
відновлений матеріал
1.2.3.2. Таксономічно залежні принципи
1.2.2.2. Фенетика
На відміну від кладизму, фенетика і традиційна систематика (або, у уточненому вигляді, філетика) виходять із цілісності живих систем і, відповідно, із дискретності біорізноманіття та зайняті пошуками гіатусів. Тому фундаментальна концепція повинна включати процес, що генерує таку дискретність. Спочатку дискретність легко пояснював креаціонізм, зараз на цю роль, схоже, претендує лише епігенетична теорія еволюції (див. вище).
З трьох конкуруючих підходів фенетика найпряміша у своїх цілях і засобах. Оскільки дискретність біорізноманіття описується у термінах схожості, а не родинності, то і задачу систематики, на думку фенетиків, слід формулювати в тих самих поняттях, а саме: система повинна явно будуватись у термінах схожості. Уникнути суб’єктивності тут можна за допомогою обчислення схожості. Методи розрахунку колективно називаються таксометриєю або частіше, але менш вдалим, нумеричною таксономією. На жаль, ці методи не були розвинуті до заявленого рівня — на мій погляд не через те, що цей рівень недосяжний, а тому що фенетика була витіснена кладизмом до того, як вона виконала свою корисну роботу. Дійсно, методи таксометриї численні і різноманітні, вибір методу, адекватного задачі, не простий і на нинішньому рівні ледь можливий без значної долі суб’єктивності. Проте, це проблема будь‑якого підходу, і кладизм тут не виняток (см., наприклад, Mickevich, 1978): виявити оптимальну область застосування конкретного методу, його сильні та слабкі сторони можна лише ціною значних зусиль. Перш ніж поступитися полем бою кладизму, фенетика значно, але недостатньо просунулася в цьому напрямку.
Однак корінна проблема фенетики, на мій погляд, інша. Схожість, з якою вона працює, це не зовсім та сама схожість, яку вимагає система організмів. Фенетики працюють з вивченими ознаками, кількість яких має бути розумно великою, але не більше того. Система ж, щоб оптимально виконувати свої функції, повинна спиратись на схожості та відмінності за всіма ознаками, вивченими і невивченими, включаючи і ті, які, можливо, ніколи не будуть вивчені. Звісно, така схожість недоступна прямому вивченню, але вона достатньо важлива, щоб спробувати оцінити її опосередковано. Саме цю мету і переслідує філетика.
1.2.2.3. Філетика
Філетика, як і кладизм, виходить із того, що організація живих істот визначається не лише і навіть не стільки умовами їх існування, скільки їх історією. Схожість історії (кровне родинство) у значній мірі передбачає і схожість організації. Різниця в тому, що методологія кладизму ігнорує все, крім родинства, фенетика ігнорує родинство, а філетика намагається врахувати і родинство, і схожість.
Для того, щоб філетичний таксон одночасно відображав і схожість, і родинні відносини, він має бути, по‑перше, фенетично (за схожістю) максимально однорідним і максимально відмінним від інших таксонів, по‑друге, монофілетичним. Монофілія розуміється у широкому сенсі порівняно з кладизмом, тобто включаючи голофілію (монофілію у кладистичному сенсі, см. Ashlock, 1971) і парафілію. Філогенетична кваліфікація таксонів визначається лініями походження, що перетинають межі таксона. Якщо його нижню межу перетинає єдина лінія (один корінь), таксон називається монофілетичним (у широкому сенсі), якщо їх більше — поліфілетичним. Точнішою, втім, буде інша формулювання: таксон слід вважати монофілетичним, доки і оскільки немає надійних свідчень його поліфілії (презумпція монофілії таксона).
Кладисти переозначили поняття монофілії, маючи на увазі не лише походження (корені) таксона, а й його подальшу долю. Якщо верхню межу монофілетичного (у нашому сенсі) таксона не перетинають філетичні лінії, тобто таксон не має явних нащадків, ми його називаємо голофілетичним, а кладисти — монофілетичним. Якщо ж його верхню межу перетнено (нащадки є), і для філетистів, і для кладистів це парафілетичний таксон. Кладисти називають голофілетичний таксон монофілетичним, оскільки з їхньої точки зору парафілетичний таксон точно так само не має права на існування, як і поліфілетичний. Вони не потребують терміну, що об’єднує голофілетичні та парафілетичні таксони: для них таке об’єднання беззмістовне. А для філетики такий термін абсолютно необхідний, оскільки там обидві ці форми таксона (голо‑ і парафілетичні) визнані і рівноправні. Саме таким, філетичним, було початкове розуміння терміна монофілія, воно ж прийняте і тут.
