Supports of Altruism. Column in ComputerreOnline #51
{"title":"","summary":"","body":"A characteristic feature of altruistic behavior in many animals, including humans, is that it concerns only individuals belonging to certain groups."}
Дмитро Шабанов
I ended my last column by noting that Simpson's paradox by itself is not enough for the development of true altruism. It can lead to an increase in the number of carriers of altruism genes only under fairly specific conditions. Nevertheless, right at the end of that column, I allowed for the presence of some additional factors that support selection in favour of altruism.
I will discuss such factors here. But before moving on to the main content of today's column, let me warn readers against a mistake that is often made. The column genre is obviously limited in length. Referring to some circumstance, I try to insert into the text the minimal explanations needed to understand it. This means that the full picture of some complex problem will not fit into a column, and I am forced to limit myself to discussing only one or a few interesting aspects of it. Please do not rush to conclude that if I have not mentioned some problem or referred to some phenomenon, I am denying its existence or importance, all right? For example, if I discuss the specifics of selection that favours the spread of genes for altruistic behaviour, that in no way means either that the phenomenon of altruism is fully explained by genes, or that I consider allele selection the sole mechanism of evolution.
Would you like a fuller picture than in this or the previous column? Read Markov. Here, by the way, is his presentation, which I am especially glad to link to. It is an expanded version of a talk at our Kharkiv conference of young scientists. We invited him a couple of years ago as a keynote speaker who opened the conference and led a round table.
Back to the subject! I am interested in the specific features of human populations that could have raised the effectiveness of group selection in favour of genes promoting altruistic behaviour. Here I will discuss three of them:
antagonistic relations between groups;
the non-random nature of group formation;
intragroup mechanisms for controlling individuals' behaviour.
I will try to reflect these factors in the model I built for the previous column. Of course, this will be a fairly conditional model; after all, I based its structure on the lifestyle of some hermaphrodites forming colonies on the sea floor. By introducing into this model factors reflecting the peculiarities of human populations, I am not claiming that it will correctly reflect our prehistory. Here the model will be used for something else. Its task is to show that the factors under consideration do indeed increase the effectiveness of group selection in favour of altruists, without providing quantitative estimates of such selection. Fortunately, models allowing quantitative assessment have already been created for human populations. Nevertheless, while writing this column and tinkering with the model, I came to a better understanding of certain features of our evolution. I hope that some readers will also expand their understanding with its help.
A characteristic feature of altruistic behaviour in many animals, including humans, is that it applies only to individuals belonging to certain sets. Grey rats selflessly support individuals from their own group and are extremely hostile toward members of other groups. People often divide those around them into "us" and "them", showing them completely different treatment.
Such altruism, limited to the bounds of a particular group, has come to be called "parochial altruism". The name is connected to the typology of political cultures by Gabriel Almond and Sidney Verba, American political scientists. They identified three types of culture: parochial, subject, and participant. Parochial altruism is part of parochial culture. In Russian-language literature (for example, in the writings of Alexander Markov) it is called not by a Russian word, but using transliteration - "parochial'nyi al'truizm". Sometimes Markov writes of "parochialism", meaning, apparently, precisely aggression toward outsiders. The use of transliteration has its pros (parishioners of any actual parish won't take offence) and cons (the term becomes opaque).
There is nothing here to take offence at. Parishes come in different kinds, but overall the link between supporting "one's own" and hating "outsiders" and belonging to particular religious communities has been shown for various human cultures. An interesting conclusion in the cited article is this: a tendency toward parochial altruism does not correlate with the frequency of prayer, but is significantly associated with the frequency of attending churches and communities. Of the six denominations studied, this effect was strongest among parishioners of the Russian Orthodox Church.
So, parochial altruism manifests itself simultaneously as love for one's own, ancestral group (which can be called patriotism) and as hatred of the foreign (xenophobia, chauvinism, and so on). There are serious grounds for supposing that genetic, biologically determined factors make a large contribution to the development of such behaviour. People prone to such behaviour are especially in demand during wars and conflicts. We remember them as heroes who combined selflessness, sincere devotion to brothers-in-arms, and hatred of enemies. In peacetime such psychology is far less adequate. That is one reason, in particular, why I am reserved about the attempts of patriotism propagandists to raise young people on the example of war heroes...
How can parochial altruism be reflected in my model? A consequence of antagonism between subpopulations is that some of them perish. If you haven't forgotten, on every step of my model seven subpopulations are formed at random. For them a group survival value was calculated, which was higher the more altruists there were in that subpopulation. I used a simple measure of a subpopulation's "strength": the product of its size and its survival value. All subpopulations whose "strength" is below the average level perish: they are destroyed by more numerous and more tightly knit competitors.
You know, this restructuring of the model did not lead to a substantial increase in the effectiveness of selection supporting altruism. At least, in the variant I chose, it is not only groups with a high share of altruists that win, but simply more numerous groups as well. This is realistic: large groups, even ones made up of less heroic fighters, have their own advantages. But if subpopulations form at random, the share of altruists in large groups turns out to be closer to the population average. Under such conditions, plain chance has a stronger effect on allele dynamics.
