
Shyshkin, 1987. Індивідуальний розвиток і еволюційна теорія
Shyshkin M.A. Індивідуальний розвиток і еволюційна теорія // Еволюція та біоценотичні кризи. М.: Наука, 1987. – С. 76–124.
Шишкін М. А. Індивідуальний розвиток і еволюційна теорія // Еволюція і біоценотичні кризи. К.: Наука, 1987. - С. 76–124.
УДК 591.3:575.8
ІНДИВІДУАЛЬНИЙ РОЗВИТОК І ЕВОЛЮЦІЙНА ТЕОРІЯ
М. А. Шишкін
Палеонтологічний інститут АН СРСР
Дарвінова теорія природного відбору має своїм об’єктом цілісні живі організми, що виникають у ході індивідуального розвитку і ведуть боротьбу за існування протягом усього життєвого циклу. Тому вирішення ключової для цієї теорії проблеми походження адаптації неминуче вимагає з’ясування законів еволюції онтогенезу, тобто законів становлення тих механізмів, які забезпечують виду надійне здійснення його фенотипічної норми. З позицій сьогоднішніх знань можна бачити, що багато заперечень, що висувалися проти класичного дарвінізму, могли здаватися суттєвими лише тому, що питання еволюції індивідуального розвитку залишилися в цьому вченні не розробленими.
Зовсім іншу основу має домінуюча нині генетична («синтетична») теорія еволюції, що означає, за висловом І. І. Шмальгаузена (1968а, с. 20), заміну дарвінізму генетикою. Абстрагування від проблем онтогенезу лежить у самих її передумовах, оскільки матеріалом відбору тут по суті уявляються не організми, а успадковані фактори або їх комбінації, тобто структури, що передують розвитку; їхня доля і має визначати хід еволюції. Питання про вплив фенотипічної реалізації цих факторів на характер відбору залишається осторонь, якщо не враховувати того, що береться до уваги можливість їх проявлення або непроявлення і кількість теоретично визнається впливом на ці процеси генотипічного середовища. Таке становище достатньо добре ілюструється тим, що до недавнього часу можна було зустріти роботи з викладом генетичної теорії, у яких поняття онтогенезу взагалі не вживалося (Шеппард, 1970). З іншого боку, характерно, що множаться в останні роки критичні висловлювання на адресу цієї теорії, які в першу чергу підкреслюють спрощеність і недостатність її передумов для розуміння еволюційної ролі процесів розвитку.
Між тим історія дарвінізму знає дві серйозні спроби зрозуміти взаємовідносини між природним відбором і індивідуальним розвитком — теорію зароджувальної плазми А. Вейсмана та вчення про стабілізуючий відбір І. І. Шмальгаузена і К. Уоддінгтона. Запропоновані ними рішення діаметрально протилежні. Якщо у Вейсмана еволюційні зміни онтогенезу становлять лише пасивний результат відбору елементів зароджувальної плазми, то у Шмальгаузена, навпаки, матеріалом для селективного перетворення генотипу служать аберації онтогенезу. Генетична теорія успадкувала від Вейсмана лише його редукціоністське розуміння успадкованості — розчленування її на незалежні фактори (типове для теорій успадкованості XIX ст.) і заміну останніх цілими організмами як субстратом відбору. Головний же об’єднуючий принцип вейсманівської концепції, що зробив її справжньо синтетичним будуванням, залишився непоміченим. Він полягав у ясному розумінні того, що механізм успадкування властивостей організму виражається у способі їх онтогенетичного здійснення, і тому визнання передачі цих властивостей незалежними носіями може означати лише їх незалежну реалізацію в онтогенезі. Іншими словами, ідея дискретної успадкованої детермінації неминуче передбачає мозаїчний (передзформований) розвиток. Вейсман побудував таку модель розвитку (опираючись на гіпотезу нерівномірного успадкування), і саме її несумісність логічно призвела до краху всієї його теорії. Однак висновки, здавалося б, що випливали звідти для розуміння механізму успадкованості, так і не були зроблені до появи теорії стабілізуючого відбору.
Що стосується останньої теорії, що аналізується далі, то ставлення до неї сучасного селекціонізму невизначене. Хоча в вітчизняній літературі її часто характеризують як найвищий сьогоднішній досягнення дарвінізму (Шварц, 1969; Галл, 1980), все ж, як правильно зазначають (Кирпичников, 1974), її визнання часто має чисто словесний характер. Причини цього зрозумілі в значній мірі. Теорія стабілізуючого відбору зі своїм спрямуванням на пояснення еволюції цілісної організації є успадкованою щодо класичного дарвінізму (ср.: Яблоков, 1981), але не щодо генетичної теорії. Їхні відносини з останньою, як буде показано нижче, є в дійсності антагоністичними. Тому для домінуючого напряму сучасного селекціонізму неможливо прийняти вчення про стабілізуючий відбір по суті, не ревізуючи при цьому своїх власних основ.
Якщо задовільна еволюційна теорія має бути по суті теорією еволюції онтогенезу і якщо дарвінове вчення містить можливість для цього, то його основоположні поняття мають бути виражені мовою індивідуального розвитку. Це стосується успадкованості, змінливості, пристосованості (як процесу), а також самого механізму природного відбору. Тому ж аналізу підлягають і поняття генетики, що використовуються сучасним еволюціонізмом, якщо описувані ними явища мають відношення до реалізації фенотипів і їх адаптації. Йдеться про те, щоб з’ясувати, чи дійсно те, що називається алелями, гомо‑ і гетерозиготами тощо, є «чистими» характеристиками успадкованої структури, що становить субстрат відбору, чи ж ми маємо справу з особливостями морфогенетичних систем, створених самим еволюційним процесом.
Перш ніж перейти до цих питань, необхідно виділити деякі найзначиміші в еволюційному плані особливості індивідуального розвитку. Еволюція — це, передусім, зміна поколінь, що веде до зміни адаптивних норм, і перехід до кожної нової норми неминуче означає те чи інше порушення попередньої. Тому в еволюції популяцій і видів мають існувати покоління з переважаючими стабільними за своїм результатом індивідуальними циклами (формуючими норму) і покоління, що характеризуються онтогенезами з більш невизначеним результатом. Закономірності переходу від другого типу до першого мають пояснювати механізм адаптаціогенезу (як це трактується теорією стабілізуючого відбору). Отже, в онтогенезі нас в основному цікавитиме співвідношення стабільних і нестабільних, або нормальних і аберантних, шляхів розвитку.
НАСЛІДКОВІСТЬ, ЗМІНЛИВІСТЬ, ПРИСТОСОВАНІСТЬ ТА ОРГАНІЗОВАНІСТЬ ЯК ХАРАКТЕРИСТИКИ СТАЛІСТІ ІНДИВІДУАЛЬНОГО РОЗВИТКУ
Характерною властивістю живих істот є стійкість їх типічної морфофізіологічної організації, позначуваної як адаптивна норма (Шмальгаузен, 1940а). Це, власне, і робить можливим існування таких типологічних методів дослідження, як порівняльна морфологія, систематика і філогенетика. У ряді поколінь ця властивість організмів проявляється, крім того, як їх здатність до самореалізації, тобто як стійкість успадкування адаптивної норми. Ці дві особливості іноді вказують як самостійні характеристики живого (Кастлер, 1967), але в дійсності вони мають одну і ту ж основу — стійкість здійснення норми в ході індивідуального розвитку.
Щодо нормально організованої структури ця основа самоочевидна, оскільки дорослий стан організму, про який йдеться, сам є складовою частиною і підсумком онтогенетичного циклу. Так само очевидний і зв’язок між розвитком норми і її успадкуванням, проте тут потрібні деякі роз’яснення. У дарвінській теорії еволюції успадкованістю називається передача фенотипічних ознак у поколіннях (Дарвін, 1951). Це «передача» означає, звичайно, відтворення ознаки заново в процесі індивідуального розвитку, що поєднує у типічному випадку гамети батьків з фенотипом нащадка; надійність результату цього процесу і визначає успадкування. Тому, говорячи про «успадковані» і «набутих» ознаки, фактично мають на увазі лише стійкість або лабільність їх онтогенетичної реалізації щодо умов розвитку, і ці поняття переважні, оскільки значно ясніше передають сутність справи (Вуджер, 1953; Шмальгаузен, 1969, 1982; Шишкін, 1981, 1984а, б, в). Словами А. Г. Гурвіча (Gurwitsch, 1912; Гурвіч, 1944) успадкованість — це процес здійснення типового розвитку (ср.: Майстер, 1934; Дубінін, 1973). Іншого змісту щодо фенотипів (для яких цей термін і введено) термін «успадкованість» не має. Позначати ним генетичну обумовленість ознак безглуздо, оскільки всі властивості організму є продуктами взаємодії генотипу і середовища розвитку і в такому випадку одночасно є «успадкованими» і «набутих» (Йогансен, 1926; де Беер, 1963; Камшилов, 1972). Якщо ж віднести поняття успадкованості до генотипів і їх елементів, то не лише змінюється його первинний сенс, а й воно просто перетворюється у «річ у собі», що не піддається іншому визначенню, крім тавтологічного («успадкованість — це передача успадкованих факторів») [Поширене твердження, що об’єктом успадкування є індивідуальна «норма реакції» (Йогансен, 1926; Добжансі, 1947) рівносильно визнанню успадкування індивідуальних генотипів. Але останні безперервно перетворюються в кожному поколінні за рахунок помилок реплікації (у загальному випадку) або в ході рекомбінації (при статевому процесі)]. Цей важливий момент треба уточнити, оскільки вся історія вивчення успадкованості несе на собі відбиток дивного на перший погляд дуалізму, коли з одного боку в ній бачать властивість певної категорії ознак, а з іншого — щось, що існує незалежно від них і належить до зароджувальної плазми (генотипу). Це протиріччя зникає лише в одному випадку — якщо вважати, що стійке відтворення (успадкування) ознак обумовлене передачею через гамети однозначно відповідних їм дискретних причинних факторів. Саме так і вважала рання генетика, і це уявлення об’єктивно залишається фундаментом побудованої на ній еволюційної теорії, незважаючи на прагнення її авторитетів (наприклад, Майр, 1968, 1974) вважати його застарілим.
Твердження, що стійкість організмів створюється селективним процесом, є центральним для теорії стабілізуючого відбору. Але якщо онтогенетична стійкість означає успадкованість, то очевидно, що і остання є продуктом відбору. Цей висновок чітко формулюється зазначеною теорією у вигляді понять про перетворення лабільних змін у успадковані, про створення відбором успадкованих механізмів, заміну зовнішніх факторів розвитку внутрішніми (Шмальгаузен, 1940б, 1941, 1968б, 1982, с. 109, 110, 161, 214) і, нарешті, про уявлення про генетичну асиміляцію адаптивних ознак (Waddington, 1953, 1957). Між твердженням, що успадковані зміни створюються відбором, і звичним уявленням, що вони створюються мутацією, лежить непрохідна світоглядна пропасть, яка має зупинити кожного, хто хотів би узгодити сутність вчення Шмальгаузена‑Уоддінгтона з генетичною теорією. Роздумуючи над змістом більш звичного другого твердження, якщо мова йде просто про генетичні зміни, їх пояснення мутаціями є тавтологією. Якщо ж маються на увазі зміни фенотипу, стійко збережені в поколіннях, це завідомо неправильно. З досвіду експериментальної генетики добре відомо, що елементарні малі мутації не гарантують стійкого аффекту (їх прояв завжди лабільніший норми) і, більш того, можуть взагалі не проявлятися. Збереження фенотипічної норми при безперервній рекомбінації генотипу в ксеногамних популяціях особливо добре ілюструє останнє. Таким чином, успадкованість (стійкість) і генетична обумовленість — це різні речі (ср.: Камшилов, 1967).
Все сказане неминуче веде до переоцінки іншого звичного постулату генетичного селекціонізму про «успадковану змінливість» як матеріалу природного відбору. Фактично він також базується на ототожненні ознак і їх успадкованих факторів. Він передбачає, що елементарні (неадаптивні) фенотипічні варіації особин мають успадковуватись, якщо вони обумовлені їх генетичними різницями. Але насправді, як уже сказано, наявність генетичної гетерогенності особин сама по собі нічого не говорить про характер її фенотипічного вираження. Вона може або взагалі не проявлятись (під покровом нормального фенотипу, см.: Четвериков, 1926), або проявлятись нестабільно (як типово для мутацій). Теорія стабілізуючого відбору, виходячи з цих фактів і розглядаючи саме властивість успадкування як продукт еволюції, природно повинна вважати сирим матеріалом останньої нестабільними ознаками, тобто лабільними елементарними реакціями (морфозами) індивідуально різних геномів (Шмальгаузен, 1982; Шишкін, 1981, 1984а, б). Саме сукупність таких реакцій, що здійснюються на гетерогенній основі, відповідає дарвінівському поняттю невизначеної змінливості, яке дозволяє говорити про зміни, не лише неадекватні факторам середовища, а й неупорядковані за характером відтворення у найближчого нащадка.
Еволюційне походження біологічної стійкості можна розглядати ще в одному аспекті. Саме це поняття має багато синонімів, що з давніх часів вживаються саме для позначення явищ, що потребують еволюційного пояснення. Серед них — цілеспрямованість, пристосованість (Ешбі, 1959, 1962; Шмальгаузен, 1968а, с. 139) і збалансованість із середовищем (Спенсер, 1899).
Усі вони означають властивість індивідів реагувати на зовнішні збурення таким чином, щоб зберігати свою нормальну життєздатність, включаючи успішне розмноження. Адаптивна організація — це організація, здатна до персистування (Wake et al., 1983). Історичне виживання найпристроєніших означає збереження і створення відбором все більш стійких типів організації, здатних протистояти максимально широкому спектру збурень. Чим ширший і різноманітніший цей спектр, тим більше нейтралізуючих реакцій потрібно організму, щоб у підсумку він міг реалізувати один із допустимих для нього ізоморфних нормальних станів (закон необхідного різноманіття; Ешбі, 1959). Ці реакції мають бути скоординовані, бо стійкість системи неможлива без взаємодії її частин (Bertalanffy, 1969). Таким чином, зростання пристосованості (стійкості) у ході відбору неминуче веде до ускладнення і підвищення інтегрованості морфофізіологічної організації. Цей дарвінівський принцип, постійно оскаржуваний, починаючи від К. Негелі і до сучасних еволюціоністів (Wright, 1964), логічно випливає з розгляду організму як цілісної системи; проте він і справді стає необґрунтованим, коли ми намагаємося замінити організми як об’єкти відбору мозаїкою їх успадкованих факторів.
Усі види, оскільки вони володіють адаптивною нормою, однаково пристосовані до свого середовища існування (тобто до свого спектру допустимих збурень) і, відповідно, рівнозначні в тому якості, яку можна назвати їх відносною стійкістю. Однак їх можна, принципово, порівняти і за абсолютною стійкістю, тобто за ступенем того різноманіття зовнішніх факторів, ефект яких вони здатні релаксувати. Цей показник, як видно з викладеного, є мірою їх організованості, тобто і мірою прогресу. Чим більш хаотичні і непередбачувані коливання факторів середовища існування, тим вищі вимоги до складності самого організму, і навпаки, чим середовище однорідніше, тим нижчі. Прості (передбачувані) коливання високоупорядкованих внутрішньоорганізмових середовищ, що використовуються паразитами, пояснюють їхню тенденцію до дегенерації.
Оскільки встановленням фізіологічної поведінки організмів є регуляція у бік норми, то вона спрямована на протидію незворотним змінам, які і становлять сутність еволюції. Стійка система (справжньо рівноважна, або квазірівноважна, якою є живі організми і взагалі відкриті системи), доки вона такою залишається, за визначенням, «не пам’ятає» своїх флюкцій (модифікацій) і, отже, не еволюціонує. Причина еволюції лежить у порушенні стійкості (Спенсер, 1899), тобто у виході значень динамічних змінних системи за межі, що дозволяють регуляцію цілого. Відновлення стійкості на новому рівні (тобто нового рівноваги зі зміненим середовищем) відбувається лише за допомогою природного відбору, який у зв’язку з цим, строго кажучи, завжди є стабілізуючим. Він являє собою механізм надорганізмової регуляції індивідуальної стійкості.
Ідеальна стійкість, тобто здатність реагувати флюкцією на будь‑який зовнішній чи внутрішній вплив, залишається, звичайно, недосяжною для організмів, але чим вищий її абсолютний показник, тим менш вони вразливі до прямої елімінації (що все більше поступається місцем диференціальній розмноженню) і тим більше зростає їхня здатність передбачати будь‑яку об’єктивно необхідну елементарну еволюційну зміну відповідною адаптивною модифікацією, тобто частково змоделювати її з «наявних можливостей» своєї морфогенетичної системи. У цьому сенсі твердження Г. Спенсера (1899) про те, що в процесі еволюції органічного світу природний відбір поступається місцем прямому пристосуванню; хоча це і не зовсім точно, сама тенденція тут зрозуміла правильно. У цій же можливості передвісного моделювання полягає і сутність ідеї «органічного відбору» Моргана‑Болдюїна, що обґрунтовує еволюційну роль модифікацій (Шишкін, 1984б).
Оскільки зростання абсолютної стійкості, або пристосованості, пов’язане з ускладненням організації, тобто рухом до все менш ймовірного стану, еволюція віддаляє організми все далі від термодинамічного рівноваги, що можливо лише за рахунок все вищого рівня споживання енергії зовні. Таким чином, зростання організованості (стійкості) пов’язане зі збільшенням енергетичних витрат, і швидкість продукції ентропії є її суттєвим показником (Goodwin, 1970).Ці витрати окупаються тими самозрозумілими перевагами, які дає висока пристосованість у боротьбі за існування.
УСТОЙКІСТЬ НОРМИ ТА ПРИНЦИПИ ТЕОРІЇ НОРМАЛЬНОГО РОЗВИТКУ
Устойчивість розвитку типового організму, що забезпечує його самопідтримання (пристрастність) і самореалізацію (спадковість), а також зв’язок цього явища з регуляторними механізмами онтогенезу відзначені ембріологією давно. Уже К. Бе́р (Ваер, 1828) порівняльним шляхом встановив зменшення ембріональної змінливості в ході розвитку, що показує, що властивістю останнього є устремленість до певного кінцевого стану. Для механіки розвитку, що виникла в кінці XIX ст. і поставила собі завдання експериментальне встановлення та локалізацію безпосередніх діючих факторів морфогенезу, ця властивість виявилася непереборною перешкодою при спробах побудувати загальну теорію розвитку на основі каузально‑аналітичного методу (Бляхер та ін., 1935).
Результат процесу не піддавався інтерпретації як сума ефектів певних початкових причин, демонструючи значну незалежність від їх варіацій. Устойчивість цього результату щодо способів його досягнення (Roux, 1895), продемонстрована багатьма експериментами, зробила неможливим прийняття мозаїчної неоперформістської концепції В. Ру, що зводить розвиток до незалежної диференціації початкового набору зачатків. Теоретичні наслідки з явищ саморегуляції розвитку та принципової нерозкладності останнього на незалежні причинно‑наслідкові ланцюги були усвідомлені Г. Дрішем (Driesch, 1908, Дриш, 1915), який охарактеризував зародок як «гармонійну еквіпотенціальну систему», тобто комплекс частин з однаковими можливостями (проспективними потенціалами), керований як ціле у своїх перетвореннях внутрішнім нематеріальним упорядковуючим фактором — ентелехією. Цей нерозкладний фактор визначає фактичну долю (проспективне значення) зачатків у відповідності до їх розташування в цілому і контролює узгодженість їх змін протягом усього розвитку, зокрема й на етапах самодіференціації, коли експериментально між зачатками не виявляються взаємозалежності. У цій концепції важливо, звичайно, не конкретне «рішення» проблеми (яке просто замінюється символом ентелехії), а чітке розуміння того факту, що розвиток — це цілісний процес, властивості якого надсумативні і стійкі щодо його складових елементів. Це стало початком системного підходу до розвитку, в основі якого лежить аристотелівський принцип «ціле існує раніше частин». Його правомірність була показана ще раніше в такому суто емпіричному узагальненні, як закон Бера (Ваер, 1828), який буквально стверджує те саме (загальне в розвитку виникає раніше специфічного); але лише з Дрішем він став використовуватися як основа теорії розвитку. На ньому будується сучасне вчення про ембріональну детермінанцію, яке, проте, на відміну від теорії Дріша визнає фактор цілісності матеріальним і пізнаваним (Gurwitsch, 1910, 1912; Світлов, 1964, 1978; Белоусов, 1963).