Існує ще ширше розуміння монофілії — як походження природного таксона кількома коренями від спільного предка у надрах предкового таксона (плюрифілія, пахіфілія, широка монофілія). Саме так зараз представляють походження ссавців, рептилій та багатьох інших великих груп: звідси і терміни мамалізація, артроподизація, ангіоспермізація тощо. Широка монофілія не лише підпадає під визначення поліфілії у кладистичній формулюванні, а й взагалі вичерпує реальне зміст цього терміна. За визначенням, за межами широкої монофілії залишається лише множинне походження таксона, природність якого заперечується (ми ще будемо мати справу з ними у розділі 1.2.3.3). Але для штучного таксона множинність його походження тривіальна, і навряд чи комусь спаде на думку це обговорювати спеціально. Мати окремий термін для такого випадку мені здається зайвим, тож я буду називати поліфілією всі випадки множинного походження таксона, як це і сформульовано вище.
Таким чином, виявляється, що багато (якщо не всі) добре вивчені таксони є поліфілетичними у зазначеному сенсі. Водночас важко сумніватися у природності рептилій, ссавців, членистоногих, покритоцвітних та багатьох інших таксонів, для яких більш‑менш переконливо продемонстровано походження більш ніж одним коренем від предкового таксона. Таким чином, поліфілія може виявитися нормою в таксономії. І все ж я вважаю, що методологічно правильніше виходити з презумпції монофілії. Тому я бачу дві причини: по‑перше, на нинішньому досить низькому рівні вивченості більшості таксонів виявлення множинності походження конкретної групи скоріше фальсифікуватиме гіпотезу її природності, ніж презумпцію монофілії. Якщо ж група дійсно природна, то додаткове дослідження, спровоковане виявленням поліфілії (см. нижче), лише підтвердить її природність і в кінцевому підсумку змусить визнати презумпцію монофілії несправедливою у даному конкретному випадку.
Існує інша причина, чому відмова від презумпції монофілії (у прийнятому тут розумінні, що виключає широку монофілію) здається методологічно небезпечною. Це означало б відмову від використання філогенезу для контролю природності таксона (см. нижче) і, відповідно, перехід до позиції фенетики. Таке рішення здається принаймні передчасним.
Повернемося до філетичної процедури. Філетичний таксон був визначений як монофілетичний континуум (Пономаренко, Расніцин, 1971). Визначення монофілії дано вище, континуум же розуміється як неперервний ланцюг, розгалужений чи ні, побудований із підпорядкованих таксонів (монофілетичних континуумів меншого об’єму) таким чином, що кожен з них фенетично ближчий до будь‑якого зі своїх безпосередніх сусідів по ланцюгу, ніж до членів будь‑яких інших таксонів (рис. 6). Таким чином, континуум відокремлений гіатусом (перервою неперервності схожості) від усіх інших континуумів.
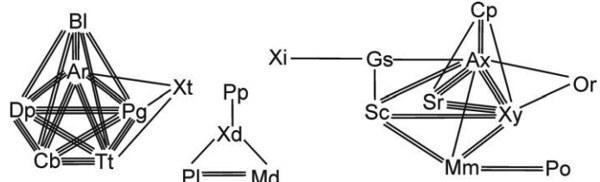
Рис. 6. Фенетична структура нижчих перепононокрилих комах на рівні родини, що демонструє континууми кількох ієрархічних рівнів (з Расницина, 19726). Лініями зображені рівні схожості: одинарною лінією — нижчий рівень із зображених, потрійною — найвищий. Скорочення: Аг — Argidае, Ах — Anахуелідае, В1 — Blasticotomidae, Сb — Cimbicidае, Ср — Серhidае, Dр — Diprionidае, Gs — Gidasiricidае, Мd — Меgаlоdontesidае, Mm — Myrmiciisае, Оr — Orussidае, Рg — Pergidae, Рl — Раmphiliidае, Ро — Раrоryssidaе, Рр — Раrараmphiliidае (нині в Sepulcidае), Sc — Sepulcidае, Sr — Siricidае, Tt — Tenthredinidае, Хd — Хyelydidае, XI — Хуеlidае, Xt — Xyelotomidae, Ху — Хiphydriidае.
