Memoirs of the Entomological Society of Washington. Vol. 17. P. 183-187.
Rasnitsyn A.P. 2000b (1999). Taxonomy and morphology of Dasyleptus Brongniart, 1885, with description of a new species (Insecta: Machilida, Dasyleptidae) // Russian Entomol. J. Vol. 8. No. 3. P. 145-154.
Rasnitsyn A. P. 2006. Ontology of evolution and methodology of taxonomy // Paleontological Journal. Vol. 40. Suppl. 6. P. S679‑S737.
Rasnitsyn A.P., Golovatch S.I. 2004. The identity of Phryssonotus burmiticus (Cocerell, 1917) (Diplopoda, Polyxenida, Synxenida) in Cretaceous amber from Myanmar // J. syst. Paleontol. Vol. 2. No. 2. P. 153-157.
Rasnitsyn A.P., Ross A.J. 2000. A preliminary list of arthropod families present in the Burmese amber collection at The Natural History Museum, London / Ross A.J. (ed.) The history, geology, age and fauna (mainly insects) of Burmese amber, Myanmar // Bulletin of The Natural History Museum, Geology Series. Vol. 56. No. 1. P. 21-24.
Rausch R.L. 1982. Cestodes in mammals: the zoogeography of some parasite‑host assemblages // Mem. Mus. nation. hist. natur. Ser. A. Vol. 23. P. 179-183.
Rotondo G.M., Springer V.G., Scott G.A.J., Schlanger S.O. 1981. Plate movement and island integration — a possible mechanism in the formation of endemic biotas, with special reference to the Hawaiian Islands // Syst. Zool. Vol. 30. P. 12-21.
Saether O.A. 1986. The myth of objectivity — post‑Hennigian deviations // Cladistics. Vol. 2. P. 1-13.
Saether O.A. 1979. Underlying synapomorphies and anagenetic analysis // Zoologica Scr. Vol. 8. P. 305-312.
Sarjeant W. A. S., Kennedy W. J. 1973. Proposal of a Code for the nomenclature of trace‑fossils // Canad. J. Earth Sci. Vol. 10. P. 460-475.
Schram F. R. 1983. Method and madness in phylogeny / Schram F. R. (ed.) Crustacean Phytogeny. Rotterdam: Balkema. P. 331-350.
Shear W.A., Bonamo P.M., Griedson J.D., Rolf W.D.I., Smith E.L., Norton R.A. 1984. Early land animals in North America: Evidence from Devonian age arthropods from Gilboa, New York // Science. Vol. 224. P. 492-494.
Shedrinsky A., Wampler Th.P., Grimaldi D. 2001. Burmese amber revisited (data on chemical composition of newly excavated material) / Krzeminska E., Krzeminski W. (eds.) Fossil insects. Second International Congress on Palaeoentomology, 5‑9 September, 2001, Krakow, Poland. Abstracts volume. Institute for systematics and evolution of animals, Polish Academy of Science, Krakow. P. 66-67.
Sher A.V., Kuzmina S.A., Kiselyov S.V., et al. 2006. QUINSIB. The Database on Quaternary Insects of Northeastern Siberia: Preliminary Version 3 (04.06).
Smuts J.C. 1987. Holism and evolution. Кейп‑Таун, N & S Press. 361 с.
Starobogatov Ya.I. 1991. Problems in the nomenclature of higher taxonomic catgories // Bull. Zool. Nom. Vol. 48. P. 6-18.
Sundberg P., Pleijel F. 1994. Phylogenetic classification and the definition of taxon names // Zoologica Scr. Vol. 23. P. 19-25.
Suno‑Uchi N., Sasaki F., Chiba S., Kawata M. 1997. Morphological stasis and phylogenetic relationships in tanpole shrimps, Triops (Crustacea: Notostraca) // Biol. J. Linnean Soc. Vol. 61. P. 439-457.
Tasch P. 1969. Branchiopoda. Treatise on Invertebrate Paleontology // Arthropoda. Vol. 4. 1. Pt. R. P. 128-191.
Tuxen S.L. 1963. Art‑ und Gattungsmerkmal bei den Proturen // Entomol. medd. Vol. 32. P. 84-98.
Tuxen S.L. 1967. Australian Protura, their phylogeny and zoogeography // Ztschr. zool. Syst. und Evolutionforsch. Vol. 5. P. 1-53.
Van Valen L. 1973. A new evolutionary law // Evol. Theory. Vol. 1. P. 1-30.
Van Valen L. 1976. Energy and evolution // Evol. Theory. Vol. 1. P. 179-229.
Van Valen L. 1982. Integration of species: stasis and biogeography // Evol. Theory. Vol. 6. P. 991‑112.
Wiley E.O. 1979. An annotated Linnean hierarchy, with comments on natural taxa and competing systems // Syst. Zool. Vol. 28. P. 308-337.
Wilson R.W. 1989. Rodent origin. // Papers on fossil rodents / Black C.C., Dawson M.R. (eds.). / Sci. Ser. Los Angeles Mus. Vl. 33. P. 3-6.
Woese C.R. 1987. Microevolution in the microscopic world // Patterson C. (ed.) Molecules and morphology in evolution: conflict or compromise? Cambridge, etc.: Cambridge University Press. P. 177-202.
Zherikhin V.V. 1999. Cladistics in palaeontology: problems and constraints. Proc. First Internat. Palaeoentomol. Conf., Moscow, 1998. AMBA projects Internat., Bratislava. P. 193-199.
Zherikhin V.V. 2002a. Insect trace fossils // Rasnitsyn A.P., Quicke D.L.J. (eds.) History of Insects. Dodrecht etc., Kluwer Academic Publishers. P. 303-324.
1.2.2.2. Фенетика
1.2.2.3. Філетика
А. П. Расницький. Теоретичні основи еволюційної біології // В. В. Жерихін, А. Г. Пономаренко, А. П. Расницький. Вступ до палеоентомології. М.: КМК. 2008. 371 с
1.2.2.3. Філетика (продовження)
1.2.3. ТАКСОНОМІЧНА НОМЕНКЛАТУРА
1.2.3.1. Таксономічно незалежні принципи
1.2.3.2. Таксономічно залежні принципи
1.2.3.2. Таксономічно залежні принципи (продовження)
1.2.3.3. Таксони, що порушують принципи номенклатури