Отже, пошуки причинного пояснення стійкості нормального розвитку привели ембріологію до розгляду цього процесу як ієрархічної системи (ціле і його частини), керованої своїм верхнім рівнем, тобто властивостями цілого. Пізніше (і по суті на трьох самих основах) було отримано і історичне пояснення зазначеної властивості розвитку. Ми знову маємо на увазі теорію стабілізуючого відбору (Шмальгаузен, 1940б, 1941, 1968б, 1982), що спирається на поняття адаптивної норми. Остання (тобто нормальний фенотип) історично змінюється набагато повільніше, ніж спосіб її онтогенетичної реалізації, який безперервно перетворюється відбором у бік підвищення його надійності. Таким чином підвищуються можливості варіювання процесів, що складають розвиток, без шкоди для стійкого здійснення норми. Остання виступає тут як фактор цілісності, що керує (через відбір) зміною своїх морфогенетичних механізмів і визначає допустимий простір індивідуального варіювання їх елементів (Шишкін, 1981). Принципове тотожність ембріологічного і історичного пояснень стійкості здається тут самозрозумілою.
Нас, однак, цікавить спочатку ембріологічне пояснення, тобто схема причинно‑наслідкових відносин, придатна для опису окремого циклу розвитку. Але спершу необхідно зупинитися на загальних теоретичних вимогах, яким має задовольняти цей опис.
Стійкість результату нормального розвитку означає цілеспрямованість цього процесу. Обидва ці визначення характеризують одне й те саме — здатність до саморегуляції кінцевого стану. Цілепокладана (телеономічна) поведінка стійкої матеріальної системи проявляється в тому, що, будучи виведена з стану рівноваги, вона реагує так, що в кінцевому підсумку повертається до нього. Відповідно для опису таких процесів у фізиці і хімії використовуються фіналістичні формулювання (принцип Ле‑Шательє тощо). Для закритих систем стан стійкості відповідає термодинамічній рівновазі, а в відкритих, включаючи живі організми, знаходиться на віддаленні від неї і характеризується як «потокове рівновагу» (Bertalanffy, 1949), або «стійке неравновагу» (Бауер, 1935), або ж як стаціонарний стан. Рух до будь‑якого типу рівноваги, або «пошук цілі», здійснюється через замкнені цикли подій з зворотним зв’язком, коли, наприклад, елемент А при збуренні впливає на Б таким чином, що останній своїм зміною коригує стан А у бік значення, що зменшує подальшу його корекцію. Система «керується своєю помилкою» (Ешбі, 1962), і її стійкість базується на взаємодії її елементів. В відкритих системах цикли корекції здійснюються постійно; у закритих же амплітуда взаємодій згасає зі зростанням ентропії і встановлюється «стійкість щодо точки» (Goodwin, 1970).
Уявлення про цілеспрямовану поведінку системи не означає, звичайно, визнання залежності подій від майбутніх умов. Воно лише відображає той факт, що кінцеві результати елементарних змін у системі визначаються загальними властивостями самої системи і не можуть бути зведені до прямих механічних наслідків цих змін. Система як ціле або взагалі не реагує на елементарний вплив, або переходить у один зі своїх альтернативних станів (модифікацій). Іншими словами, телеономічна залежність виявляється при порівнянні подій або властивостей, що відповідають різним ієрархічним рівням системи, а саме при порівнянні її повільно змінних параметрів (що характеризують її цілісну поведінку) і швидко варіюючих значень її елементів (динамічних змінних). Фіналістична форма опису таких співвідношень відображає принципову неможливість їх каузального опису, бо властивості цілого не зводяться однозначно до станів його елементів. «Те, що сприймається як стійка структура певного рівня, насправді утримується безперервним обміном компонентів найближчого нижчого рівня» (Bertalanffy, 1969), тобто одна і та сама властивість цілого зберігається при різних комбінаціях елементарних взаємодіючих причин. Іншими словами, для міжрівневих відносин характерна різка асиметрія причин і наслідків (Белоусов, Чернавський, 1977), не притаманна процесам, що піддаються каузальному опису. [З цієї точки зору віталістична концепція розвитку Дріша виступає як характерна реакція дослідника, що констатує відсутність звичної для нього однозначної причинної залежності між пов’язаними явищами і не бачить іншої альтернативи для опису їх зв’язку, окрім введення нематеріальних факторів]. Тому не можна погодитися з поширеним уявленням, що фіналістичні та каузальні формулювання — це лише два рівноправних способи опису змін у тих самих циклічних причинних ланцюгах. Ієрархічно рівноправні елементи таких циклів не мають асиметричних співвідношень між собою і, навпаки, властивості різних рівнів системи не пов’язані каузальною залежністю.
Все сказане має пряме відношення до розуміння механізму індивідуального розвитку. Ми приходимо до висновку, що його каузальне пояснення можливе лише в тому випадку, якщо весь ланцюг причинних подій, що ведуть до цілісного кінцевого результату (нормальної організації), буде представлений як послідовність рівноправних (одно‑рівневих) цілісних станів. Наявність у ході розвитку ланцюга таких станів зі стійкими характеристиками є не лише теоретичною вимогою, а й експериментально встановленим фактом. Тут дійсно виявляються послідовні періоди, що характеризуються внутрішньою цілісністю (топологічною ізоморфністю) і спрямованістю перетворень, при зниженої чутливості до експериментальних порушень (Світлов, 1960; Белоусов, 1979). Водночас індивідуальний розвиток, так само як і будь‑яка інша незворотна зміна, має бути пов’язаний з переходом через фази порушення стійкості (Bertalanffy, 1969; Волькенштейн, 1981б). Ці фази також виявляються в індивідуальному розвитку у вигляді «чутливих періодів», що характеризуються лабільністю детермінанції і трансформацією топологічних рисунків. Отже, загальна теорія нормального онтогенезу повинна мінімально включати в себе такі передумови.
1. Розвиток — це ланцюг взаємозалежних структурно цілісних станів. 2. Кожен з них протягом свого існування визначає хід і узгодження окремих морфогенетичних процесів (тобто діє як «ентелехія» за Дрішем). 3. Реалізація цих процесів кожного разу має на результат певне порушення стійкості цілого і його відновлення потім на новому рівні, що контролює подальшу диференціацію. 4. Оскільки в ході розвитку організація зародка ускладнюється, кожен новий стан цілісності стабілізується на все більшій відстані від істинної рівноваги.
Ймовірно, єдиною концепцією розвитку, що узгоджується з цими передумовами, є сьогодні теорія біологічного поля, запропонована А. Г. Гурвичем (Gurwitsch, 1922; Гурвич, 1944). Поняття фізичного поля, тобто простору, властивості якого визначають поведінку часток у ньому, добре відповідає уявленню про шуканий матеріальний фактор цілісності, що контролює всю сукупність процесів розвитку. Як правильно зазначив П. Г. Світлов (1964), принцип поля ясно виражений уже в концепції Дріша, який вказав, що проспективне значення окремого елементу в розвитку є функцією його розташування в цілому [Це уявлення Дріша вступало в очевидний протиріччя з його визначенням фактора цілісності як «неекстенсивної» величини, що не має просторових характеристик]. Теорія Гурвича, якщо відкласти в сторону її подальшу деталізацію (1944), пов’язану з ідеєю клітинного поля, дозволяє наблизитися до розуміння простих і загальних законів, що лежать в основі онтогенетичного процесу. Приймається, що, починаючи з яйцеклітини, зародок утворює навколо себе анізотропне векторне поле, структура якого передвизначає результат розвитку на найближчому нескінченно малому його етапі. Після заповнення простору поля останнє «з’їдає себе» і реорганізується у поле з новими параметрами, обумовленими кінцевим станом зародка, досягнутим на попередньому етапі. Таким чином створюється установка розвитку на новий найближчий відрізок тощо. У ході процесу відбувається також формування полів окремих зачатків, підпорядкованих полю цілого. У цій концепції фактор цілісності (поле), безперервно спрямовуючи розвиток, сам у той же час є безперервною функцією шляху, пройденого субстратом його впливу. Ми маємо тут «закон Дріша в диференціальній формі» (Белоусов, 1979), тобто спрямоване ціле розглядається вже не як кінцева мета, а як властивість послідовних стадій, що перетворюються за законами причинності [Відчастково такий хід подій усвідомлював вже Дріш (1915), вказуючи, що виконання кожного етапу розвитку веде до «зміни чергового завдання ентелехії». По суті мова йде про зміну властивостей самого фактора цілісності]. Розвиток виступає як лавиноподібний процес з позитивним зворотним зв’язком між зачатком і його полем при цілеспрямованій поведінці зачатка щодо кожного нового встановленого стану поля. Хоча докази теорії стосуються скоріше часткових морфогенезів і стосуються в основному просторової сторони змін як найдоступнішої для вивчення, вони виглядають дуже вагомими. На багатьох прикладах показано (Гурвич, 1944), що детермінанція цілого зачатка здійснюється при невизначеному стані складових його елементів. Останні лише статистично детерміновані як сукупність («нормовані» за Гурвичем), підпорядковуючись полю цілого. Серед добре відомих прикладів — випадковий характер просторово‑часового розподілу окремих мітозів щодо осей симетрії в таких зачатках, як розвиваючийся корінь цибулі або сітківка ока; загальний результат цих випадкових подій залишається упорядкованим. Реальність поля наочно доводиться явищем «динамічної преформації» зачатка, коли орієнтація клітин його стінки визначається не фактичною конфігурацією останньої, а силовою поверхнею поза нею, що відповідає її проспективним контурам, набутим на найближчій наступній стадії.
Хоча побудова загальної теорії онтогенезу — справа майбутнього, вона, безсумнівно, не зможе обійтися без системних принципів, закладених у її основу Гурвичем. На користь цього говорять вже найпростіші міркування. 1. Розвиваючийся організм — це цілісна динамічна система, що виключає однозначний стан складових її процесів або лежачих в їх основі початкових елементів. 2. У кожен момент розвитку не існує іншої цілісності, ніж та, що властива зародку на цій стадії. Тому ідея перетворюваного цілого стає неминучою.
МНОЖИННІСТЬ ПОЧАТКОВИХ СТАНІВ ЯК ОСНОВА СТІЙКОСТІ НОРМИ
У стійкій системі збереження її параметрів базується на безперервній зміні її взаємодіючих частин. Таким чином регуляція цих параметрів при кожному збуренні і, відповідно, процес їх досягнення з початкового неравноважного стану, що становить телеономічну модель індивідуального розвитку, здійснюються однаково ефективно при множині варіантів початкових значень елементів системи. Цю незалежність кінцевих властивостей динамічної системи від її початкового стану називають еквіфінальністю в широкому розумінні терміну (Bertalanffy, 1969) [Еквіфінальність розуміла Дрішем (1915) як здатність особин одного виду до різних шляхів регенерації цілого організму при одному і тому ж експериментальному пошкодженні, тобто мова йшла про явище, близьке до того, що в порівняльній ембріології називається «окольним розвитком» (схожість початку і кінця типового розвитку у різних форм при різниці шляхів у проміжку). У значенні незалежності підсумку процесу від початкового стану це поняття було використано Л. Берталанфі (Bertalanffy, 1949, 1969), який вважає таку поведінку властивістю лише відкритих систем. Це здається правильним лише в тому сенсі, що для відкритої системи теоретично можливо збереження будь‑якого параметра, тоді як у закритій зовнішнє збурення веде до певних незворотних змін. Але на практиці для відкритих систем здатність до регуляції поширюється кожного разу не на всі їх властивості; наприклад, розвиток морських їжаків з ізольованих бластомерів або регенерація асцидій з оперованих особин веде до формування хоча й нормальних, але зменшених організмів (Дриш, 1915). З іншого боку, еквіфінальність закритих систем щодо максимуму ентропії завжди є абсолютною; проте для наших задач зручніше називати її принципом множинності початкових станів. У розвитку організмів ця закономірність має безліч очевидних проявів; особливо наочні випадки, коли для виду можливі різні способи розмноження (наприклад, статеве і вегетативне у асцидій, кишечнополостних тощо), при яких початкові стадії розвитку не мають між собою нічого спільного, хоча результат їх тотожний. Сюди ж входять усі експериментальні факти щодо онтогенетичної регуляції, зокрема випадки регенерації цілого організму з фрагментів спеціалізованих тканин (плоскі черв’яки, немертини тощо) або явище самоскладання зародка (наприклад, гаструли морських їжаків) з ізольованих клітин з подальшим відновленням нормального розвитку (Світлов, 1964, 1972). Взагалі, не лише реалізація цілого організму, а й виконання будь‑якого морфогенетичного акту в нормальному розвитку базується на відомій незалежності його результату від початкових умов. Наприклад, нормальна індукція здійснюється при значних коливаннях у масі і часі взаємодії членів індукційної системи, варіаціях у чутливості реактора, концентрації і складі активуючих речовин тощо (Шмальгаузен, 1964, 1982). Іншими словами, весь розвиток побудований на відносно стійких актах з «множинним забезпеченням», якщо розуміти під цим терміном будь‑які, а не лише якісні різниці у здійсненні одного і того ж морфогенезу. У загальній системі розвитку ці відносно стійкі події складають проміжний ієрархічний рівень (або рівні) між однозначно детермінованим цілим (взрослою нормою) і лежачими в основі системи статистично обумовленими елементарними процесами, наприклад, клітинними поділами в «регуляційних» онтогенезах. У цьому і полягає принцип нормування Гурвича: детермінація і формування окремого зачатка не пов’язані з жорсткою фіксацією початкового стану його елементів.
Принцип множинності початкових станів має далеко‑йдучі наслідки для теорії нормального розвитку, що стосуються вже тієї її області, яка історично виділилася в вчення про спадковість. Рухаючись у глибину розвитку аж до зиготи, ми повинні будемо зробити висновок, що стійкість реалізації дорослої норми (тобто ...){"translated_text":"це успадкованість) не може бути зведена до фіксованого набору станів якихось клітинних одиниць, у тому числі й одиниць хромосомного апарату (геному) зиготи або яйцеклітини. Цей набір має бути детермінований лише статистично (нормовано), тобто має зберігати невизначеність у межах, що допускають нормальне (еквіфінальне) завершення розвитку. Саме це і стверджується теорією стабілізуючого відбору. Згідно зі Шмальгаузеном (1982, с. 84, 174), стійкість організації не є властивістю елементів спадкової субстанції, а виразом взаємодії частин, що беруть участь у розвитку. Організм «стійкіший за свій генотип» (якщо розуміти останній як набір певних станів хромосомних одиниць). Це один із найважливіших висновків теорії, заснований на узагальненні емпіричних даних, є одночасно дедукцією з положення, що властивості системи не зводяться до властивостей її елементів. Адаптивна норма детермінується лише цілісною видоспецифічною структурою зародкової клітини, якій відповідає множина взаємозамінних варіантів геному, здатних реалізувати дану норму в типових умовах розвитку. Для цього набору варіантів можна ввести поняття «генотипічної норми».\\nЯкі факти підтверджує розглянутий висновок? Перш за все, встановлена С. С. Четвериковим (1926) генетична гетерогенність («насиченість мутаціями») природних амфімікотичних популяцій, прихована під покровом адаптивної норми і виявлена інбридингом. Безперервне перерозподілення елементів індивідуальних геномів у ході рекомбінації у поколіннях не змінює результат їх фенотипічної реалізації в основній масі. Однак принцип множинності початкових станів не містить заборон, які обмежували б варіабельність початкових факторів розвитку розв’язною здатністю гибрідологічного (менделевського) аналізу. Ми маємо підозрювати, що і в спільнотах, які на цьому рівні виглядають однорідними (у чистих лініях самопилювальних і клональних популяціях), насправді завжди існує приховане генетичне різноманіття. У цьому переконують численні експерименти з вирощування в екстремальних умовах жорстко відселених сортів автогамних рослин (злаків, бобових) і партеногенетично розмножуваних комах (тлі). При цьому завжди виявляється різноманіття індивідуальних фізіологічних реакцій, серед яких найжиттєздатніші варіанти піддаються закріпленню відбором, що супроводжується появою у них стійких морфологічних особливостей (Самохвалова, 1951, 1954; Шапошников, 1961, 1966; Агаєв, 1978). Хоча ці експерименти інтерпретуються по-різному, здається цілком очевидним, що мова йде про генетичну змінливість, що отримала фенотипічне вираження внаслідок виходу умов середовища за межі, що допускають нормальний (еквіфінальний) розвиток [Ці факти суперечать звичному погляду про неможливість відбору у чистих лініях, заснованому на класичних експериментах Йогансена (1933) з квасолею. Вчення про генетичну асиміляцію нестабільних ознак тепер дозволяє надати цим експериментам іншу інтерпретацію. Щоб лабільна реакція могла бути закріплена відбором, недостатньо лише прихованої гетерогенності її носіїв. Потрібно також: а) щоб останні здійснювали цю реакцію в одних і тих самих відхилених умовах і б) щоб відбір спочатку вівся в цих же самих умовах. Обидві ці вимоги в експериментах Йогансена не дотримувалися, і індивідуальні причини однакових модифікацій розмірів зерен не контролювалися. Приблизним аналогом такого експерименту була б спроба закріпити якийсь термоморфоз у дрозофілі за допомогою його проявів, отриманих при протилежних відхиленнях температури розвитку, і вести відбір серед безпосередніх нащадків, вирощених у нормальних умовах]. Результати цих експериментів принципово не відрізняються від результатів, отриманих при генетичній асиміляції (стабілізації) індивідуальних структурних і фізіологічних морфозів у ксеногамних організмів, таких як дрозофіла (Камшилов, 1941; Waddington, 1957). З цього можна зробити висновок, що при будь‑якому способі розмноження адаптивна норма дійсно реалізується на основі множинності допустимих станів геному зародкової клітини (або вегетативного зачатка).\\nЦя виродженість відповідності між варіаціями генетичної основи і результатом нормального розвитку становить, як уже сказано, один із найважливіших принципів теорії стабілізуючого відбору. За її уявленнями, стійкість норми виражається у створенні відбором регуляторного епігенетичного механізму, здатного в широких межах нивелювати (каналізувати) варіації генетичних факторів і умов середовища. Таким чином, формуючи адаптивну норму, відбір неминуче має підвищувати і допустиму генетичну варіабельність у її основі, як це і спостерігається на практиці. Підсумок нормального розвитку не зводиться до фіксованої суми початкових причин. Навпаки, для генетичної теорії еволюції, що намагається описувати фенотипи в термінах конкретних генів, логічно очікуваним результатом відбору є створення в популяціях максимальної однорідності за всіма відбираними алелями або їх комбінаціями. Невідповідність цього очікування реальній генетичній структурі норми змушує вдаватися до різних додаткових гіпотез (про баланс відбору і мутаційного процесу, розноправний або частотно‑залежний відбір тощо), сам їх надмірність (Kojima, 1971; Солбриг, Солбриг, 1982; Айала, 1981) свідчить про непереборність виникаючих труднощів.\\nЕПІГЕНЕТИЧНА СИСТЕМА ЯК ВИРАЗ ЗВ’ЯЗКУ ГЕНОТИПА І ФЕНОТИПА\\nАдаптивна норма не вичерпує можливостей розвитку індивідуальної зародкової клітини. Навколо нормального фенотипу лежить область різноманітних нестабільних відхилень (морфозів), що виникають при порушеннях умов розвитку. Сукупність таких реакцій, реалізованих на основі гетерогенної сукупності зигот (спроможних у звичайних умовах до нормального формоутворення), утворює «мобілізаційний резерв» популяції, тобто приховану змінливість, що, за теорією Шмальгаузена (1941, 1968б), є потенційним матеріалом еволюційних перетворень (Шишкін, 1981, 1984а, б). Але поряд з цими відхиленнями існує, на перший погляд, інше їх джерело — мутаційні варіації геному зародкової клітини, що порушують розвиток адаптивної норми навіть у звичних для неї умовах. Саме ця категорія аберацій розглядається більшістю еволюціоністів як матеріал природного відбору. Постає питання: чи дійсно мова йде про дві різні категорії відхилень і як вони співвідносяться з нормальним розвитком? У чому полягає спільність функціонування «нормальних» і «мутантних» геномів, що дозволяє розглядати їх як варіанти єдиного видового генотипу? Адже якщо одиничні мутаційні порушення не виводять розвиток за межі видоспецифічної області аномалій (Дубінін, 1966б, с. 240; Майр, 1968), то це тим більше стосується порушень, викликаних зовнішніми факторами. Підказує думка, що обмеження в обох випадках однакові. Насправді, паралелізм фенопроявлення мутаційних і модифікаційних змін, спостережуваний як у природі, так і експериментально, а також паралелізм між успадкованими (стійкими) і модифікаційними ознаками у близьких розах і видах становить добре відомий факт, закладений у основу ряду історично пов’язаних, хоча й глибоко різних еволюційних теорій (неоламаркістські концепції, ідея «органічного відбору» Моргана‑Болдуїна і теорія стабілізуючого відбору). Ця спільність дає нам право вважати, що вся притаманна виду сукупність можливих шляхів онтогенетичної реалізації є проявом стійких властивостей цілісної системи розвитку і що еволюційний процес веде до зміни структури всієї цієї системи.\\nЯкщо кожному виду дійсно властивий обмежений набір варіантів онтогенетичної реалізації, то очевидно, що він становить специфічний «простір можливостей», що характеризує поведінку даної системи. Для окремої зиготи цей простір дефінітивних станів може бути виражений на площині поля (розірваного на дискретні ділянки), а шляхи його реалізації — пучком розходящихся траєкторій, з яких у типічному випадку лише одна (з її кінцевими розгалуженнями) відповідає адаптивній нормі, розуміючійся як більш‑або‑менш вузько обмежений участок поля (рис. 1, а, б). Для всіх зародкових клітин виду зазначений простір є однаковим (еквіфінальним), і індивідуальні відмінності між ними (і, передусім, їх геноми) полягають лише у відносній ймовірності здійснення різних траєкторій розвитку за даних зовнішніх умов. Усі ті варіанти геному, для яких нормальний фенотип становить найймовірніший (стійкий) результат розвитку (рис. 1, б, n1–n3), можуть бути виділені як генотипічна норма; інші ж становлять те, що зазвичай називають мутаціями, хоча насправді всі геноми в популяції є мутантами один щодо одного. Будь‑яке порушення системи розвитку, що не руйнує її (тобто дозволяє завершити розвиток), може в такому випадку лише змінити вибір конкретної траєкторії, але не здатне дати результат, що виходить за межі видоспецифічного простору можливостей. Іншими словами, реакція системи на порушення завжди в кінцевому підсумку відтягуватиме специфіку самої системи незалежно від того, чи мало місце зовнішнє впливання на хід розвитку, чи порушення структури самої зародкової клітини.\\n \\n\\nРис. 1. Співвідношення між зиготами та типами розвитку\\na — еквіфінальність типового розвитку нормальних гетерогенних зигот; б — обмеженість видоспецифічного простору можливостей розвитку при будь‑яких варіаціях зародкового геному. Суцільні лінії — стабільний шлях, пунктирні — лабільні шляхи розвитку; p1–n3 — нормальні зиготи, m — аномальна, N — адаптивна норма"}Подібна теорія системи розвитку, що описує співвідношення між індивідуальною структурою зиготи, умовами розвитку та кінцевим результатом, ще далека від створення. І все ж головні її принципи зрозумілі вже давно. Можна стверджувати, що основу для неї заклало уявлення Р. Голдшмідта (Goldschmidt, 1938, 1940) про зводимість усіх фенотипічних відхилень незалежно від їх початкових причин до кількісних зсувів всередині системи розвитку. Іншим важливим елементом є вчення К. Воддінгтона (Waddington, 1957) про епігенетичний ландшафт як структуру цієї системи та про генетичну асиміляцію як спосіб її перебудови. Ми спробуємо пов’язати ці погляди в одну непротиворечливу концепцію і простежити найочевидніші висновки з них, що узгоджуються з експериментальними даними генетики. Поняття видоспецифічної системи розвитку використовується нами за вказаними авторами поряд із такими синонімами, як «реактивна система» або «епігенетична система». Подібне значення має і загальноприйняте поняття «норми реакції» (Johannsen, 1926), що означає можливості епігенетичної реалізації індивідуального генотипу.