Будувати систему, засновану на двох різних критеріях (у даному випадку це родинність і схожість), ризиковано тим, що допускає довільність у їх застосуванні — якщо лише сфери компетентності двох критеріїв чітко не розмежовані. Кладизм і фенетика вирішують проблему надзвичайно просто — відмовляючись використовувати той чи інший з двох критеріїв: кладизм ігнорує схожість, фенетика — родинність. Філетика діє тонше: вона використовує обидва критерії, але по‑різному. Схожість використовується як критерій, що задає систему, родинність — як критерій, що контролює якість системи. Таксон виділяється простежуванням гіатусів (розривів у схожості), а потім перевіряється на єдність походження.
Отже, філетика вимагає, щоб члени її таксона були достатньо схожі, щоб задовольнити критерій континууму, і при цьому не виявляли ознак поліфілії (походження від більш ніж однієї лінії предків). Якщо континуум виявляється поліфілетичним, критерії вступають у конфлікт. У цьому випадку систематик навряд чи відкине все, що було зроблено, і почне роботу спочатку. Його природна реакція буде спроба фальсифікації цього результату шляхом додаткового вивчення відповідної групи. Спроба зазвичай виявляється успішною в тому чи іншому сенсі. Наприклад, більш широкий аналіз може виявити, що схожість поліфілетичних груп поверхнева. Порядок сітковокрилих комах (Neuroptera) довгий час включав групи, близьке родинство яких згодом не підтвердилося. Так, ще Шарп (1910) об’єднував під цим іменем, окрім сітковокрилоподібних у сучасному розумінні, також ембіоптеру (Embioptera), терміти (Isoptera), сенокрил (Psocoptera), пухокрил (Mallophaga), стрекози (Odonata), веснянки (Plecoptera), піденки (Ephemeroptera), скорпіонниці (Mecoptera) і ручейники (Trichoptera). Подібна була доля ссавців порядку товстокожих (Pachydermata): таксон використовувався у XVIII‑IX ст. для об’єднання слонів, тапірів, носорогів, бегемотів і свиней, але потім був відкинутий на користь виділення більш природних (монофілетичних) порядків.
В інших випадках вдалося фальсифікувати гіпотезу поліфілії, як у випадку зайцеподібних. Їх виділили у свій час у окремий порядок Lagomorpha, ніби лише конвергентно схожих на гризунів (Gidley, 1912). Хоча статус порядку за зайцеподібними зберігся, за сучасними уявленнями вони утворюють з гризунами монофілетичну групу Glires (Wilson, 1989).
Цілком можливий випадок, коли обидві гіпотези (схожості і поліфілії) виявляться рівно правдоподібними. Швидше за все, це означатиме, що наша спроба була недостатньою, і треба спробувати заново проаналізувати ситуацію і усунути конфлікт. Але не можна виключати, що ми дійсно дійшли до реальної множинності коренів природного таксона, про яку ми вже неодноразово говорили. Як відрізнити одну ситуацію від іншої, у мене рецепту немає, проте на всяк випадок краще ще раз спробувати проаналізувати систему і філогенез групи на більш широкому матеріалі.
Існують і зовсім очевидні випадки, коли традиційна систематика зберігає беззаперечно поліфілетичні таксони, які не можуть бути без значних втрат замінені монофілетичними. Я маю на увазі різні паратаксони у таких галузях, як палеонтологія, паразитологія, мікологія, а також багато таксонів прокаріот (детальніше см. розділ 1.2.3). Існування паратаксонів ще раз підтверджує, що традиційна систематика в кінцевому підсумку орієнтується на схожість, а родинність використовується лише як спосіб опосередковано оцінити тотальну схожість (в тому числі за ще невивченими ознаками), яка недоступна прямій оцінці. Отже, родинність тут використовується приблизно так само, як у генніговій версії кладизму апоморфія використовується для встановлення дивергенцій, а у бразерської версії дивергенції слугують для визначення послідовності апоморфій.
Таким чином, усі розглянуті підходи однаково послідовні (моністичні, не еклектичні): генніговий кладизм у підсумку орієнтований на послідовність актів дивергенції, бразерський — на послідовність апоморфій, фенетика — на наявну (доступну безпосередньому вивченню) схожість, філетика — на тотальну схожість (недоступну прямому спостереженню, але опосередковано оцінювану за допомогою родинності).
Тепер ми можемо повернутися до питання про порівняльні переваги та недоліки кладизму і філетики. На мій погляд, найважливішою перевагою філетики є її кінцева опора на схожість, тобто на ознаки як такі. Вона може задовольнитися схожістю у наявних ознаках, якщо надійні свідчення родинності відсутні, і буде при цьому так само емпірична, як фенетика. Фактично це і буде фенетика. Але вона ж буде використовувати всі (і лише!) достатньо надійні філогенетичні дані, щоб підвищити надійність створеної нею системи, не роблячи її при цьому надто гіпотетичною. На відміну від філетики, кладистична система і безпосередньо, і в підсумку опирається на родинність, тобто на інтерпретацію ознак (на припущення щодо еволюційної послідовності станів ознак). У результаті кладистична система значно більш гіпотетична, ніж філетична.