But is it always true that the ratio of egoists to altruists in subpopulations is determined at random? Doesn't it sometimes happen that the share of altruists in many groups systematically exceeds the level one would expect from a Poisson distribution? We should take into account the processes that lead to such deviations. The first is inbreeding within small groups. The second is assortativeness in group formation, that is, the presence of certain preferences when groups form.
The model reflects assortativeness, the ability of altruists and egoists to group "by interest". When subpopulations form, individuals are added to them one after another. The probability of adding individuals with genotype AA ("pure altruists"), aa ("pure egoists"), and Aa (neither one nor the other) depends on the frequencies of alleles A and a in the population. As the number of altruists in a subpopulation grows, the probability of adding further altruists to it increases (at the expense of a decreasing probability of adding Aa individuals); the same holds for the accumulation of egoists.
After the changes described, the effectiveness of group selection in supporting altruists increased sharply. But before showing the result, let me tell you about one more change. It is not only individuals that influence the properties of the group they belong to. Groups, too, influence the behaviour of the individuals that make them up. Let me give two examples of such influences. The first. In colonies of social insects, worker individuals, who are not supposed to reproduce, keep watch over one another. If, for example, a worker wasp selfishly starts laying her own eggs (instead of altruistically tending the queen's eggs), her brood will be destroyed by the other workers.
The second. In groups of people where it is customary to help one another, an egoist who refuses to follow the accepted norms of behaviour may experience social condemnation that forces them to change their behaviour. Recall: in the previous column I mentioned reciprocal (mutual) altruism. One consequence of it is the ability of individuals to assess each other by their willingness to help those close to them. A consequence of this mechanism's operation can be the phenomenon of reputation. Individuals who act altruistically earn a good reputation, which helps raise their status (and, for example, increase the number of their descendants). In such groups, not only carriers of genes promoting altruistic behaviour but also genetic egoists may behave altruistically!
In my model this is reflected as follows. After a subpopulation forms, the ratio of "pure altruists" (AA) to "pure egoists" (aa) within it is determined. If there are more altruists, Aa individuals behave as altruists (both in terms of the group's viability and in terms of their own chances of survival); if there are more egoists, they behave as egoists. When the numbers of individuals from the AA and aa groups are equal, the characteristics of representatives of the Aa genotype turn out to be intermediate between the two extremes.
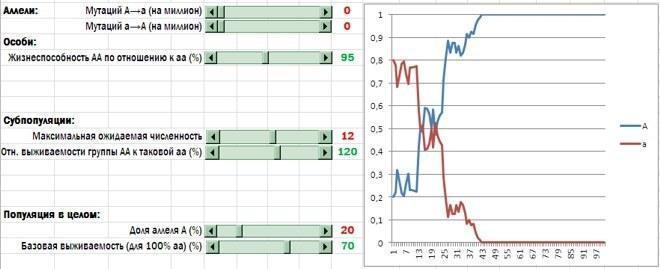
So, I have introduced into the model analogues of group antagonism, assortative group formation, and the action of intragroup behavioural norms. The stage of asexual reproduction of the remaining individuals was no longer needed, and I removed it. Here is the modified (and significantly simplified!) model. Although it describes a different mode of population reproduction than the one characteristic of humans, what matters to us is that the changes made strengthened selection in favour of altruists.
Compare this screenshot with the figures in the previous column. The changes described here significantly simplified the model and strengthened support for altruism. Now altruists systematically win even at values of the model's initial parameters that would otherwise have left them almost no chance
By the way, the considerations set out here can help not only in understanding the evolution of altruism. Among other things, they also help in searching for answers to other questions, such as this one. Why are there many religions, and why are they different?
One could say that all people reach toward one and the same truth, but find it for themselves in different ways. Another (widely held!) version holds that there is one true faith, and all the others are the result of devilish temptation leading heretics astray from the true path. I like the explanation that regards the existence of religions and other collective rituals as an essential part of the mechanism that created us in the course of evolution.
And look what follows. The factors we have examined reinforce one another's action. Antagonism between groups leads to the more successful groups being those united by parochial altruism; parochial altruism strengthens antagonism between groups. Non-random group formation and the emergence of mechanisms supporting group cohesion favour the victory of subpopulations united by parochial altruism. As the evolution of altruism proceeds, ever more tightly knit groups win out in intergroup competition. Devotion to the faith of one's fathers is part of the mechanism that drove our evolution. But blind faith has evil eyes...
The complex of mechanisms we have examined is bound together by positive feedback loops. One of the interesting features of positive feedback loops is that their action, sooner or later, moves the system under consideration into a new state. Once evolution by the mechanism described here began in some group of animals, the process of directed change within it was bound to become self-sustaining. To an outside observer it would seem directed toward a certain outcome.
And what do we have to do with this? We - are that very outcome...