\\n\\nРис. 1. Співвідношення між зиготами та типами розвитку\\na — еквіфінальність типового розвитку нормальних гетерогенних зигот; б — обмеженість видоспецифічного простору можливостей розвитку при будь‑яких варіаціях зародкового геному. Суцільні лінії — стабільний шлях, пунктирні — лабільні шляхи розвитку; p1–n3 — нормальні зиготи, m — аномальна, N — адаптивна норма"}Подібна теорія системи розвитку, що описує співвідношення між індивідуальною структурою зиготи, умовами розвитку та кінцевим результатом, ще далека від створення. І все ж головні її принципи зрозумілі вже давно. Можна стверджувати, що основу для неї заклало уявлення Р. Голдшмідта (Goldschmidt, 1938, 1940) про зводимість усіх фенотипічних відхилень незалежно від їх початкових причин до кількісних зсувів всередині системи розвитку. Іншим важливим елементом є вчення К. Воддінгтона (Waddington, 1957) про епігенетичний ландшафт як структуру цієї системи та про генетичну асиміляцію як спосіб її перебудови. Ми спробуємо пов’язати ці погляди в одну непротиворечливу концепцію і простежити найочевидніші висновки з них, що узгоджуються з експериментальними даними генетики. Поняття видоспецифічної системи розвитку використовується нами за вказаними авторами поряд із такими синонімами, як «реактивна система» або «епігенетична система». Подібне значення має і загальноприйняте поняття «норми реакції» (Johannsen, 1926), що означає можливості епігенетичної реалізації індивідуального генотипу. 
Рис. 2. Залежність алельних змін фенотипу від кількісних градацій фактора, що керує перемиканням шляхів розвитку (на прикладі крилавої мутації vestigial у дрозофіли). Підсилення теплового впливу на личинковий розвиток веде до зміни фенотипу мутантів у бік слабших алелей, аж до збігу з нормою: момент відхилення розвитку від нормального шляху зсувається на все пізніші стадії.
N — норма, vg1–vg5 — фенотипи алельної серії
Уявлення Голдшмідта про системні властивості індивідуального розвитку виходять з уже згаданого факту паралелізму мутаційних і модифікаційних змін (вперше експериментально показаного з генетичним контролем Н. В. Тимофеєвим‑Ресовським у 1926 р.). Як випливає з дослідів з дрозофілою та іншими організмами, ефект практично будь-якої мутації, включно з її плейотропними проявами, може бути отриманий у вигляді морфозу (фенокопії) за допомогою шокових впливів на той чи інший чутливий період розвитку. Відтак Голдшмідт зробив висновок, що обидва типи явищ мають одну і ту ж фізіологічну основу — неспецифічні порушення нормальної координації подій розвитку, а саме розбіжності швидкостей морфогенетичних реакцій та зміни кількостей, концентрацій і часу взаємодії реагуючих речовин. Можливості таких порушень, що допускають завершення розвитку, мають обмеження в межах системи, а отже обмежений і набір здійснених для неї фенотипічних відхилень. Мутації в даній системі здатні викликати лише ті аномалії, які можуть бути отримані і зовнішніми впливами, що призводять до аналогічних кількісних зсувів у ній. Мутаційні ефекти, включно з усіма їх плейотропними проявами, не є властивістю гена, а ембріологічним наслідком часу, місця і типу первинного порушення розвитку, викликаного мутацією (Голдшмідт, 1933; Goldschmidt, 1938, 1940, 1955; Камшилов, 1940; Шмальгаузен, 1982). Характер аномалій у підсумку відображає цілісні властивості самої системи, а не специфіку дії того чи іншого хромосомного локуса.
Усе це обґрунтовується численними даними феногенетичного аналізу мутацій, наприклад, таких, як Bar або vestigial у Drosophila melanogaster. Фенотипи всього ряду алелей (до норми) для кожної з них можуть бути отримані градацією температурних впливів на личинку. Очевидно, що специфічні алельні зміни фенотипу тут є реакцією системи розвитку на чисто кількісні зміни якогось фактора, чутливого до температури середовища. При цьому чим менше відхилення фактора від норми (воно нейтралізується у зазначених мутацій підвищенням температури), тим пізніше порушується розвиток (рис. 2). Так, при максимальному вираженні ефекту vestigial дефект проявляється вже на стадії уявного диска крила, і воно не розвивається далі основи; при мінімумі редукції крила формується повністю, і лише потім його пластина частково лізується у куколки (Goldschmidt, 1938, 1955). У ряді випадків природу кількісного фактора, що детермінує характер якісної фенотипічної аномалії, можна визначити дещо конкретніше. Наприклад, це може бути швидкість росту і сегментації зачатка, як показано для мутації aristopedia (у дрозофіли), пов’язаної з перетворенням перистого відділу антени (арист) у структуру, схожу на лапку кінцівки. У мутанта швидкість росту уявного диска підвищена до рівня, притаманного в нормі зачатку кінцівки; але при затримці росту за допомогою колхіцину розвиток залишається в межах нормального шляху (Балкашіна, 1928; Goldschmidt, 1938, 1955). У мишей дія мутації Dh, що викликає полідактію задніх кінцівок, базується на уповільненні загибелі клітин апікального ектодермального гребеня, внаслідок чого продовжується його індуктивна дія на нирку кінцівки. Навпаки, мутація Os, що прискорює загибель гребеня і скорочує термін індукції, веде до олігодактілії або навіть редукції самої нирки (Конюхов, Нончев, 1981). Приклади подібного роду змушують багатьох дослідників визнавати неспецифічний і непрямий вплив генів на вибір реалізованого фенотипу (Wolpert, 1976; Alberch, 1982).
Якщо характер аномалії дійсно не визначається прямо специфікою початкового порушення, то очевидно, що це має виявлятися і при різних способах зовнішнього впливу на розвиток. Експерименти з фенокопіями підтверджують це. Багато типів шоків, спрямованих на один і той же чутливий період розвитку, дають однаковий результат, і, навпаки, один і той же шок може викликати якісно різні аномалії залежно від зачепленої фази розвитку, а також його сили і тривалості (Goldshmidt, 1955). З іншого боку, ми повинні очікувати, що і різні за своєю природою генетичні порушення приведуть до тотожних аномалій. Це дійсно має місце. Увесь досвід експериментальної генетики говорить про те, що унікальних за своїм ефектом мутацій, схоже, не існує (Тимофеєв‑Ресовський, Іванов, 1966), і це означає лише інше формулювання тези Голдшмідта про неспецифічність впливу мутацій на систему розвитку. Відповідно дослідниками виділяються більш чи менш обширні «гетерогенні групи генів», що мають різну локалізацію, але викликають схожі або ідентичні аномалії при мутації. Характерний приклад — група minute у дрозофіли (скорочення торакальних щетинок), що охоплює близько 60 локусів у трьох хромосомах (Тимофеєв‑Ресовський, Іванов, 1966). Точно так само фенотип bithorax (подвоєння середньогруди) може бути отриманий мутацією одноіменного локуса в III хромосомі, комбінацією мутацій у трьох різних хромосомах і, нарешті, на основі «нормального» геному — за допомогою шокового впливу на личинку (Waddington, 1966). Те саме виявляється і при аналізі природних фенотипічних відхилень, де одні і ті ж їх типи виявляються пов’язаними з мутаціями в різних хромосомах або просто з зовнішніми впливами (наприклад, фенотип Abnormal abdomen у дрозофіли; Голубовський та ін., 1974). Нарешті, добре відомо, що однакові нормальні або аберантні ознаки у близьких видів чи географічних рас одного виду дуже часто реалізуються на різній генетичній основі; до таких звичних прикладів належать паралелі у забарвленні листків у видів бавовники (Harland, 1937) або крил у метеликів, зокрема жовте забарвлення у німецькій і італійській формах Callimorpha dominula або біле — у англійській і канадській формах Biston betularia (Goldschmidt, 1940; Sheppard, 1970). При генетичному аналізі ці аналоги дають різні типи розщеплення. У випадку різних видів, де такий аналіз зазвичай ускладнений, подібні приклади прийнято пов’язувати з паралельними, або гомологічними, мутаціями; але насправді ми маємо мати на увазі не наявність «того ж гена», а схожість епігенетичних систем, що допускають здійснення одного і того ж шляху розвитку при різній генетичній структурі (Goldschmidt, 1945).
Усі ці факти ясно вказують на невизначений у цілому характер відповідності між первинними елементарними порушеннями процесу розвитку і його тим чи іншим конкретним підсумком. Сукупність таких підсумків зберігає в межах виду стійкість щодо варіацій умов розвитку, тобто жодна фенотипічна ознака не детермінується фіксованою комбінацією станів хромосомних локусів і зовнішніх факторів. Цей контраст (асиметрія) між множинністю можливих умов розвитку і обмеженістю спектра його можливих результатів показує, що тут порівнюються події, що належать до різних ієрархічних рівнів однієї цілісної динамічної системи, що володіє стійкою поведінкою. По суті феногенетичний аналіз мутантних аномалій виявляє те саме системне властивість розвитку, з якою раніше зіткнулася експериментальна ембріологія, — принципову неможливість зведення підсумку розвитку до певної суми початкових причин. Але додатково стає очевидним, що це стосується не лише нормального ходу онтогенетичного процесу, а й усього простору його аберацій.
Все це, звичайно, не означає заперечення специфіки первинної функції геномних локусів. Йдеться лише про те, що її зміни самі по собі не детермінують ознаки фенотипу, а виражаються в кількісних порушеннях певних параметрів системи розвитку, на які вона реагує якісно, змінюючи вибір реалізованого в ній фенотипічного результату. При цьому не мається на увазі ідентичність первинних продуктів різних елементарних порушень, що викликають однакові аберації. Механізм їх дії має бути різним. При геномних змінах — це ті чи інші ланцюги реакцій, що ведуть до нерегульованих зсувів у певній фазі розвитку; зовнішні ж стимули порушують цю фазу більш прямим шляхом (Goldschmidt, 1955).
Викладені уявлення передбачають, що параметри морфогенетичних процесів, що складають розвиток, характеризуються в кожній епігенетичній системі певним рядом порогових значень, від досягнення яких відбувається вибір тієї чи іншої траєкторії розвитку. Необхідно уточнити, як цей вибір здійснюється, і спробувати зрозуміти, у чому полягає різниця між нормальними і аберантними шляхами розвитку в межах однієї системи. Відповідь на ці питання значно прояснює запропонована Воддінгтоном (1947, 1960, 1970а; Waddington, 1957, 1966) модель епігенетичного ландшафту, що описує загальні властивості системи розвитку у вигляді серії розгалужених нахилених долин, що дивергують з загальної початкової точки (рис. 3, 7). Ця модель має подвійний сенс. По‑перше, вона відображає звичну інтерпретацію розвитку як ієрархії етапів все більш специфічної диференціації зачатків, що веде до послідовного обмеження їх подальших формативних можливостей. При цьому система траєкторій, або долин, якими «рухаються» окремі процеси, символізує обмеженість і певну дискретність шляхів диференціації, можливих для дериватів даного зародка або зачатка. Це властивість розвитку становить добре відомий факт, що демонструється, наприклад, поведінкою ембріональних тканин у експлантатах (Светлов, 1964). Інший, більш суттєвий сенс моделі Воддінгтона полягає в тому, що вона дозволяє зобразити шлях нормального розвитку окремого зачатка (і в граничному випадку — всього організму) на фоні всього поля потенційних можливостей його розвитку в межах епігенетичної системи, властивої даному виду. У цьому випадку нормальному шляху відповідає глибока долина, або креод (буквально — «необхідний шлях»), а альтернативним можливостям — її більш пологі відгалуження. Відхилення на будь‑яке з них пов’язане з подоланням більш‑менш високого порогу (що відокремлює цю долину від дна креода) і означає порушення нормального ходу розвитку. Але оскільки точки розгалужень відповідають зниженню стінок креода (рис. 3), вони позначають моменти відносної нестабільності у виборі детермінації, тобто чутливі періоди, вплив на які пов’язаний з отриманням експериментальних аберацій. 
Рис. 3. Співвідношення між будовою епігенетичного ландшафту та характером пошкоджуючого впливу
а — відхилення розвитку в бічну долину за рахунок сильного зовнішнього впливу (довга стрілка); в — таке ж відхилення під дією сильної мутації, що порушує креод; б — проміжний стан (за Воддінгтоном, 1957, інтерпретація змінена)
Стійке (каналізоване) розвиток, або рух по креоду, що веде до адаптивної норми, забезпечується його регуляцією. Остання виражається в тому, що процес, будучи відхилений тим чи іншим впливом, знову «скочується» в русло долини, якщо зсув не вийшов за межі її схилів. Оскільки з ходом нормального розвитку здатність його до регуляції зазвичай падає, це означає поступове випрямлення креода. При наявності кількох адаптивних норм система розвитку має кілька альтернативних креодів, вибір серед яких контролюється або факторами середовища (при модифікаційному поліморфізмі), або закономірним рекомбінацією хромосомного апарату (наприклад, детермінація статі). На відміну від норми, аберантні шляхи розвитку, представлені випрямленими долинами, мають обмежені можливості регуляції, і їх підсумок відносно лабільний (що є загальним правилом для мутацій і неадаптивних морфозів; Шмальгаузен, 1968б, Воддінгтон, 1944, 1970а). Іншими словами, дискретність цих шляхів щодо норми не означає їх стійкості. У свою чергу сама ця дискретність не абсолютна і проявляється тим слабше, чим пізніше в розвитку настає незворотне відхилення. Наприклад, вона мінімальна або відсутня у слабких виразах таких мутацій у дрозофіли, як eyeless або vestigial (Goldschmidt, 1938; Rapaport, 1943).
Отже, епігенетичний ландшафт характеризує видоспецифічний простір можливостей розвитку, що охоплює області стійкого течії процесу (креоди), області найймовірніших аберацій (бічні долини) і зони з мінімальною ймовірністю здійснення (водоразділи між долинами, які траєкторія розвитку завжди прагне залишити). Ця структура відображає властивості цілісної динамічної системи, показуючи, що її реакція на збурення залежить від того, яка точка її просторово‑часового протягнення була піддана впливу. Чим ближче вона до області креода, тим імовірніше, що різноманітні за своєю природою впливи будуть однаково «забуферені», і навпаки, у зонах нестабільності схожі причини можуть мати глибоко різні наслідки. У більш загальній формі хід усього розвитку як багатокомпонентної системи може бути описаний рухом точки у багатовимірному (фазовому) просторі, де її координати в кожен момент часу відповідають мірам окремих взаємодіючих елементарних компонентів (Waddington, 1957). Креоди відповідають найстабільнішим траєкторіям, здатним притягувати до себе близькі точки; вся ж сукупність можливих траєкторій у цьому просторі становить «фазовий портрет» системи (Белоусов, 1979; Белоусов, Чернавський, 1977), наочним відображенням якого є тривимірна модель епігенетичного ландшафту. Демонстрована нею відносно проста упорядкованість системи розвитку є «властивістю вищого порядку» щодо функцій елементарних геномних одиниць (Waddington, 1957, с. 34) і базується на взаємодії всієї їх сукупності.