Інша сторона сказаного полягає в тому, що філетика «всеїдна», а кладизм «вибагливий». Кладизм пред’являє значно жорсткіші вимоги до використовуваних ним ознак, ніж філетика (і фенетика), бо ознаки, що не піддаються надійній поляризації, марні. Це, правда, стосується лише геннігового кладизму (ручного, або «кладизму пера і паперу»). У комп’ютерному кладизмі можна використовувати будь‑які ознаки, але якщо програма не зможе впевнено реконструювати послідовність змін певної ознаки (а вона навряд чи зможе це зробити значно краще, ніж хороший систематик), то ця ознака буде зайвою у розрахунках.
Більше того: ознаки, добре працюючі в якій‑сь частині досліджуваної сукупності, але виявляючі численні паралельні зміни (гомоплазії) в інших її частинах, як ми бачили, девальвуються і враховуються лише в малій мірі — навіть там, де насправді вони філогенетично інформативні. Менш вивчені таксони, наприклад, вимерлі, охарактеризовані меншою кількістю ознак і тому часто не знаходять точного місця в комп’ютерних кладограмах, навіть якщо зберігають філогенетично важливі ознаки. У результаті кладисти змушені оголошувати паратаксонами («плезіонами») багато груп, розглянутих філетикою як нормальні (ортотаксони), але недостатньо відомих для точного охарактеризування їх родинних зв’язків з краще вивченими таксонами. Очевидно, це знижує таксономічну ефективність кладизму.
Філетична система, що враховує і схожість, і родинність, більш інформативна, ніж інші. Дж. С. Фарріс (Farris, 1979) у якості заперечення нагадує, що кладистична система у її найповнішій формі вказує походження всіх ознак і тому більш інформативна, ніж фенетична (і, відповідно, філетична). Цей висновок не безспірний, оскільки вичерпна повнота системи недосяжна. Дійсно, чим більше інформації ми набираємо про поляризацію відомих ознак, тим більше виявляється нових ознак, характер еволюції яких ще не відомий і які тому не можуть бути використані у «ручному» кладистичному аналізі. У результаті доводиться працювати з неповним набором ознак у порівнянні з філетикою, і в цьому сенсі система несе менший обсяг інформації. Комп’ютерний кладизм може використовувати всі доступні ознаки, але поляризацію трансформаційних серій він проводить автоматично, виходячи просто з розподілу станів ознак (за методом зовнішньої групи), тобто з досить обмеженої інформації. Усі інші знання про історію ознак, широко використовувані філетикою і ручним кладизмом, тут ігноруються, так що загальні втрати інформації великі.
Фактично будь‑який із порівнюваних таксономічних методів використовує лише частину доступної інформації, причому ці частини не сильно перекриваються. Комп’ютерний кладизм використовує обширне поле даних, але дуже вибірково і поверхнево. Фенетика діє подібно, але не практикує вибірковості. Ручний кладизм використовує обмежений «елітний» матеріал, відбір з необмежено широкої сукупності. Філетика також «просіює» необмежений обсяг вихідних даних, але вона вільніша, ніж «ручний» кладизм, у його сортуванні, і ніж фенетика і комп’ютерний кладизм, у його інтерпретації. Тому вона здатна до вилучення більш обширної та різноманітної інформації, ніж інші підходи.
Існує і інший аспект проблеми. З сказаного вище видно, що інформація закладена не в самій системі, а в її описі або, скоріше, у тому, що лежить в її підґрунті. Система як така містить лише дуже обмежену інформацію про топологію відповідної дендрограми і про довжину її міжвузлів, інтерпретовану в таксономічних рангах. Так, симетрична дихотомічна дендрограма про чотири кінцеві таксони без додаткової інформації про їх ранги містить три біти інформації і не більше. Деталізація ранжування таксонів збагачує інформативний вміст системи, і в цьому плані найбагатшими є оригінальні системи Геннига з їх неймовірною кількістю нумерованих рангів, що вичерпно відображають передбачувану послідовність актів дивергенції. Однак цей підхід був визнаний непрактичним, і його фактично залишено на користь традиційної лінеарної ієрархії рангів (Wiley, 1979).