Розглянемо тепер характер реакцій епігенетичної системи на елементарні збурення. Очевидно, що такі впливи не можуть перетворювати саму систему, а лише змінюють її стан. Вони або змінюють вибір траєкторії в межах її ландшафту, або піддаються регуляції і взагалі не змінюють результату розвитку. У обох випадках кінцевий результат визначається властивостями самої системи.
У випадку впливів, інтенсивність яких перевищує можливості регуляції каналізованого розвитку, їх ефект, очевидно, може мати два кількісних інтерпретації. Або рівень якогось критичного фактора, здатного відхилити розвиток у бічну долину, перевищує поріг, допустимий у даній часовій точці для нормального ходу процесу, або зникає сам цей поріг, що відокремлює креод від даної долини (тобто підвищується чутливість до перемикаючого фактора). Нарешті можливі обидва типи змін одночасно. Очевидно, у цьому і полягає сутність тих кількісних зсувів, до яких, як показав Голдшмідт, неминуче мають зводитися наслідки всього різноманіття елементарних впливів на систему розвитку.
Виходячи з сказаного, можна описати можливі результати впливу окремого пошкоджуючого фактора на індивідуальні цикли розвитку в нормальній гетерогенній популяції. Усі зиготи одного виду належать варіантам однієї й тієї ж епігенетичної системи; їх генетичні різниці за однакових умов розвитку зумовлюють індивідуальні деталі моделювання ландшафту (різну висоту порогів між окремими долинами, різницю у виразності останніх). Для нормальних зигот ці різниці мінімальні, тобто шлях, що веде до нормального фенотипу, є для них найстабільнішим (каналізованим) і виявляє в цьому відношенні лише часткові варіації, виражені як локальні різниці у висоті захисних порогів креода.
Для елементарних впливів, здатних викликати тут відхилення розвитку від норми, можливі три основні ситуації (Шишкін, 1984а, б). 1. Різке зміна геному, що призводить до настільки сильного зниження захисного порогу на певному ділянці креода, що незалежно від варіацій його попередньої висоти розвиток незмінно відхиляється на один і той же бічний шлях (рис. 3, в). Це — ідеальна мутація, найзручніша для генетичного аналізу, тобто таке локусне зміна, яке при введенні в будь‑який варіант геному викликає з максимальною ймовірністю певну аномалію розвитку.{
"translated_text": "Проте на практиці результат все ж виявляється не зовсім стабільним, оскільки згладженість аберативних долин ландшафту сама по собі виключає ефективну регуляцію ними онтогенетичних траєкторій. І дійсно, на практиці вираження навіть сильних «сирих» мутацій залишається змінним (Waddington, 1970a). 2. Протилежна ситуація — предельне сильне зовнішнє впливання на хід розвитку, що перевищує будь‑який поріг його стабільності у даній часовій точці і призводить у цьому циклі розвитку до того ж результату, що й сильна мутація (рис. 3, а). 3. Між цими двома крайніми випадками лежить величезна область проміжних станів, коли характер і сама можливість ухилення залежать від конкретного співвідношення між особливостями епігенетичного ландшафту та умовами розвитку (рис. 3, б). Дія однієї і тієї ж мутації буде або лежати нижче порогу порушення, або перевищувати цей поріг у різних точках креода — залежно від індивідуальних особливостей ландшафту (визначених вихідною конституцією геному) і коливань факторів середовища. І навпаки, одні й ті ж ухилення мають виникати при різних комбінаціях зовнішніх і внутрішніх умов розвитку. У цих випадках говорять про мутації з нестабільним вираженням і проявом, тобто не показуючі при аналізі правильного менделевського успадкування.\nЦя остання, найтиповіша ситуація відповідає реальній картині невизначеної змінливості, спостережуваної в природних популяціях. Загальновідомий факт відсутності або рідкості в них тієї категорії аберацій, яка може бути охарактеризована як домінантні мутації з хорошим проявом (Gershenzon, 1941). При цьому навіть великі однотипні аберації при аналізі виявляються пов’язаними з різними хромосомами або частково індукованими зовні (наприклад, Abnormal abdomen у дрозофіли; Holubovskyi et al., 1974). При наявності достатньо великих вибірок таких фенотипів вони демонструють найрізноманітніші градації за стабільністю успадкування — від співвідношень, близьких до менделевських, до повної втрати прояву (наприклад, фенотип «плямисті очі» у дрозофіли; Dubinin et al., 1937). Тому всередині таких груп ізореагентів автори часто взагалі не наважуються провести межу між успадкованими (мутаційними) і неуспадкованими (модифікаційними) змінами (Balkashina, Romashov, 1935) або ж шукають її між лініями з мінімальним успадкуванням аберації і лініями з повним її відсутністю. Цілком очевидно, що мова йде про межу, якої немає в природі. Усі фенотипи одного класу являються варіантами реалізації однієї і тієї ж онтогенетичної траєкторії, що різняться за ступенем стабільності в силу того, що вони обумовлені найрізноманітнішими поєднаннями індивідуальної генетичної конституції та факторів середовища. Поняття «мутації» і «модифікації» на практиці абсолютно несумісні, оскільки перше стосується порівняння особин, а друге — порівняння можливостей розвитку однієї і тієї ж особи. Усі однакові фенотипи (як і будь‑які інші) завжди генетично не ідентичні і тому можуть розглядатися як приховані мутанти щодо один одного безвідносно до результатів гібридного аналізу. З іншого боку, будь‑який фенотип, оцінений на підставі такого аналізу як мутантний, являє собою лише одну з можливостей розвитку в межах епігенетичної системи даної зиготи, тобто одну з альтернатив (модифікацій) щодо нормальної для виду траєкторії. Останнє особливо очевидне для випадків, коли повернення на цю траєкторію практично легко здійснити шляхом зміни умов розвитку (наприклад, мутації pennant, vestigial, Abnormal abdomen тощо у дрозофіли; Shmalhausen, 1968).\nОтже, гетерогенність однотипних аномалій розвитку, передбачувана моделлю епігенетичного ландшафту, підтверджується реальною картиною їх успадкування в природних популяціях. Точно так само підтверджується і інший висновок з цієї моделі — що одні й ті ж зовнішні збурення, як правило, викликатимуть розпрямлені відхилення від нормального розвитку відповідно до індивідуальної конституції уражених ними зигот. Посилення змінливості при порушенні нормальних умов, відмічений ще Дарвіном, становить добре відомий факт, порівнюваний з розкладом променя світла при проходженні через призму (Lobashev, 1947). Перехід популяції під дією екстремальних умов від фенотипічної однорідності до прояву розпрямлених варіацій був позначений як виявлення «мобілізаційного резерву» змінливості (Shmalhausen, 1941; Gershenzon, 1941). Це явище означає, що каналізуючі механізми розвитку, які буферизують індивідуальні генетичні відмінності особин, виявляються порушеними при досягненні певного порогу зовнішніх впливів, в результаті чого ці відмінності проявляються у вигляді фенотипічних аберацій. Усі нормальні зиготи в популяції різняться як за шириною інтервалу умов, що допускають каналізоване розвиток, так і за характером морфозів, здійснюваних ними в однакових надпорогових умовах (рис. 4, 5).\nНа підставі уявлення про епігенетичний ландшафт можливі й інші передбачення, доступні експериментальній перевірці. Якщо порушення нормального розвитку органу А, обумовлене мутацією (або будь‑яким пошкоджуючим впливом), зводиться до зміни міри якогось фактора, дестабілізуючого траєкторію А, то можна очікувати, що при штучному переключенні розвитку даного зачатка в напрямку органу Б його нова траєкторія буде незалежна від зазначеного фактора, тобто виявиться «поза досяжністю» мутацій, що впливають лише на орган А. І навпаки, мутації, що зачіпають розвиток органу Б, мають діяти і на будь‑який інший зачаток, детермінований у напрямку Б, тобто чутливість зачатків до тих чи інших порушень має визначатися не стільки їхньою нормальною детермінацією, скільки вибором фактичного шляху їх розвитку. Ці закономірності дійсно були встановлені I. A. Rapaport (1941), який показав, що у дрозофіли ефект домінантної мутації Met, що веде до поглинання презумптивного матеріалу крила мезотораксом, не піддається жодним змінам при введенні в геном різних крилих мутацій. З іншого боку, мутації, які в нормі впливають на будову спинних щетинок, але не зачіпають крила, здійснюють таку ж дію і на гігантський мезоторакс, змінений мутацією Met.\n \nРис. 4. Індивідуальні внутрі‑популяційні різниці у стабільності нормального формоутворення\nа — криві залежності фенотипічної реалізації окремих геномів від зовнішніх умов суцільні ділянки відповідають нормі (з модифікаціями А і Б), пунктирні — неадаптивним морфозам; б — модифікаційні спектри, контрольовані цими геномами в одному і тому ж інтервалі умов, що виходить за межі нормальних. Ділянки спектрів, що відповідають модифікаціям А і Б, заштриховані\n
\nРис. 4. Індивідуальні внутрі‑популяційні різниці у стабільності нормального формоутворення\nа — криві залежності фенотипічної реалізації окремих геномів від зовнішніх умов суцільні ділянки відповідають нормі (з модифікаціями А і Б), пунктирні — неадаптивним морфозам; б — модифікаційні спектри, контрольовані цими геномами в одному і тому ж інтервалі умов, що виходить за межі нормальних. Ділянки спектрів, що відповідають модифікаціям А і Б, заштриховані\n \nРис. 5. Залежність формоутворення від умов розвитку\nСуцільна лінія відповідає загальній для популяції адаптивній нормі (з двома модифікаціями); пунктирні — індивідуальним морфозам. По горизонталі — зміна середовища, по вертикалі — результат розвитку: а–а1 — інтервал умов, що допускає нормальний розвиток\nЗ розглянутих уявлень випливає ще один висновок — що дискретність змін фенотипу, викликаних мутаціями одного хромосомного локуса, виражає властивості цілісної системи розвитку, а не безпосередньо самих цих мутацій. Ще ранніми роботами Голдшмідта (1916–1917) щодо механізму визначення статі та S. Wright (Wright, 1916) щодо феногенезу мутацій альбінізму було показано, що в основі алельних змін лежать градації одного і того ж фактора (наприклад, кількості речовини або швидкості реакції), що діють з пороговим ефектом. З точки зору моделі епігенетичного ландшафту ця дискретність реагування (зменшувана у пізніх стадіях розвитку) є неминучим наслідком того, що стабільність каналізованої траєкторії до коливань рівня тих чи інших морфогенетичних факторів знижується у часі періодично, утворюючи перепади в точках «чутливих періодів», що відповідають гілкам аберативних долин (рис. 6, а). Чим більше рівень даного фактора виходить під дією мутацій за регульовані межі, тим ранішою з цих критичних фаз виявляється ураженою, тобто реалізується траєкторія, все більш глибоко дивергентна щодо нормальної. Коливання фактора між двома пороговими рівнями (рис. 6, б, рівні В і С), що визначають предельні можливості здійснення даного ухилення, не отримують фенотипічного вираження, і всі викликаючі їх однолокусні мутації будуть розцінюватися як один і той же алель. Перевищення фактора за один з порогів призведе до дискретної зміни фенотипу, тобто переключення розвитку на більш раннє або більш пізнє гілкування нормальної траєкторії.\nЛедве чи потрібно пояснювати, що концепція епігенетичної системи об’єктивно лежить в основі теорії стабілізуючого відбору. Уявлення про розвиток як систему з обмеженим набором найймовірніших траєкторій (фенотипічних станів) дозволило зруйнувати непрохідну межу між мутаційними та екзогенними змінами, що існувала для класичної генетики (Dobzhansky, 1947, s. 209), зводячи і ті, і інші до дискретних реакцій цілісної системи на кількісні зміни перемикаючих епігенетичних факторів (Shmalhausen, 1982, s. 82, 89, 103, 170–173). Встановлення цієї закономірності відкрила шлях до розуміння того, що стабільність успадкування являє собою не властивість окремих хромосомних генів, а результат створення відбором нової організації генотипу, що забезпечує каналізоване розвиток колишньо лабільного (неуспадкованого) признаку. Спадковість таким чином перетворилася на цілісну і історично обумовлену властивість, визначувану як стабільність результату епігенетичних взаємодій (Shmalhausen, 1982; Waddington, 1957), а природний відбір із сортувальника незалежних від нього успадкованих одиниць перетворився в механізм створення успадкованих змін.\n
\nРис. 5. Залежність формоутворення від умов розвитку\nСуцільна лінія відповідає загальній для популяції адаптивній нормі (з двома модифікаціями); пунктирні — індивідуальним морфозам. По горизонталі — зміна середовища, по вертикалі — результат розвитку: а–а1 — інтервал умов, що допускає нормальний розвиток\nЗ розглянутих уявлень випливає ще один висновок — що дискретність змін фенотипу, викликаних мутаціями одного хромосомного локуса, виражає властивості цілісної системи розвитку, а не безпосередньо самих цих мутацій. Ще ранніми роботами Голдшмідта (1916–1917) щодо механізму визначення статі та S. Wright (Wright, 1916) щодо феногенезу мутацій альбінізму було показано, що в основі алельних змін лежать градації одного і того ж фактора (наприклад, кількості речовини або швидкості реакції), що діють з пороговим ефектом. З точки зору моделі епігенетичного ландшафту ця дискретність реагування (зменшувана у пізніх стадіях розвитку) є неминучим наслідком того, що стабільність каналізованої траєкторії до коливань рівня тих чи інших морфогенетичних факторів знижується у часі періодично, утворюючи перепади в точках «чутливих періодів», що відповідають гілкам аберативних долин (рис. 6, а). Чим більше рівень даного фактора виходить під дією мутацій за регульовані межі, тим ранішою з цих критичних фаз виявляється ураженою, тобто реалізується траєкторія, все більш глибоко дивергентна щодо нормальної. Коливання фактора між двома пороговими рівнями (рис. 6, б, рівні В і С), що визначають предельні можливості здійснення даного ухилення, не отримують фенотипічного вираження, і всі викликаючі їх однолокусні мутації будуть розцінюватися як один і той же алель. Перевищення фактора за один з порогів призведе до дискретної зміни фенотипу, тобто переключення розвитку на більш раннє або більш пізнє гілкування нормальної траєкторії.\nЛедве чи потрібно пояснювати, що концепція епігенетичної системи об’єктивно лежить в основі теорії стабілізуючого відбору. Уявлення про розвиток як систему з обмеженим набором найймовірніших траєкторій (фенотипічних станів) дозволило зруйнувати непрохідну межу між мутаційними та екзогенними змінами, що існувала для класичної генетики (Dobzhansky, 1947, s. 209), зводячи і ті, і інші до дискретних реакцій цілісної системи на кількісні зміни перемикаючих епігенетичних факторів (Shmalhausen, 1982, s. 82, 89, 103, 170–173). Встановлення цієї закономірності відкрила шлях до розуміння того, що стабільність успадкування являє собою не властивість окремих хромосомних генів, а результат створення відбором нової організації генотипу, що забезпечує каналізоване розвиток колишньо лабільного (неуспадкованого) признаку. Спадковість таким чином перетворилася на цілісну і історично обумовлену властивість, визначувану як стабільність результату епігенетичних взаємодій (Shmalhausen, 1982; Waddington, 1957), а природний відбір із сортувальника незалежних від нього успадкованих одиниць перетворився в механізм створення успадкованих змін.\n \nРис. 6. Дискретність аберацій як вираження поступового зниження стійкості до перешкод каналізованого розвитку\nа — ділянка епігенетичного ландшафту, що показує перепади висоти стінок (пороги стійкості) креода у місцях гілкування аберативних долин; б — залежність характеру аберації від співвідношення між рівнем пошкоджувального морфогенетичного фактора та стійкістю креода. У проміжку між двома порогами стійкості (заштриховано) коливання міри фактора не змінюють тип розвитку; a, b, c, d — порогові рівні стійкості послідовних ділянок креода, A1–A3 — аберативні шляхи розвитку, N — нормальний хід змін рівня фактора і типічний шлях розвитку. По вертикалі — рівень фактора, по горизонталі — час\nУявлення Шмальгаузена (1968a, 1982) про історичне становлення нових адаптивних ознак легко інтерпретуються як опис перетворень видоспецифічного епігенетичного ландшафту. Весь процес, згідно з цими поглядами, починається щоразу з лабілізації розвитку попередньої норми (рис. 7, а, б; N) і відбору однієї з виникаючих при цьому елементарних неадаптивних реакцій. Це означає, що в нових умовах індивідуальні варіанти ландшафту реалізують різні аберативні траєкторії, з яких одна веде до найбільш життєздатного фенотипічного ухилення (рис. 7, б, N1). У міру відбору на її користь попередня нормальна траєкторія все більше втрачає стійкість безвідносно до умов розвитку, тобто її долина згладжується, що означає стирання різниць у стабільності між попередньою нормою та відбираним ухиленням (рис. 7, б, N, N1). Поетапна селективна стабілізація нової адаптивної траєкторії перетворює її на креод (рис. 7, в, г, N1); разом з цим вся зона ландшафту навколо неї переходить з області малоймовірних подій у область найпоширеніших ухилень, тобто вона розщеплюється новими долинами. Таким чином, поступово змінюється весь малюнок ландшафту (рис. 7, а–г). Іншими словами, відбувається зміна аберативного простору епігенетичної системи.\n
\nРис. 6. Дискретність аберацій як вираження поступового зниження стійкості до перешкод каналізованого розвитку\nа — ділянка епігенетичного ландшафту, що показує перепади висоти стінок (пороги стійкості) креода у місцях гілкування аберативних долин; б — залежність характеру аберації від співвідношення між рівнем пошкоджувального морфогенетичного фактора та стійкістю креода. У проміжку між двома порогами стійкості (заштриховано) коливання міри фактора не змінюють тип розвитку; a, b, c, d — порогові рівні стійкості послідовних ділянок креода, A1–A3 — аберативні шляхи розвитку, N — нормальний хід змін рівня фактора і типічний шлях розвитку. По вертикалі — рівень фактора, по горизонталі — час\nУявлення Шмальгаузена (1968a, 1982) про історичне становлення нових адаптивних ознак легко інтерпретуються як опис перетворень видоспецифічного епігенетичного ландшафту. Весь процес, згідно з цими поглядами, починається щоразу з лабілізації розвитку попередньої норми (рис. 7, а, б; N) і відбору однієї з виникаючих при цьому елементарних неадаптивних реакцій. Це означає, що в нових умовах індивідуальні варіанти ландшафту реалізують різні аберативні траєкторії, з яких одна веде до найбільш життєздатного фенотипічного ухилення (рис. 7, б, N1). У міру відбору на її користь попередня нормальна траєкторія все більше втрачає стійкість безвідносно до умов розвитку, тобто її долина згладжується, що означає стирання різниць у стабільності між попередньою нормою та відбираним ухиленням (рис. 7, б, N, N1). Поетапна селективна стабілізація нової адаптивної траєкторії перетворює її на креод (рис. 7, в, г, N1); разом з цим вся зона ландшафту навколо неї переходить з області малоймовірних подій у область найпоширеніших ухилень, тобто вона розщеплюється новими долинами. Таким чином, поступово змінюється весь малюнок ландшафту (рис. 7, а–г). Іншими словами, відбувається зміна аберативного простору епігенетичної системи.\n \n\nРис. 7. Перебудова системи розвитку в процесі становлення нової елементарної адаптації\n1 — зміни епігенетичного ландшафту, 2 — типові зміни модифікаційних спектрів, 3 — зміни дисперсії перетворюваного адаптивного ознака в популяції: а — каналізоване розвиток фенотипу N (що відповідає головній смузі в спектрах і піку варіаційної кривої); б — дестабілізація розвитку, що веде до зниження частоти фенотипу N і зменшення його ролі в спектрах; в, г — дві послідовні стадії каналізації онтогенетичної траєкторії N1 з розвитком навколо неї нової мережі аберативних долин. Фенотип N1 з нестабільного морфозу перетворюється на нову адаптивну норму; попередня норма стає морфозом або зникає. Жирними лініями виділені креоди; заштриховані типи розвитку, що відповідають послідовним нормам. По вертикалі — частота зустрічності ознаки, по горизонталі — зміна ознаки."
\n\nРис. 7. Перебудова системи розвитку в процесі становлення нової елементарної адаптації\n1 — зміни епігенетичного ландшафту, 2 — типові зміни модифікаційних спектрів, 3 — зміни дисперсії перетворюваного адаптивного ознака в популяції: а — каналізоване розвиток фенотипу N (що відповідає головній смузі в спектрах і піку варіаційної кривої); б — дестабілізація розвитку, що веде до зниження частоти фенотипу N і зменшення його ролі в спектрах; в, г — дві послідовні стадії каналізації онтогенетичної траєкторії N1 з розвитком навколо неї нової мережі аберативних долин. Фенотип N1 з нестабільного морфозу перетворюється на нову адаптивну норму; попередня норма стає морфозом або зникає. Жирними лініями виділені креоди; заштриховані типи розвитку, що відповідають послідовним нормам. По вертикалі — частота зустрічності ознаки, по горизонталі — зміна ознаки."
}Отже, на будь‑якому етапі своїх еволюційних змін система виявляє специфічний набір можливостей розвитку, притаманний їй лише в даний період її історії. Тому твердження, що матеріалом еволюції є випадкові геноваріації, є, за висловом Воддінгтона, «порожнім» (Waddington, 1957, с. 188). Якими б випадковими не були нуклеотидні зміни хромосомної ДНК, їх можливий вплив на фенотип завжди обмежений історично сформованою структурою (ландшафтом) епігенетичної системи. Зміна цієї структури виражається на популяційному рівні як зміна характеру невизначеної змінливості (Камшилов, 1967). Тому вказівка Дарвіна (1952) про те, що змінливість не викликається відбором, має бути правильно зрозуміла. Незалежним від відбору є лише саме її існування, а не характер її фенотипічного вираження.
**ЕПІГЕНЕТИЧНА СИСТЕМА ТА РОЗУМІННЯ МЕНДЕЛЬСЬКОГО ФАКТОРА**
Наведені уявлення неминуче підводять нас до проблеми онтогенетичної інтерпретації мендельського успадкування. Поняття мендельського фактора означає наявність двох стійких альтернативних (аллельних) станів ознаки, кожен з яких успадковується в відповідній «чистій лінії» однозначно, а у гибридів (другого покоління) — у певних числових співвідношеннях. З епігенетичної точки зору це означає, що розвиток ознаки у порівнюваних груп особин каналізовано в двох різних напрямках, причому стійкість кожного з них базується на взаємодії всіх елементів генотипу і виражається в нечутливості до ефекту рекомбінацій, що виникають при схрещуванні всередині даної групи (лінії). При міжлінійному схрещуванні обидва напрямки (креоди) об’єднуються в одному епігенетичному ландшафті як стабілізовані розгалуження однієї траєкторії, і вихід розвитку в принципі може коливатися між двома крайніми ситуаціями. При одній з них реалізується та чи інша проміжна аберантна траєкторія, що часто дає не зовсім стійкий результат, який у практиці мендельського аналізу описується як відсутність однорідності гибридів першого покоління (Філіпченко, 1924). У другому випадку (рис. 8) розвиток спрямовується до однієї з двох наявних областей притягання (креодів), тобто має місце виражене домінування. При будь‑якому варіанті вибір шляху розвитку залежить від неспецифічного зсуву у фазі нестабільної детермінанції (точці розгалуження креодів), пов’язаного в кінцевому підсумку з функцією тієї чи іншої пари гомологічних геномних одиниць. Остання ототожнюється моргановською хромосомною генетикою з алельними станами мендельського гена. Як уже зазначено, феногенетичний аналіз алельних серій фенотипів показує, що їх дискретність відображає не властивості самого детермінуючого фактора, а порогову реакцію системи розвитку на його кількісні зміни. З цієї точки зору дія пари локусів, відповідальних при даному стані системи за перемикання шляху розвитку, може контролювати в ній три градації фактора — два крайні (батьківські) — АА і аа і одну проміжну (гибридну) — Аа, позначувані генетикою відповідно як гомо‑ і гетерозиготні стани. Між ними лежать або два критичні пороги, або у випадку домінування лише один (між рівнями аа і Аа — рис. 8). При цьому здається очевидним, що для упорядкованого здійснення кожного з батьківських фенотипів не потрібна абсолютно однакова інтенсивність дії «ізоаллельних» локус‑перемикачів (або тим більше ідентичність їх нуклеотидної структури). Достатньо лише, щоб сумарний ефект будь‑якої індивідуальної їх пари, взятої з лінії аа, не перевищував порогу, що допускає розвиток у напрямку відповідного фенотипу (рис. 8). Як у загальному випадку, так і при домінуванні це — поріг між градаціями аа і Аа. При схрещуванні гибридів першого покоління згідно з законами розходження хромосом у мейозі виникають, як відомо, зиготи з локусними комбінаціями усіх трьох типів у кількісних співвідношеннях 1:2:1, що означає реалізацію в тих же пропорціях трьох відповідних мереж перемикаючого фактора і визначуваних ними варіантів розвитку, тобто відбувається мендельське розщеплення за фенотипами. При наявності в системі лише одного порогу перемикання (домінування) виникають лише два вихідних стабілізованих варіанти у співвідношенні 3:1.
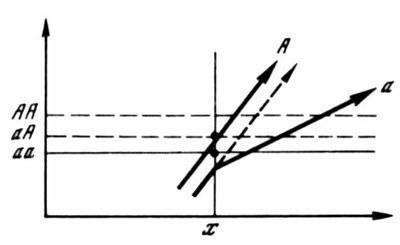
У першій критичній фазі два реалізованих варіанти розвитку дають співвідношення 3:1 залежно від того, чи перевищується порогове значення (аа) фактора I. У другій критичній фазі, де вибір визначається рівнем фактора II (з пороговим значенням вв), кожен з варіантів диференціюється на два нових — також у співвідношенні 3:1. В результаті виникає чотири класи фенотипів у відношенні 9:3:3:1. АА, Аа, аа, ВВ, Bв, вв — рівні перемикаючих факторів, визначені рекомбінацією двох пар локусів; АВ.., Ав.., аВ.., aв.. — класи здійснюваних фенотипів.
Будь‑яка упорядкованість біологічних явищ має розглядатися як продукт природного відбору (Майр, 1981), і щодо правил мендельського успадкування цей зв’язок цілком очевидний. Упорядкованість результату розвитку тут базується на наявності в епігенетичній системі батьків відповідних креодів, які можуть виникати лише внаслідок стабілізуючої дії відбору. Геномні мутації, як і інші одиничні впливи на систему, можуть, як показано, лише порушувати креоди, але самі по собі не створюють стійких шляхів розвитку. Відповідно, вираз «сирих» мутаційних аномалій, як природних, так і експериментальних, дуже змінний і загалом погано підкоряється мендельським правилам (Дубінін та ін., 1937; Гершензон, 1941; Камшилов, 1940; Шмальгаузен, 19686), що і призвело до виникнення понять генотипічного середовища гена, а також експресивності і пенетрантності мутантної ознаки. Лише ефекти найбільших геномних порушень показують відносно малу залежність від варіацій генотипу, тобто такі мутації досить однозначно детермінують певний тип аномалії; проте це відповідність ніколи не є повністю стабільною [Останнє, зокрема, стосується великих генетичних аномалій у людини, які, навіть будучи домінантними, іноді не проявляються у гомозиготі і демонструють загальну змінність вираження (наприклад, полідактилія; Гершкович, 1968)].
Зумовленість правильного успадкування попередньою стабілізацією схрещуваних фенотипів очевидна вже з того факту, що закони Менделя були встановлені саме на жорстко відселених (стабілізованих) лініях носіїв альтернативних ознак. Про специфічність умов, за яких виконуються ці закони, особливо наочно говорить зміст другого з них — закону незалежної рекомбінації, що вказує, що при успадкуванні кожна пара алельних факторів «веде себе як єдина» (Філіпченко, 1924), тобто завжди дає у другому поколінні гибридів розщеплення на два батьківські фенотипи на фоні будь‑яких варіантів рекомбінації всіх інших факторів. З точки зору хромосомної генетики визнання універсальності цього правила було б рівнозначним неприпустимому для неї твердженню, що характер вираження гена не залежить від варіацій генотипічного середовища. Очевидно, що така незалежність можлива лише за умови, що вплив рекомбінації на хід розвитку нейтралізовано попереднім відбором, так що при будь‑якому її варіанті зберігається лише вибір між двома можливостями, визначений у якийсь критичний момент розвитку станом лише однієї пари локусів. Відповідно, дійсно визнається, що необхідною передумовою генетичного аналізу, що спирається на мендельські правила успадкування, є попередній відбір ліній за аналізованими ознаками (Лобашев, 1966); і, зокрема, з цього починається аналіз мутантних фенотипів. У уявленнях генетики результатом такого відбору є створення чистої лінії носіїв мутантного алеля, тобто гомозиготної за тим локусом, з яким ототожнюється даний аномальний признак. З епігенетичної ж точки зору ця процедура означає стабілізацію раніше нестабільної траєкторії розвитку ознаки, засновану на реорганізації всього генотипу даної лінії (Waddington, 1957).
Постає питання: у чому ж полягає «вирішальна» роль однієї локусної пари у визначенні такого ознаки, виявлена аналізуючим схрещуванням? По‑перше, вона характеризує лише його упорядкований розвиток, створений селекцією, а не вихідний стан, у якому він був лише нестабільною аномалією. По‑друге, ця визначальна роль стосується не однозначного каналізованого розвитку ознаки, спостережуваного у батьківських лініях, а гібридного варіанту, де в одній епігенетичній системі поєднані дві різні можливості такого розвитку. Суть онтогенетичної дії «детермінуючої» локусної пари в системі гибрида полягає в тому, що на часовому відрізку, що відповідає вибору між двома траєкторіями, її ефект виходить за межі регуляції одного з альтернативних креодів, але залишається в межах норми для другого, так що все одно здійснюється один зі стабілізованих варіантів розвитку. Від перемикача тут залежить лише вибір шляху розвитку, а не саме його здійснення, визначене всім генотипом. Що стосується гомозиготності батьківської лінії за одним із членів «детермінуючої» пари, то, як уже сказано, це поняття означає не відсутність варіабельності локусу, а обмеженість її тими межами, при яких дія його подвійної дози не порушує стійкого розвитку ознаки, маркуючої дану лінію (рис. 8). Подібні межі варіабельності, що обмежують можливості каналізованого розвитку, існують у цій системі для будь‑якої пари локусів, тобто всі вони є в цьому сенсі «детермінантами» ознаки.
Отже, поняття мендельського гена передбачає певний стан видового епігенетичного системи, при якому в ній завдяки відбору стають можливими два стійкі варіанти розвитку ознаки, обраних залежно від поєднання станів певної пари хромосомних локусів. Тому закони їх рекомбінації, визначені поведінкою хромосом під час мейозу і запліднення (розходження і незалежне комбінування гомологів), отримують при такій організації системи однозначне вираження в успадкуванні альтернативних ознак, і ця однозначність становить сутність законів Менделя, тобто мендельський ген — це показник певної упорядкованості епігенетичної системи, а не сам хромосомний локус. Останній набуває властивостей перемикача алельних ознак лише в тій мірі, оскільки в системі розвитку гибридів існує сама можливість перемикання, що проявляється у наявності в ній двох стабілізованих креодів. Лише за цих умов набувають сенсу поняття гомо‑ і гетерозиготності, бо вони позначають ті градації епігенетичного ефекту локусної пари, які в даній системі визначають вибір дискретних шляхів розвитку; положення порогів між цими градаціями, як і наявність самих включених шляхів, характеризують систему, а не ділянку геному. При відсутності в системі другого креоду (тобто при існуванні у ознаки лише однієї норми) будь‑яка геномна зміна (рекомбінація, мутація) може мати лише два ефекти — або вона не зачіпає нормального розвитку, або призводить до нестабільних аберацій, що не демонструють правильного успадкування (як це спостерігається щодо природної невизначеної змінливості). Ця властивість аберацій означає, що для однокреодної системи поняття алеля взагалі втрачає зміст, оскільки в ній просто немає альтернативної можливості стійкого розвитку, що могла б бути активована при будь‑якій локусній зміні.
З сказаного випливає, що мендельський фактор — це не матеріальна частинка, а відношення між двома стійкими альтернативними станами епігенетичної системи («чистими лініями»), виявлене в гибридному аналізі. Це відношення не існує поза порівнянням зазначених станів. Кількість мендельських генів, яким визначається аналізований признак у даній системі схрещувань, означає не що інше, як кількість бінарних виборів між послідовними розгалуженнями каналізованих траєкторій, які мають бути здійснені в системі розвитку гибрида, щоб у підсумку отримати один із батьківських фенотипів (рис. 9). Легко бачити, що якщо кожен такий вибір залежить у даній системі від однієї пари локусів, що дає співвідношення 3:1 при реалізації двох відповідних траєкторій (креодів), то підсумкова кількість можливих результатів розвитку (тобто різних фенотипів) складе 2ⁿ, а їх кількісне співвідношення (3+1)ⁿ, де n — кількість послідовних критичних фаз, що відповідають розвилкам між дочірніми креодами, або, що те саме, кількість «детермінуючих генів» (рис. 9). Ці співвідношення і становлять сутність мендельського успадкування.
Все це робить очевидним, що поняття мендельського алеля і геномної мутації позначають зовсім різні явища, що належать до різних ієрархічних рівнів системи. Мендельський ген зі своїми алельними станами характеризує такий спосіб організації системи, при якому широкий спектр варіацій геному в цілому не виводить розвиток за межі обмеженого набору дискретних стійких шляхів; вибір серед них кожного разу залежить лише від сумарного рівня активності однієї визначеної пари гомологічних локусів. Навпаки, геномна мутація означає елементарне порушення в системі, яке може лише порушити її упорядкованість, але не здатне створити нову. Ототожнювати мендельський ген з ділянкою хромосоми, а алельну зміну — з мутацією цієї ділянки — означає наділяти елемент системи властивостями самої системи.
Це ототожнення, прийняте, хоча й не без коливань, моргановською хромосомною генетикою (Moran, 1937), змушує нас повернутися до питання про можливість зміни гена під дією відбору. Така можливість, ніби витікла з ряду експериментів (Castle, 1916), енергійно відкидалася ортодоксальним моргановським вченням, що бачило у гені локус, здатний лише спонтанно мутувати. Але навіть якщо мати на увазі саме локус, то в сучасних уявленнях це полінуклеотидна матриця, чутливість якої до відбору визнається в певній мірі навіть нейтралістською теорією молекулярної еволюції. Тим більше немає сумнівів щодо селективної обумовленості мендельських факторів (які і виявляються методами класичної генетики), адже, як показано вище, вони представляють собою просто символи епігенетичних співвідношень, створених відбором. Проще кажучи, мендельський ген — це продукт відбору (Шишкін, 1984в).
Найбільш ясним експериментальним доказом цього є досліди К. Воддінгтона з генетичної асиміляції (стабілізації) лабільних ознак (Waddington, 1957). Тут відбирали крила‑морфози у дрозофіли, такі як dumpy або bithorax, отримані тепловим впливом на личинку, після чого нащадки їх носіїв знову піддавали тепловому шоку на тій же стадії і відбору за тією ж дефінітивною аномалією. Після кількох поколінь відбору зазначені фенотипи стали проявлятися без дії шоку, і з них були отримані чисті лінії. Таким чином, ознаки стали стійкими (успадкованими). При схрещуванні їх носіїв з вихідною нормальною культурою вони відокремлювалися з тією чи іншою степенню пенетрантності (високою у bithorax), що дозволило розглядати їх як прояви відповідних мутантних алелів. Оскільки у вихідній лінії цих фенотипів явно не було, виникає питання: звідки з’явився новий ген? Можливості відповіді з традиційних позицій тут дуже обмежені. По‑перше, допускається, що отримані стійкі фенотипи є ефектами прихованих мутацій, що за звичайних умов лежать нижче порогу проявлення; відбір же об’єднав їх у полиген з більш сильним ефектом. Це найпоширеніше пояснення (Sheppard, 1970; Майр, 1974; Р’юз, 1977), змушене апелювати до припущень, що виходять за межі аналізу; проте і воно визнає, що детермінуючий комплексний фактор є продуктом відбору. Інша можливість — допустити існування гіпотетичного ефекту Болдуїна, тобто появу в ході кожного експерименту мутацій, що збігаються за вираженням з вихідним морфозом; але тоді сама незмінність їх появи в процесі відбору перетворюється на чергову загадку.
[Інтерпретація цих дослідів самим Воддінгтоном розкриває дуалізм його уявлень, що відображають прагнення поєднати епігенетичне пояснення стійкості фенотипу з ідеєю дискретної успадкованості. Як ембріолог‑феногенетик Воддінгтон усвідомлює, що отримання успадкованої ознаки, викликане генетичною асиміляцією, означає каналізацію її розвитку, зумовлену всім генотипом і не залежну від певного локуса, тобто пошуки останнього завідомо безпідставні. Але, як генетик, він все ж ставить питання: звідки виник ген, що визначає включення асимільованої ознаки? При цьому поняття перемикача креодів у гибридів несвідомо замінюється поняттям гена, що детермінує ознаку в асимільованій лінії.]У розглянутих випадках (фенотипи dumpy і bithorax) Воддінгтон, звичайно, не знаходить задовільної «генетичної» відповіді і змушений допускати тут появу випадкових мутацій у напрямку відбору (ефект Болдуїна) всупереч власному визначенню механізму генетичної асиміляції (Waddington, 1957, с. 166, 176, 180–182)].
Навпаки, епігенетичне пояснення експерименту не стикається з жодними труднощами, оскільки результати тут прямо відповідають очікуванням. Менделєвський ген у загальному випадку виявляється лише там, де є два стійкі альтернативні стани ознаки; ці два стани означають наявність у системі розвитку гибридів двох креодів; креоди ж створюються стабілізуючим відбором. Стабілізація первинно нестабільного морфозу в розглянутих експериментах призвела до створення лінії, у якій шлях його розвитку набув властивостей креода. Поєднання останнього в одній системі з попереднім креодом, що характеризує нормальну лінію (гібридний аналіз), призвело до здійснення двох альтернативних варіантів ознаки, тобто до менделєвського розщеплення. Відсутність повного прояву асимільованої ознаки вказує лише на недостатню стабільність її траєкторії.
Таким чином, можливість строго упорядкованих епігенетичних перемикань, що становить сутність менделєвського гена, виникла в результаті відбору. Тут мова йде, перш за все, не про створення відбором нового стану локуса, а про набуття цим локусом властивостей перемикача.
З усього сказаного легко зрозуміти, який сенс має вкладатися концепцією епігенетичної системи в процедуру менделєвського аналізу. Останній розкриває нам не «генетичну основу» досліджуваних ознак, а структуру цілісної видоспецифічної системи розвитку, яку ми «проявляємо» крок за кроком за допомогою стабілізації різних альтернативних траєкторій, ієрархічно будуючих цю систему і характеризуючих її потенційні можливості. Залежно від умов відбору одні й ті ж ознаки можуть бути стабілізовані різними шляхами, у результаті чого вибір між їх траєкторіями у гибридів буде залежати щоразу від різних локусів (що сприймається дослідниками як обумовленість даної ознаки в цих випадках неідентичними генами). Іншими словами, справжні інваріанти, що визначають вибір у ознаковому просторі даної системи, – це не стани хромосомних локусів, а ієрархічні послідовності розвилок (чутливих точок) онтогенетичних траєкторій, упорядковане перемикання яких можна організувати за допомогою відбору різними способами. Сукупність цих розгалужень і становить ландшафт системи.
Інваріантність загального рисунку траєкторій системи щодо їх потенційно можливих «пускових факторів» особливо наочно виявляється в експериментах з детермінантом статі. У більшості ксеногамних організмів альтернативні комбінації статевих ознак мають простий хромосомний механізм перемикання, тобто успадковуються як гетерозигота і рецесивна гомозигота за одним менделєвським фактором. При цьому визначення статі (наприклад, у риб і амфібій) в принципі можна повністю спотворити гормональними впливами або міжвидовим схрещуванням так, що зигота з генетичною конституцією самця розвивається в фенотипічну самку з нормальною репродуктивною здатністю і навпаки; іноді вдається за допомогою відбору перенести механізм визначення статі з статевих хромосом на аутосоми; нарешті, у ряді випадків у різних природних расах одного виду детермінанція статі може здійснюватися на основі протилежних варіантів гетерогаметності, тобто чоловічого — XY і жіночого — WZ (Астауров, 1966). Все це ясно показує, що здатність гомологічних геномних елементів перемикати розвиток є виразом структури самої системи, що допускає такі перемикання, і що у ролі диспетчерів ці гомологи можуть змінювати ролі, а також можуть бути замінені безліччю інших агентів з тим самим неспецифічним ефектом, що визначає вибір однієї з наявних каналізованих можливостей. Справжня причина, що визначає можливості вибору в такій системі, — це організація відповідного їй цілісного генотипу.
Розглянуті погляди на співвідношення хромосомних одиниць, менделєвських генів і ознак фенотипу не перебувають у такому різкому несумісності з теоретичними уявленнями хромосомної генетики, як це може здаватися. Несумісності слід скоріше шукати всередині самих цих уявлень, які об’єднують редукціоністське тлумачення числових співвідношень успадкування в групах особин («ознаки визначаються дискретними генами») і цілком системне розуміння ролі геномних одиниць у визначенні підсумку розвитку окремої особини («кожна ознака визначається всім геномом»). Основою першого з цих положень вважається менделєвський аналіз; основу другого складають дані експериментальної генетики, що показують, що одна й та сама ознака може порушуватись безліччю різнорідних мутацій і, отже, її нормальний розвиток залежить від функції багатьох локусів (Морган, 1924, 1937а, б, в; Воддінгтон, 1947). Останнє уявлення отримало свій завершений вигляд у теорії генетичного балансу (Bridges, 1922; Морган, 1937д), згідно з якою кожна ознака є виразом рівноваги, створюваної в розвитку дією всіх генів, а зміна в будь‑якому з них (мутація) призводить до нової рівноваги, що дає інший кінцевий продукт. Тому ген не створює ознаку, а лише схиляє загальний баланс взаємодії до того чи іншого фенотипічного результату. Таким чином, теорія визнає, що кінцевий ефект окремої хромосомної одиниці виражає не її власні властивості, а властивості всієї системи взаємодій, в рамках якої дана одиниця функціонує. Все це по суті дуже близько до визнання ідеї про неспецифічний вплив гена на підсумок розвитку, сформульованої пізніше Гольдшмідтом.
Ідея генетичного балансу становить основу уявлень хромосомної генетики про онтогенетичну («фізіологічну») дію гена (Морган, 1937д; Лобашев, 1963; Майр, 1968, 1974; Дубінін, 1976; тощо). Відповідно, уявлення про однозначний зв’язок між генами і ознаками формально відкидаються генетичною теорією; їх характеризують як помилку Вейсмана (Дубінін, 1966а, с. 238; Морган, 1937а, б, в), або як «генетику горохового мішка», що відображає позиції раннього менделізму (Майр, 1968, с. 216), або, нарешті, як некомпетентну думку, помилково приписувану генетиці (Тимофеєв‑Ресовський, Іванов, 1966, с. 116). Роль окремих генів зводиться лише до зсуву шляхів розвитку (Мюнтцинг, 1967, с. 73).
Проте прийняття такої позиції зобов’язує відповісти на питання: як у такому випадку мають інтерпретуватись результати менделєвського аналізу, що становить основу пізнавального інструменту класичної генетики? Останній, як відомо, побудований саме на однозначному проєкціонуванні генів на ознаки (Шмальгаузен, 1982; Столетов, 1967; Майр, 1968), і це дозволяє зрозуміти, чому принцип визначення ознаки всім генотипом іноді називають чисто теоретичною декларацією генетики, що не змінює її редукціоністських уявлень (Светлов, 1964). Насправді, чи можна узгодити обидва підходи? Якщо, наприклад, різниці двох константних (чистолінійних) фенотипів зводяться за результатами аналізу до різниці в одному гені, що розуміється як хромосомний локус, то перенести цей висновок на мову балансової гіпотези можна лише одним способом — визнати, що геноми всіх особин обох ліній абсолютно ідентичні, за винятком єдиного ділянки, яка зсуває генетичну рівновагу до одного з двох варіантів. Але це припущення неприйнятне, оскільки в дійсності геноми будь‑якої лабораторної лінії чи природної раси завжди варіабельні за багатьма локусами (ср.: Камшилов, 1939; Дубінін, 1966а). На практиці цих протиріч зазвичай навіть не помічають, тобто в інтерпретації менделєвського аналізу не вбачають особливої проблеми. Вказується, правда, що його конкретний результат не слід розуміти як виявлення всієї генетичної основи нормальної ознаки, яка завжди дуже складна. Аналіз кожного простого розщеплення виявляє лише окремий її елемент; повне ж уявлення про неї дає дослідження всіх можливих мутаційних аномалій ознаки. Але очевидно, що таке тлумачення несумісне з балансовою гіпотезою, оскільки в її уявленнях будь‑яка ознака особини має завжди одну й ту ж генетичну основу, а саме весь геном. Таким чином, очевидні два висновки. 1. Хромосомна генетика не містить пояснення менделєвського аналізу, сумісного з балансовою теорією. 2. Зведення монофакторної різниці між двома чистими лініями до зміни в одному хромосомному локусі не відображає фактичних різниць їх генотипів.
Отже, питання полягає в тому, чи можна взагалі уникнути поєднання в генетичній теорії двох взаємовиключних принципів: «ген (група генів) — ознака» і «геном (або генотип) — ознака»? Для цього логічно є лише одна можливість: визнати, що ген балансової гіпотези (хромосомний локус) і ген менделєвського аналізу — це не одне й те саме. Іншими словами, менделєвський фактор має виражати таку різницю між двома варіантами генотипу, яка принципово не зводиться до властивостей чи станів їх окремих елементів. Саме до цього висновку ми і прийшли вище, спираючись на теорію епігенетичної системи і відповідаючи їй реальну феноменологію успадкування. Правильне менделєвське розщеплення виражає взаємовідношення двох станів системи, що відповідають двом альтернативам каналізованого розвитку. Вибір між ними визначається у гибридів кількісним рівнем активності однієї гомологічної пари локусів, яка у цьому випадку виконує роль «диференціатора». Ця роль, виражена символом менделєвського гена, забезпечується виключно властивістю гібридних генотипів здійснювати два стійкі варіанти розвитку; при втраті цього властивості зникає і сама упорядкованість успадкування, що дозволяє виділити його контрольний фактор.
Це уявлення про сутність гена в основі своїй не є новим для генетики. Приблизно таке саме було і у автора терміну «ген» Йогансена (Johannsen, 1926), який розумів під ним не частинку успадкованого матеріалу, а одиницю різниці двох генотипів (Baur, 1922). Досягнення хромосомної теорії успадкування в очах багатьох дослідників, особливо феногенетиків, не скасовували основу такого уявлення про ген. Останній, навіть будучи віднесений до хромосоми, розглядався ними як причина різниці в розвитку, що не існує поза властивостями цілісного генотипу (Промптов, 1934; Камшилов, 1934); припущення про самостійну дію гена прирівнювалося до можливості «ізольованого існування дірки від бублика» (Промптов, 1934). За словами Камшилова (1935, с. 141), ген — це не локус, а відмінність двох цілісних генотипів, «виявлена у своєрідному типі розвитку». Навіть для Моргана визнання хромосомної локалізації гена супроводжувалося певними коливаннями, і певний час допускалося, що мова йде лише про абстрактну властивість, якимось чином пов’язану з даним ділянкою хромосоми (Морган, 1937г). У наш час з розвитком біохімічних методів генетичного аналізу, де локус виступає як одиниця матричного синтезу, його нетождественність менделєвському гену знову стає для генетиків усе більш очевидною (Grant, 1980, с. 290; Holubovskyi, 1982).
Визнання системної обумовленості «генних» властивостей локуса частково знайшло відображення і в гіпотезі Фішера (Fisher, 1930) про еволюцію домінантності. За останньою, здатність локуса детермінувати ознаку у присутності свого неідентичного за властивостями гомолога (тобто в гетерозиготі) є результатом його взаємодії з системою модифікаторів, створеної відбором. Таким чином визнається, що принаймні класична картина менделєвського розщеплення за фенотипами (3:1) не є властивістю гомологічних хромосомних одиниць і виникає лише як продукт реорганізації відбором усього генотипу. Звідси лишається лише крок до поширення цього висновку на будь‑яке упорядковане відповідність між локусом і ознакою, виявлену гібридним аналізом.
З іншого боку, перший крок до усвідомлення того факту, що в основі вибору між алельними станами лежать якісь кількісні різниці, був зроблений ще ранніми менделістами, отримавши вираз у відомій гіпотезі «наявності‑відсутності» Бетсона‑Пеннета, запропонованій для пояснення домінування (домінантна ознака визначається наявністю фактора в гаметах, рецесивна — його відсутністю). Критика зі сторони морганівської школи, що вказала на несумісність цього погляду з явищем множинного алелізму і уявленням про лінійне розташування генів у хромосомах, призвела до нової формулювання гіпотези, у якій ще чіткіше говорилося про кількісний характер алельного різниця (рецесивний фактор — це щось, що втратило порівняно з домінантним; Пеннет, 1930). І нарешті, Гольдшмідт, незалежно обґрунтувавши ідею про «кількість гена» як основу алельних змін, розвинув її у уявленні про порогові рівні морфогенетичного ефекту, що визначають вибір фенотипу у критичних точках розвитку (Goldschmidt, 1927, 1938, 1940).
Як показано вище, наявність у хромосомної одиниці властивостей менделєвського фактора означає, що дана структура генотипу допускає кілька стійких шляхів розвитку ознаки. При відсутності такого вибору, тобто можливості перемикання стабілізованих траєкторій, не може бути і локуса‑перемикача, тобто гена, «визначального» ознаки. Отже, у загальному випадку роль окремого локуса в детермінанті властивостей фенотипу залишається невизначеною і може бути описана лише як елемент у складній системі функціонування всього генотипу в процесі розвитку.
Але якщо окремий локус не має самостійного вираження у фенотипі, а останній у кожній своїй ознаці визначається всією зародильною плазмою (генотипом), то в чому тоді полягає принцип корпускулярної (дискретної) успадкованості, що лежить в основі хромосомної теорії? Актуальність цього питання стала очевидною для Моргана ще у 1918 р., до появи балансової гіпотези, і його відповідь заслуговує уваги. Він стверджує, що якщо зазначені співвідношення між ознаками і зародильною плазмою дійсно мають місце, то остання все ж у будь‑якому випадку залишається побудованою з елементів, незалежних щодо мутації, кросинговеру, а також розходження гомологів і комбінування їх пар у процесі дозрівання статевих клітин (Морган, 1924, с. 232, 235). «У цьому, і лише в цьому сенсі, ми маємо право говорити про корпускулярну будову плазми і корпускулярну успадкованість» (там же, с. 235). Ці визнання вражають. З них випливає, що морганівська теорія успадкованості описує лише закономірності перерозподілу хромосомних елементів, але при цьому не знає, яким чином вони можуть бути пов’язані з успадкуванням ознак! І таким чином знову опосередковано визнається, що менделєвські правила успадкованості ознак описують щось зовсім інше, ніж елементарні властивості хромосомних одиниць.
Відсутність методу для інтерпретації підсумку розвитку в термінах дискретних хромосомних генів, неявно визнаного Морганом, не є наслідком неповноти знань, як часто вважають. Воно має принциповий характер. Дані ембріології не залишають сумнівів, що розвиток — це нерозкладний епігенетичний процес, заснований на взаємодії всіх його елементарних факторів і безперервному збільшенні якісного різноманіття. Явища цитоплазматичної прелокалізації зачатків і пов’язаних з нею фаз преформованого (мозаїчного) розвитку становлять тут лише часткові епізоди, обумовлені епігенетичними взаємодіями в оогенезі і знову змінювані в ході розвитку регуляторними процесами (Шпеманн, 1925; Девідсон, 1972; Светлов, 1978). Детермінанція частин у розвитку визначається лише детермінанцією цілого, і ознаки дорослого організму не можуть мати корелятів у зиготі чи її геномі (Гурвіч, 1944; Светлов, 1964, 1978). Як ми бачили, будь‑який підсумок розвитку — нормальний чи аберантний — завжди є еквіфінальним щодо варіацій значень його елементарних причинних факторів. Стійкість ознак «не властивість генів, а вираз взаємозалежностей частин у кореляційних системах розвиваючогося організму» (Шмальгауен, 1982, с. 174). Ця несводимість процесу розвитку до преформаційної моделі, що розглядає організм як суму наслідків незалежних початкових причин, давно зрозуміла експериментальною ембріологією (Бляхер та ін., 1935; Белоусов, 1979) і формально визнається також хромосомною генетикою, що бачить у цьому одне зі своїх головних відмінностей від вейсманівської теорії зародильної плазми (Морган, 1937а, б; Дубінін, 1966а).
Як уже зазначалося, відсутність специфічної ролі індивідуальних хромосомних генів (локусів) у визначенні властивостей фенотипу найповніше продемонстровано Гольдшмідтом, і він виявився одним із небагатьох провідних генетиків, хто ясно побачив, до яких наслідків це веде для теорії розвитку (а отже й успадкованості). «Факти генетики, звичайно, можуть описуватись у термінах генів, але теорія зародильної плазми повинна повністю звільнитися від концепції генів як одиниць» (Goldschmidt, 1938, р. 311), тобто генотип є нерозкладною основою розвитку фенотипу. «Зародишова плазма як ціле контролює певну реактивну систему, яка є не мозаїкою окремих ефектів, а єдиною системою розвитку, керованою як ціле одним фактором… Для багатьох генетиків явно важко мислити в таких поняттях, оскільки більшість їх настільки пов’язані аксіоматичною вірою в атомістичну генну теорію, що не в змозі думати інакше; але ембріологи, фізіологи і, можливо, систематики не знайдуть труднощів у прийнятті цієї концепції» (Goldschmidt, 1940, р. 218).
Прогнози Гольдшмідта щодо прийняття генетикою цих поглядів (які, здавалося б, підходили їй близько) цілком виправдалися, і причини цього зрозумілі.Системні узагальнення генетики, у яких не залишається місця для генів як детермінантів ознак, цілком належать до її теоретичних уявлень про індивідуальний розвиток, тобто тієї області, де вона не має особливо великих успіхів. Навпаки, редукціоністський підхід до вивчення спадковості, пов’язаний із застосуванням менделєвського аналізу, є основою всіх тих величезних і очевидних практичних досягнень, які дозволили генетиці претендувати на роль точної науки і завоювали їй небувалий авторитет у біології ХХ ст. Не дивно, що в цих умовах, коли головним щоденним завданням залишається встановлення зв’язків між геномними одиницями та ознаками організму, здається недоречним згадувати про те, що в теорії за генами не визнаються такі детермінуючі властивості і що їх слід відносити лише до генотипу. Самоочевидність виявлених дискретних зв’язків і можливість їх експериментальної перевірки зазвичай заважає задуматися над тим, якою насправді є їх природа і за яких умов вони виникають. Властивість локуса перемикати шляхи розвитку при схрещуванні двох стабілізованих відбором фенотипів абсолютизується як його постійна властивість, а сама його функція перемикача, виявлена лише у гибридів, розглядається як доказ його особливої детермінуючої ролі щодо даної ознаки. Одним із красномовних свідчень тріумфу цього редукціоністського стилю мислення є класичне твердження, що принцип «чистоти гамет» сам по собі забезпечує дискретність успадкування (незмішуваність) ознак, тобто що закони розходження хромосомних гомологів повинні завжди прямо відображатися у властивостях фенотипів, незалежно від стійкості їх розвитку.
Необхідність ув’язати ці уявлення з системними вимогами балансової гіпотези на практиці виливається в «компромісне» рішення, тобто концепцію генотипічного середовища. Участь усього генотипу в реалізації ознаки розуміється тут як взаємодія «основного» гена (або генів), що визначає ознаку, з рештою спадкових елементів, які модифікують його специфічну дію і грають щодо нього роль «шумів». Таке розуміння співвідношень гена і ознаки закріпилося в умах багатьох дослідників як типове для хромосомної генетики незалежно від того, відкидають вони його (Goldschmidt, 1940) чи вважають прийнятним (наприклад, Астауров, 1971, с. 218; Bertalanffy, 1969, р. 73). В цілому очевидно, що ця інтерпретація цілком залишається в рамках преформістських уявлень про дискретну (мозаїчну) детермінацію ознак. По суті вона мало відрізняється від концепції Вейсмана, у якій можливість прояву кожного детермінанта подібним же чином залежала від загального результату боротьби спадкових зачатків. Це, здавалося б, постійно відкинута, але насправді міцно вкорінена віра в незалежне визначення ознак хромосомними генами косвенно виявляється навіть там, де словами підкреслюється існування у видового генотипу та контрольованої ним епігенетичної системи особливих цілісних властивостей, не порушуваних елементарними мутаціями. Бо коли мова йде про порівняння таких генотипів, їхні відмінності насправді оцінюються не в цілісних властивостях, а лише в кількості неідентичних генів (Dobzhansky, 1947, с. 106, 110, 337, 338; Майр, 1968, с. 432). Сутність генотипу як цілого, виражена лише через індивідуальний розвиток (Камшилов, 1934), і надалі зводиться тут до суми хромосомних елементів.
* * *
В історичному плані появу кожної нової галузі природничих наук зазвичай починає осягнення предмета дослідження аналітичними методами і, як наслідок, домінування редукціоністського підходу до його опису; час системних узагальнень приходить пізніше. Так було і в історії традиційних експериментальних галузей біології — механіки розвитку і генетики, з яких остання пройшла шлях від типічної «механіки спадковості» (Вейсман і ранні менделєвці) до системних побудов Голдшміда, Шмальгаузена і Воддінгтона. Очевидно тому, що появу і бурхливий розвиток нового фронту досліджень — молекулярної генетики, що вивчає будову і функції матричних структур клітини, знову привели до відродження редукціоністських уявлень про зв’язок гена і ознаки, виражених тепер у формулі «один ген — один фермент». Це внесло новий елемент дуалізму у світогляд класичної генетики, багато представників якої, заперечуючи в теорії пряму детермінацію генами ознак фенотипу, одночасно змушені допускати її можливість щодо первинних генних продуктів (Тимофеєв‑Ресовський, Іванов, 1966; Waddington, 1966); останні (передусім білки) розглядаються як ознаки, які найадекватніше відображають свою генетичну основу (Гершенсон, 1974). Все це не могло сприяти зміцненню системних поглядів у класичній генетиці, вплив якої й так був обмеженим. Однак і в молекулярній біології тим часом починають звучати нові ноти. Давнє висловлення, що принцип «ген‑фермент» представляє крайнє спрощення реальних співвідношень (Haldane, 1954), тепер знаходить усе більше підтверджень. Явища поліморфізму ферментів і схожість просторової структури їх гомологів у різних організмів вже самі по собі змушують думати, що саме ця структура грає головну роль у визначенні функціональних властивостей білка, тоді як утворюючі його амінокислотні послідовності (принаймні в ділянках, що лежать поза активними центрами) можуть варіювати. Таким чином, існує виродженість відповідності не лише між нуклеотидним кодом ДНК і лінійною структурою білка, а й між останньою і характером його функції (Waddington, 1970b; Волкенштейн, 1981a, b). Більш спеціалізовані дослідження показують, що весь багатоступеневий процес, що веде від цитрону до ферменту, тобто транскрипція‑процесінг (посттранскрипційна редукція іРНК) — трансляція і посттрансляційні модифікації — є поліваріантним на кожному своєму етапі (Інге‑Вечтомов, 1976; Михайлова, Симаров тощо, 1981). Одним із яскравих проявів цієї поліваріантності є той факт, що в нормі з одного і того ж сегмента ДНК можуть зчитуватися в різних тканинах або на різних стадіях онтогенезу різні транскрипти і, відповідно, утворюються різні білки (Голубовський, 1985). В цілому така неоднозначність може бути обумовлена впливами найрізноманітнішої природи, що створює, з одного боку, можливість відхилення кінцевого продукту синтезу від норми, а з іншого — створює основу для його регуляції («фенотипічного супресування» мутацій) при помилках коду. Така регуляція, зокрема, продемонстрована у дріжджів‑цукрових грибків для посттранскрипційних етапів синтезу, де її можна викликати зміною температури, pH, осмотичного тиску та дією інших зовнішніх агентів (стрептоміцину, гліцерину тощо). Ці процеси запобігають втратам у структурі поліпептидного ланцюга і полегшують його функціональну регуляцію на посттрансляційному рівні (Михайлова, Симаров тощо, 1981). Остання може експериментально індукуватися тими ж агентами, що і посттранскрипційна регуляція, але у багатьох випадках вона спостерігається як природний процес, що призводить до збереження мутантом нормальної біохімічної функції. При цьому часто має місце не виправлення помилок у амінокислотних послідовностях, а нивелювання їх ефекту. Найвідоміший механізм цієї регуляції, вивчений у різних організмів, — взаємодія продукту активності мутантного локуса з нормальними або мутантними субодиницями того ж білка («міжаллельна комплементація»). Вона призводить або до поповнення втрат у первинній структурі однієї поліпептидної ланцюга за рахунок інших, або до відновлення функції цілого за рахунок конформаційних змін мутантних субодиниць, тобто до такого об’єднання наявних фрагментів, що їх комплекс набуває нормальної просторової структури шляхом самозбирання. При цьому амінокислотні зміни поза активними центрами не порушують нормальної функції (Інге‑Вечтомов, Сойдла, 1978).
Явища комплементації продуктів синтезу, що порушують колінеарне відповідність між цитроном і ферментом, проливають світло на еволюційне значення посттрансляційних змін загалом. Той факт, що активні центри білків зберігають відносну автономність і часто можуть функціонувати незалежно від їх зв’язку (наприклад, дві неідентичні субодиниці триптофансинтетази, розділені у кишкової палички, але закодовані як одне ціле у нейроспори), змушує дослідників вважати, що багато мультифункціональних білків могли виникати шляхом об’єднання ферментів, закодованих різними генами, і, навпаки, що мультиферментні комплекси в ряді випадків є продуктами епігенетичного розщеплення мультифункціональних білків‑попередників, як це прямо спостерігається у деяких вірусів (Інге‑Вечтомов, Сойдла, 1978). Тут вже не треба говорити про правило «ген‑фермент». Необхідність враховувати можливий ефект посттрансляційних модифікацій стає все більш очевидною для дослідників, змушуючи утриматися від прямого ототожнення рядів ізоферментів з продуктами ізоаллельних мутацій (Солбриг, Солбриг, 1982, с. 256).
Усі ці факти показують, що, незважаючи на велику відстань, що розділяє локус і фенотип у класичній і молекулярній генетиці, їхні взаємовідносини в принципі схожі. Функціональну стійкість кінцевого продукту синтезу неможливо звести тут до стійкості самих матриць (ДНК і РНК). Вона базується на регуляторних епігенетичних взаємодіях, що охоплюють всю систему синтезу і здатні буферизувати певні помилки генетичного коду, а також порушення процесів транскрипції і трансляції. Як і в «макроонтогенезі», регуляційні можливості цієї системи чутливі до зовнішніх факторів. Синтезована молекула в такій же мірі є продуктом цитрону, як і всіх елементів системи синтезу, що не залежать від нього (ферментів, РНК тощо), і не обов’язково колінеарна цитрону. Все це змушує вважати, що і еволюційний механізм становлення білків в принципі є тим самим, що і у інших елементів адаптивної норми. Білкова молекула також має «фенотип», частково визначений середовищем, і її відповідність субстрату дії має бути історично стабілізована (Waddington, 1970b). Очевидно, що першим кроком еволюційного становлення нового типу білка має бути його виникнення як однієї з нестабільних посттрансляційних модифікацій існуючого ферменту. Якщо остання виявилася в нових умовах адаптивно цінною, то шлях її реалізації перетворюється відбором у напрямку максимальної стійкості до перешкод. Очевидно, це проявляється передусім у поступовому вирівнюванні і спрощенні всієї послідовності етапів синтезу, тобто у встановленні все більшої колінеарності між новою молекулою і її вихідною матрицею. З цієї точки зору випадки упорядкованих посттрансляційних перетворень ряду нормальних білків можна інтерпретувати як проміжні етапи стабілізації їх морфогенезу, «рекапітулюючи» хід первинно нестабільних перетворень. До таких прикладів належить, зокрема, утворення у ссавців інсуліну шляхом протеолізу гігантської молекули‑попередника і розщеплення її на дві субодиниці, або ж утворення основного білка у фаґу Т4, де близько 900 однакових поліпептидних ланцюгів спочатку спонтанно агрегуються, а потім відщеплюються N‑кінцеві ділянки (Стент, Келіндар, 1981). Ймовірно, також вирізання неактивних (інтронних) послідовностей з матричної РНК, що становить звичайний етап білкового синтезу, є відображенням перетворень, що колись відбувалися на посттрансляційному рівні. Все це змушує вважати, що матриця ДНК, що лежить в основі стабільно синтезованого білка, історично є не причиною його виникнення, а, навпаки, наслідком стабілізації його морфогенезу, первинно заснованого на модифікації продукту, закодованого іншим варіантом матриці. Якщо це вірно, то ми повинні зробити висновок, що пов’язувати історичне появу нових гомологічних білкових субодиниць, наприклад b‑гемоглобіну, з дуплікацією відповідного гена (як це зазвичай роблять) — означає міняти місцями причини і наслідки; онтогенетична і історична причинність не тотожні. До частково схожих висновків прийшов Ю. М. Оленев (1977), зазначивши, що потреба в новому продукті виникає раніше, ніж подвоюється відповідний ген, і що ця проблема в еволюції вирішується спочатку обхідними шляхами. Можливість того, що розширення функцій гена може передувати його дуплікації, визнає також Т. Р. Сойдла (1983).
Отже, механізм матричного синтезу може бути описаний у своїй основі як система, регульована до певного кінцевого стану і, очевидно, перетворювана в еволюції «зверху вниз» (від кінцевого продукту до матриці), тобто у відповідності з теорією стабілізуючого відбору. Можна погодитися з Б. М. Медниковим (усне повідомлення), який вважає, що білок по суті є органом, до розвитку якого застосовні всі закономірності, справедливі для звичайних органів.
Слід додати ще одне міркування. Ефект біохімічних мутацій може фенокопіюватися (Goldschmidt, 1955), і якщо це правило так само універсальне, як щодо звичайних фенотипів, то треба зробити висновок, що будь‑який продукт синтезу, отриманий на основі одиничної мутації даного генотипу, може бути в принципі отриманий на тій же основі і без мутації, за допомогою підбору зовнішніх впливів, певним чином порушуючих транскрипцію, трансляцію або пізній епігенез. Зазвичай такі випадки індукованого синтезу пояснюються депресією оперонного механізму, що містить необхідну матрицю, згідно зі схемою Жакоба і Моно (Waddington, 1964; Волкенштейн, 1981); але в світлі міркувань, викладених вище, допустимо вважати, що це явище має більш загальну основу — обмеженість спектра потенційних можливостей первинного синтезу, допустимих для даного генотипу як цілісної видової системи при всіх його елементарних мутаційних змінах.
ЕПІГЕНЕТИЧНА ТЕОРІЯ ЕВОЛЮЦІЇ
Вище підкреслювалося, що питання про походження нормально‑адаптивної організації, що становить головну проблему еволюційної теорії, зводиться до пояснення того, яким чином нові властивості організмів набувають стійкості, стаючи незворотними. Це пояснення в загальній формі дає концепція епігенетичної системи, в рамках якої елементарна еволюційна зміна означає перехід індивідуального розвитку на одну з аберантних траєкторій з подальшим перетворенням її у каналізований шлях розвитку (креод). Теорія стабілізуючого (каналізуючого) відбору, що розглядає такий хід подій як основу всього еволюційного процесу, може називатися епігенетичною теорією еволюції (Шишкін, 1984a). Ця назва виправдана, по‑перше, тому, що ініціуючим фактором еволюції тут визнаються порушення ходу онтогенезу. По‑друге, вона дозволяє уникнути помилкового уявлення, згідно з яким стабілізуючий відбір є лише одним із часткових еволюційних механізмів поряд з «ведучим», «дизруптивним» та іншими формами відбору, а вчення Шмальгаузена‑Воддінгтона становить просто розділ синтетичної теорії, що розглядає зміни у стабільному або коливному середовищі. Хоча відмінності епігенетичної теорії від традиційних поглядів і підкреслювали її автори, вони все ж ніколи не узагальнювалися, а саме її виклад залишався в значній мірі забрудненим чужими для неї поняттями. Тому спочатку необхідно коротко охарактеризувати її зміст.
У основі епігенетичної теорії лежить уявлення про адаптивну норму, або типову організацію, як об’єкт еволюційних змін. Під адаптивністю, або доцільністю організації, розуміється її здатність до самопідтримки і самовідтворення (наслідування), тобто її стійкість. Остання, у свою чергу, виражається у здатності нормального індивідуального розвитку релаксувати в широких межах зовнішні та внутрішні збурення на шляху до здійснення даної зрілої організації. Пояснення цієї властивості розвитку і має становити ключове завдання еволюційної теорії.
Епігенетична концепція вирішує цю проблему, виходячи з уявлення, що видоспецифічне індивідуальне розвиток є цілісною динамічною системою з обмеженим і структурованим простором можливих кінцевих станів, серед яких нормальний (адаптивний) фінал відповідає рівновазі системи, а вся область потенційних фенотипічних відхилень — її більш‑менш нестабільним флюктуаціям. Будь‑яке відхилення підсумку розвитку від рівноваги у відповідь на пошкоджуючий вплив завжди представляє собою системну реакцію, вибір якої визначається не специфікою пошкоджуючого фактора (наприклад, типом мутації), а лише мірою, місцем і часом збурення, внесеного ним у хід розвитку.
Уся сукупність можливих станів системи (рис. 1, 7), або її фазовий простір (епігенетичний ландшафт), становить її цілісну характеристику, визначувану загальною організацією (генотипом) зародкової клітини і не розкладну на незалежні ефекти дії якихось елементарних факторів всередині цієї клітини або її онтогенетичних похідних (Гурвіч, 1944; Goldschmidt, 1940; Waddington, 1957). Іншими словами, можливі альтернативні стани видового фенотипу — це параметри системи розвитку, а стани її окремих взаємодіючих елементів (починаючи з елементів структури геному) — динамічні змінні, що характеризують нижчий ієрархічний рівень системи. Огромному різноманіттю комбінацій значень елементарних складових (що варіюють у кожному індивідуальному циклі розвитку) відповідає на верхньому рівні обмежений простір варіацій фенотипічних параметрів, тобто останні утворюють щодо цього різноманіття інваріантну (еквіфінальну) сукупність.{
"translated_text": "При цьому чим вище відносна успішність (ймовірність здійснення) даної фенотипічної аберації, тим ширший спектр можливих станів нижчого рівня, в рамках якого вона реалізується. Нормальний фенотип, що відповідає рівновазі системи, за ступенем еквіфінальності різко переважає всі інші, тобто в нормальних умовах він реалізується подавляючою більшістю індивідуальних варіантів зигот, що існують у природних популяціях.\nЦі уявлення про системний механізм індивідуального розвитку спираються на досвід експериментальної ембріології, популяційної генетики та феногенетики, який показав високу стійкість нормального розвитку щодо варіацій морфогенетичних процесів і генетичної структури зародкової клітини, а також взаємозамінність різних комбінацій зовнішніх і внутрішніх факторів при здійсненні одних і тих самих аберантних фенотипів. Єдиною альтернативою такому розумінню розвитку може бути уявлення, згідно з яким кожна особливість (або стан) фенотипу детермінується в онтогенезі своїм особливим причинним фактором або їх сумою. Це — преформістська модель розвитку Ру‑Вейсмана, несумісність якої була доведена експериментальною ембріологією.\nПодані погляди на природу індивідуального розвитку передбачають ряд неминучих наслідків.\n1. Жодне порушення в системі розвитку, викликане внутрішнім або зовнішнім агентом (мутація, вплив середовища) не в змозі змінити властивості самої системи. Воно або релаксується в ході індивідуального розвитку, або призводить до зміни вибору онтогенетичної траєкторії в межах, визначених простором можливостей даної системи. Таким чином, мутація не може бути прирівняна до еволюційної зміни (перетворення системи).\n2. Мутація (як і будь‑яке порушення) не детермінує певного фенотипу, оскільки кожен з них є цілісною реакцією системи, здійснюваною при різних комбінаціях значень елементарних взаємодіючих причинних факторів. У свою чергу, кожне з цих значень може за певних умов співучаствувати у здійсненні будь‑якого з фенотипів, «дозволених» даною системою. Тому жодна фенотипічна зміна не може бути описана в редукціоністських поняттях (зокрема, і в термінах алельних станів генів).\n3. Всі індивідуальні результати розвитку, реалізовані даною видоспецифічною системою, базуються на більш‑або менш ідентичних комбінаціях елементарних причинних факторів (як у випадку ксеногамних популяцій, так і клонів). Тому будь‑яка група ізореагентів (циклів розвитку з однаковим фенотипічним результатом) завжди представляє собою генетично неоднорідну вибірку.\n4. Будь‑яке відхилення розвитку від рівноважної траєкторії, незалежно від викликаної ним елементарної причини (мутація або зовнішній вплив), є порушенням стійкості даної індивідуальної системи. Це означає більшу або меншу нестабільність відтворення (успадкування) відповідного аберантного фенотипу в ряді нащадків даної особини і, в кінцевому підсумку, його витіснення або поглинання більш стійкою нормою (тобто релаксацію відхилення на популяційному рівні).\nОтже, справжня еволюційна зміна — це зміна структури системи розвитку (характеризованої певним рисунком допустимих для неї онтогенетичних траєкторій), а не флуктуація в межах існуючої структури, до якої зводиться ефект елементарних мутацій. Саме розуміння суті цієї проблеми привело колись Голдшмідта (Goldschmidt, 1940) до ідеї «системних мутацій», що передбачає реорганізацію системи шляхом стрибкоподібної перебудови всього хромосомного апарату зародкової клітини. По суті це була невдала відповідь на правильно поставлене питання про механізм еволюційного перетворення простору можливостей розвитку.\nЕпігенетична концепція вирішує цю проблему інакше. Перш за все підкреслимо, що перебудова системи розвитку означає зміну її рівноважного стану (адаптивної норми) і тим самим — появу нової каналізованої онтогенетичної траєкторії, що реалізує цей стан. Положення останньої у фазовому просторі системи, у свою чергу, визначає властивості самого цього простору, оскільки всі існуючі в ньому аберантні шляхи розвитку є відгалуженнями каналізованої траєкторії. У перекладі на мову традиційних понять це означає, що характер адаптивного фенотипу визначає спектр аберантної змінливості.\nТаким чином, зсув адаптивної норми завжди є показником перебудови структури (епігенетичного ландшафту) системи. При цьому очевидно, що між будь‑якими двома послідовними історичними станами цієї структури (які відповідають мінімальному зсуву норми) має існувати пряма преемність. Нова траєкторія рівноваги не з’являється з нуля і повинна відповідати одному з аберантних онтогенетичних шляхів, що існували в рамках попередньої структури (рис. 7). Її стабілізація розглядається теорією як результат природного відбору на користь відповідної фенотипічної аберації (морфози), що здійснюється частиною зигот у екстремальних умовах розвитку. Процес відбору має вести до поступового зростання надійності реалізації даного відхилення в ряді поколінь, тобто до підвищення його успадкованості. Цей ефект отримав назву «генетичної асиміляції ознак» (Waddington, 1953, 1957). Таким чином, перебудова системи розвитку, починаючи щоразу з вибору одного з фенотипічних варіантів, нестабільно реалізованих у межах наявної організації видового генотипу, зрештою призводить до перетворення самої цієї організації, тобто система перебудовується в напрямку від дорослого фенотипу до зиготи (Шишкін, 1981, 1984а, б). «Не зміни генотипу визначають еволюцію і її напрямок. Навпаки, еволюція організму визначає зміну його генотипу» (Шмальгаузен, 1940б, с. 57). У підсумку еволюція розглядається як процес репарації стійкості норми, періодично розшатаної історичними змінами середовища; ця репарація досягається щоразу ціною селективного перетворення самої норми разом із реалізуючою її системою розвитку.\nЗупинимось тепер докладніше на механізмі встановлення нової рівноваги системи, або, іншими словами, на стабілізуючому дії відбору. Будь‑яка незворотна зміна зовнішніх умов у бік межі, що обмежує можливості каналізованого розвитку в межах даної системи, веде до реалізації рознорідних відхилень, спектр і відносна частота яких визначаються властивостями цієї системи. Будь‑яка різниця цих фенотипів за життєздатністю в даних умовах дає селективну перевагу певному їх класу. Але оскільки цей фенотип, як і всі інші реалізовані класи аберацій, належить до нерівноважних станів системи, його відтворення на початкових етапах селекції виявляється надзвичайно нестабільним, тобто належні до нього ізореагенти практично повторюють у своєму потомстві весь той спектр відхилень, з якого вони самі були відібрані. Відбір на цьому етапі ніби черпає решето води. Збереження адаптивно цінної аберації в ряді поколінь тут зумовлене не стільки більшою ефективністю її індивідуального успадкування у порівнянні з іншими морфозами, скільки обмеженістю самого аберантного простору даної системи, внаслідок чого відбираний варіант знову незмінно відтворюється нею серед інших.\nКожен елементарний крок відбору, що охоплює два покоління, означає переважне збереження особин, які змогли відтворити фенотип своїх раніше відібраних батьків, незважаючи на комбінування їх гамет при схрещуванні (у ксеногамних організмів) та різні інші генетичні зміни в процесі самого гаметоутворення (мейотична рекомбінація та помилки реплікації). Тому історія будь‑якого фенотипу, збереженого тривалим відбором, — це ланцюг послідовних випробувань його носіїв на здатність відтворювати самих себе в умовах безперервної зміни простору варіацій їх геномів. У результаті, чим більшим числом поколінь відбору відокремлені такі носії від вихідної групи ізореагентів, що дала їм початок, тим більший розмах генетичних змін ця лінія змогла витримати, не змінюючи свого фенотипу. Це означає, що генотип новоствореної («асимільованої») лінії перебудовується відбором у напрямку все більш стійкого здійснення даного фенотипу. Відповідно, останній починає все ефективніше поглинати у схрещуваннях інші аберації, властиві даній системі розвитку, тобто реагує на їх гібридизацію як на регульовані онтогенетичні перешкоди. Збережене відхилення стає, таким чином, все більш однозначно успадкованим, перетворюючись у нову норму.\nЦе перетворення норми за допомогою відбору по суті являє собою вираження загальної здатності системних об’єктів релаксувати викликані в них порушення, тобто змінюватися цілеспрямовано. Відновлення рівноваги, або «пошук цілі» (Ешбі, 1962), здійснюється системою шляхом послідовної корекції її стану, що веде до затухання початкового порушення. Саме така ситуація, але лише пов’язана з якісною зміною самої системи, виникає в процесі еволюційного зсуву норми. При переході популяції в екстремальні умови її система розвитку дестабілізується і замість реалізації попередньої норми переходить до хаотичних і нестабільних індивідуальних флуктуацій. Подальше виживання системи в нових умовах залежить від того, чи зможе вона стабілізуватися в одному з цих змінних станів. Цей пошук нової рівноваги здійснюється системою шляхом переважного збереження (відбору) індивідуальних варіантів розвитку, що реалізують найжиттєздатнішу флуктуацію. Процес відбору тут не є нічим іншим, як ланцюгом затухаючих циклів корекції з зворотним зв’язком, що призводить до стабілізації нової норми. Кожен акт збереження відбором носіїв адаптивно цінної аберації — це зсув стану системи в бік майбутньої рівноваги; «шум» при відтворенні цього фенотипу в наступному поколінні означає нове відхилення від рівноваги; черговий акт просіювання знову зсуває образ популяції в напрямку майбутньої норми і т.д. доки кожне нове покоління не стане фенотипічно однорідним і подібним до батьківського. Адаптивно цінна зміна стає стійкою характеристикою системи.\nУсе це показує, що творча роль відбору, як і будь‑якого творчого процесу, в кінцевому підсумку полягає у «запам’ятовуванні випадкового вибору» (Кастлер, 1967), що виражається в даному випадку у виборі однієї з відносно рівноймовірних флуктуацій системи розвитку і перетворенні її у стабільно здійснювану нову норму. Остання протягом усього свого становлення виконує роль «цілі», що визначає напрямок корекції властивостей системи під час її перетворення відбором.\nЦі уявлення кардинально відрізняються від традиційного тлумачення відбору як механізму просіювання і комбінування «спадкових змін», що розуміються як специфічні ефекти певних генів і їх поєднань. При такому підході пояснення успадкованості еволюційних нововведень стає зайвим, оскільки в ній бачать просто іманентну властивість відповідних генів, що не залежить від відбору. Уся процедура «створення» відбором елементарної зміни по суті прирівнюється тут до одноактного вибору її причинного фактора, за яким має слідувати автоматичне відтворення нової ознаки у поколіннях. Навпаки, для епігенетичної теорії стійкість відтворення (успадкованість) — це саме те, що потребує пояснення на основі принципу природного відбору.\nЕтапи елементарного зсуву адаптивної норми, що відображають перебудову видоспецифічної системи розвитку (розмивання старої та стабілізацію нової рівноважної траєкторії і пов’язане з цим перетворення фазового простору системи) спеціально розглянуті вище (рис. 7). Але цей процес може бути описаний і дещо інакше — з точки зору змін, що переживає сукупність індивідуальних онтогенетичних циклів, що належать даній системі (рис. 10).\n \n\nРис. 10. Перетворення індивідуальних модифікаційних спектрів у ході елементарного зсуву адаптивної норми\n1 — співвідношення між зиготами та реалізованими фенотипами; 2 — модифікаційні спектри, що характеризують серію зигот в одному і тому ж інтервалі умов. I — стабільне (еквіфінальне) здійснення групою зигот початкової адаптивної норми А в середніх допустимих для неї умовах N; II — дестабілізований розвиток тих самих зигот в інтервалі екстремальних умов N1, де серед реалізованих відхилень (b), с, d, k, т, z морфоза (b) має адаптивну перевагу; III — стабілізація морфози (b) і перетворення її в адаптивну модифікацію В у рамках нової поліморфної норми АВ; IV — подальша стабілізація фенотипу В на фоні втрати попередньої норми або її збереження у вигляді нестабільної морфози (a). F1 — F2 — Fn— Fx — зміна поколінь; заштриховані здійснені типи розвитку; їх еліміновані варіанти показані низхідними стрілками. F1— F2 — виявлення змінливості; F2 — Fn, Fn — Fx — етапи стабілізуючого відбору"
\n\nРис. 10. Перетворення індивідуальних модифікаційних спектрів у ході елементарного зсуву адаптивної норми\n1 — співвідношення між зиготами та реалізованими фенотипами; 2 — модифікаційні спектри, що характеризують серію зигот в одному і тому ж інтервалі умов. I — стабільне (еквіфінальне) здійснення групою зигот початкової адаптивної норми А в середніх допустимих для неї умовах N; II — дестабілізований розвиток тих самих зигот в інтервалі екстремальних умов N1, де серед реалізованих відхилень (b), с, d, k, т, z морфоза (b) має адаптивну перевагу; III — стабілізація морфози (b) і перетворення її в адаптивну модифікацію В у рамках нової поліморфної норми АВ; IV — подальша стабілізація фенотипу В на фоні втрати попередньої норми або її збереження у вигляді нестабільної морфози (a). F1 — F2 — Fn— Fx — зміна поколінь; заштриховані здійснені типи розвитку; їх еліміновані варіанти показані низхідними стрілками. F1— F2 — виявлення змінливості; F2 — Fn, Fn — Fx — етапи стабілізуючого відбору"
}Оскільки ймовірності здійснення однакових шляхів розвитку для різних зигот одного виду завжди різні через їх генетичні відмінності, то в будь‑якому конкретному діапазоні умов, що виходять за межі звичайних, кожна з них виявляє свій власний спектр реалізованих фенотипічних ухилень (морфозів). З цієї точки зору популяція нормальних зигот може бути представлена як серія різнорідних модифікаційних спектрів, у яких основну частину завжди становить адаптивний фенотип, а граничні відрізки — різні поєднання морфозів (рис. 4; 10, I). В звичайних умовах усі зиготи розвиваються еквіфінально, реалізуючи норму (рис. 10, I). У міру зміни середовища в бік критичного порогу каналізоване розвиток змінюється на розбіжне, тобто виникає все більше морфозів відповідно до специфіки індивідуальних спектрів (рис. 10, II). При збереженні таких умов у ряді поколінь починається відбір на користь найжиттєздатнішого морфозу, що веде до поступового зростання його стійкості і дестабілізації попередньої норми (рис. 10, III). Елімінація інших типів аберантних реакцій спочатку мало ефективна, оскільки вони знову виникають у потомстві відбираного варіанту через його слабку онтогенетичну стійкість. Але у міру стабілізації відбираного фенотипу його успадкування стає все однозначніше, і він (у випадку ксеногамного розмноження) все більше поглинає у схрещуваннях інші аберації, які залишаються нестабільними. Це закріплення адаптивно ланцюгової реакції, що перетворює її в нову норму, призводить до того, що в модифікаційних спектрах послідовних поколінь зигот вона займає все більше місця за рахунок старої норми. Таким чином, обидві норми співіснують на певному етапі в індивідуальних спектрах як дві адаптивні модифікації, реалізовані залежно від коливань умов (рис. 10, III), доки, нарешті, нова повністю не переважає. Можливість реалізації попереднього нормального фенотипу в встановлених нових умовах, якщо не зникає зовсім, то зводиться до рівня аберантної атавістичної варіації (рис. 10, IV).
Отже, кожен елементарний крок у селективному перетворенні нормальної організації зводиться до екстремального розбіжного модифікування (дестабілізації) існуючої норми і подальшого фіксування найоптимальнішого з виявлених варіантів індивідуальної реакції. Модифікування і стабілізація «неперервно кооперуються» (Shmalkhausen, 19686, с. 315). Перша фаза цього кроку означає індивідуалізацію фенотипічного вираження окремих геномів (рис. 10, I—II), друга — його уніфікацію, тобто створення механізму каналізованого розвитку нової адаптації, що нивелює часткові генетичні відмінності (рис. 10, III—IV). Перша з цих фаз — період нестабільності, що розділяє кожні два послідовних стійких стану норми.
Все це дозволяє усвідомити, що, говорячи про рухому функцію відбору, ми маємо на увазі кінцевий результат цього процесу, а не його механізм. Первинне порушення попередньої норми, як ми бачили, взагалі не пов’язане з відбором, а з порушенням умов розвитку, що веде до прояву прихованої змінливості; створення ж нової норми не зводиться лише до вибору оптимального варіанту зміни. Відбір не може безпосередньо зберігати нестабільні реакції, що становлять прихований резерв змінливості. Зсув норми в користь однієї з них здійснюється не за рахунок безпосередньої елімінації інших (бо спочатку вони виникають у поколіннях знову і знову), а за рахунок стабілізації оптимальної реакції, яка стає внаслідок цього все більш універсальною для всієї сукупності розвиваються особин. Таким чином, рухомого відбору як окремого процесу щодо стабілізуючого не існує. Це поняття описує загальний підсумок довгого ряду чергуваних фаз виявлення резерву змінливості і стабілізації певних варіантів змін (Shishkin, 1984a, b). Якщо, за Дарвіном (1952, с. 139), природний відбір — це «збереження корисних індивідуальних змін», то це означає не що інше, а лише набуття ними онтогенетичної стійкості. Зауваження Shmalkhausena (1968b) про нерозривний зв’язок рухомої і стабілізуючої форми відбору (на жаль, залишене неконкретизованим) потребує уточнення. Річ йде не лише про дві сторони, а про два різноякісних і різномасштабних виміри одного процесу.
У тому, що рухомий відбір дійсно оснований на стабілізації часткових аберацій норми, переконує широкий спектр спостережень за ходом її перетворення в різних природних і експериментальних умовах. Найстрогіше підтвердження дають розглянуті вище опити з генетичної асиміляції структурних морфозів (Waddington, 1957), оскільки тут не залишаються сумніви в надзвичайно нестабільному характері вихідного класу реакцій, що формують нову норму. Іншу важливу категорію фактів складають численні експерименти з асиміляції фізіологічних морфозів, тобто примусовим селективним адаптуванням різних груп організмів (зазвичай комах) до нових факторів середовища, наприклад, опити M. M. Kamshilova (1941) щодо виховання холодостійкості у Drosophila, I. V. Kozhanchikova (1941) щодо вирощування жук‑листоїдів на незвичному кормі, подібні опити G. V. Samokhvalova (1951, 1954) і особливо G. X. Shaposhnikova (1961, 1965) щодо зміни господаря у тлі та ін. [Спостережуваний при цьому розвиток нових стійких адаптацій часто приписують «тривалим модифікаціям» у випадках, коли використовуються партеногенетичні клони (наприклад, у ряді опитів з тлями), оскільки їх вважають спадково однорідними. Але різноманітність їх індивідуальної реакції в екстремальних умовах і весь подальший хід перетворення явно вказують на приховану гетерогенність. Остання виникає за рахунок помилок реплікації геному, які неминучі при будь‑якому способі формування генеративних клітин].
Першою реакцією на різку зміну умов, як правило, є висока смертність або зниження плодючості і, безсумнівно, екстремальний фізіологічний стан виживших особин. У ході відбору на стійкість цього стану воно з сублетального перетворюється в одне з звичайних або навіть оптимальне. Цілком очевидно, що ця толерантність до раніше шкідливого фактора виникає тут як нова якість, створена відбором і не існуюча в межах попередньої норми як готова спадкова варіація. Наприклад, у експериментах з охолодженням личинок Drosophila в початковій популяції не було особин, стійко резистентних до холоду або що‑небудь віддавали перевагу помірно зниженій температурі для розвитку (Kamshilov, 1941, 1979), а в експериментах з тлею Dysaphis anthrisci majkopica до 8–11 поколінь виховання на новому господарі не існувало особин, що віддавали перевагу його старому (Shaposhnikov, 1961, 1965). Послідовність змін, спостережуваних у подібних випадках (зокрема, у експериментах з Dysaphis), добре відповідає теоретично очікуваному ходу подій. Спочатку фенотипічно однорідна популяція, що потрапила в незвичні умови, демонструє різке підвищення змінливості, включаючи диференціальну плодючість і життєздатність. Це — фаза виявлення мобілізаційного резерву змінливості (рис. 10, I—II). Далі спостерігається все більша уніфікація фенотипів на користь нової адаптивної норми, причому спочатку ряд поколінь не виявляє переваги старому чи новому варіанту умов. Це — перехідна фаза співіснування обох норм в одних і тих же індивідуальних спектрах як двох адаптивних модифікацій (рис. 10, III). Нарешті, нова норма остаточно стабілізується як єдина адаптація (див. рис. 10, IV). Її відокремлення від носіїв попередньої норми може досягати рівня репродуктивної ізоляції (Shaposhnikov, 1978).
Для описаних перетворень відомо величезна кількість природних аналогій, переважно стосовно випадків «привикання» різних форм комах до пестицидів. Генетична природа виникаючої при цьому резистентності до певного фактора може бути у різних випадках різною (Dubinin, 1966a). У підсумку виникають нові раси, що при схрещуванні з вихідними демонструють різні типи розщеплення (Dobzhansky, 1947). Немає підстав вважати, що процес їх становлення відрізняється від описаного вище і що нова властивість існувала в стабільній формі у одиничних мутантів до початку відбору.
Між тим, як уже зазначалося, саме це припущення становить основу домінуючих уявлень про механізм рухомого відбору. У ньому бачать лише процес накопичення мутацій із спадковим фенотипічним ефектом, тобто вважають, що останній не створюється відбором, а лише підхоплюється ним. Зокрема, появу стійкої до пестицидів раси комах, якщо вона відрізняється від вихідної одним менделевим фактором, розглядають як самоочевидний результат поширення одиничної мутації резистентності (Dobzhansky, 1947, с. 190). Не вдаючись поки до теоретичної сторони цих поглядів, зупинимось на їх співвідношенні з спостережуваними фактами. По‑перше, вони суперечать досвіду експериментальної генетики, що показує, що ефекти «сирих» мутацій, особливо малих (які становлять потенційний матеріал еволюції), нестабільні у порівнянні з адаптивною нормою і в тій чи іншій мірі поглинаються нею. По‑друге, варто звернути увагу на характер використаних доказів. Останні зазвичай зводяться до демонстрації змін частоти певної ознаки в різних популяціях, расах або інших групах, простежуваних або в просторі (варіації типів забарвлення тіла, жилкування крила тощо; Zimmermann, 1933; Тимофеев‑Ресовський та ін., 1965), або навіть у геологічному часі (наприклад, зміни морфотипів зубів у еволюції ссавців; Simpson, 1953). При цьому передбачається, що мінімальні частоти ознаки відповідають її існуванню у вигляді мутацій, а вищі — її входженню до складу норми. Але якщо між мутантним фенотипом і його адаптивним аналогом не передбачається різниці в онтогенетичній стійкості, то ми заздалегідь прирівнюємо акт мутації до створення елементарної адаптації, замість того щоб це довести.
Насправді завдання саме в тому, щоб з’ясувати, якою була фактична успадкованість «мутантної» аберації до початку її експансії і яке взагалі її походження. Чи була вона завжди стійка, чи спочатку представляла лабільну реакцію, поступово стабілізовану і посилену відбором? У більшості випадків ми цього перевірити не можемо, і, говорячи про поширення «мутацій», дослідники мають на увазі складені адаптивні екотипи (як, наприклад, у випадку меланізму у хом’ячків; Gershenson, 1946), у межах яких зазвичай виявляється спадкова різноманітність (Dubinin, 1966a, с. 280). Але там, де передісторія таких форм хоча б частково відома, ми не знаходимо доказів їх первинної стійкості. Прикладом може слугувати еволюція індустріального меланізму у березовій п’ядениці Biston betularia, що, здавалося б, є класичним випадком «включення корисної мутації в норму». Сучасна меланістична морфа цієї метелика в Англії, що домінує у більшості популяцій, переважає над світлою морфою і дає з нею моногенний розщеплення. Проте жодна з цих властивостей не є історично первинною. У більш ранніх меланістів (ймовірно, гібридів), зловлених півтора століття тому, забарвлення було світліше сучасного, тобто домінування було неповним, навіть попри те, що вже тоді темна забарвлення, очевидно, входила до поліморфної норми, даючи адаптивну перевагу на темнолистих деревах (Kettlewell, 1956). Ні домінування, ні правильного розщеплення не виявляються також при схрещуванні англійської темної морфи Biston зі світлою з Канади (де меланістів ще немає); замість цього спостерігається проміжне успадкування (Sheppard, 1970). Домінування — це вираження стійкості фенотипу (Shmalkhausen, 1982, 1968b), і воно, безсумнівно, посилювалося в процесі становлення темної морфи Biston. Екстраполюючи від двох відомих її послідовних станів (в Європі) до початкового моменту її адаптивної історії, ми можемо з великою впевненістю вважати, що вихідним матеріалом для неї слугували слабо виражені ухилення від світлої забарвлення, чутливі до коливань зовнішніх факторів і генетичної конституції особин, тобто однотипні морфози норми, які відбір стабілізував і посилював крок за кроком. Потенційна здатність до такої меланістичної реакції взагалі широко розповсюджена у метеликів (Dubinin, 1966a), і її можна викликати штучно, наприклад, охолодженням гусениць (Standfuss, 1902). Очевидно, що немає підстав приписувати властивості сучасних нормальних меланістів елементарним мутаціям зі стійким ефектом, що виникли у їх предків.
Ще одне загальне джерело передбачуваних доказів появи стійких адаптацій шляхом одиничних генетичних змін складають біохімічні мутації у нижчих організмів, зокрема у бактерій. При пересіванні тієї чи іншої культури на неповноцінне середовище (наприклад, штаму кишкової палички, що ферментує галактозу, на середовище з лактозою) виживають лише окремі мутантні клітини, причому методом реплік можна показати, що адаптивно цінна зміна не індукована новими умовами, а існувала в початковій культурі (Dubinin, 1966a, 1976). Це виглядає як спонтанне появлення нової адаптації. Але легко побачити, що тут відбирається не стійке пристосування, а просто одна з елементарних модифікацій. Так, у нашому прикладі здатність ферментувати лактозу проявляється у мутантних клітин (у тій чи іншій мірі) лише на відповідному провокаційному фоні, тоді як на вихідному галактозному середовищі вони виконують свою попередню нормальну функцію; інакше такі клітини загинули б до пересіву. У модифікаційних спектрах мутантів даного типу обидві реакції присутні одночасно, і вибір між ними визначається лише зовнішніми умовами. Стійке ж закріплення нової реакції, очевидно, може відбуватися лише в ході розмноження зміненого штаму в новому середовищі по мірі відбору серед багатьох індивідуальних клітин. Остання можливість допускається у багатьох випадках, коли ясно видна поступовість пристосування бактерій до зростаючих доз шкідливого фактора (Dubinin, 1976). Проте, схоже, не завжди усвідомлюється, що в цьому і полягає загальна закономірність становлення нових адаптацій. Потрібно враховувати, що при високій швидкості розмноження бактерій стабілізація відобраних морфозів у новому середовищі відбувається надзвичайно швидко, затемнюючи їх первинну нестабільність.
Отже, уявлення про еволюцію як відбір фенотипічно стабільних геноваріацій будується на фактах, які не можуть його довести. Ці факти характеризують не процес адаптаціогенезу як такового, а або його вихідну основу (прояви біохімічних мутацій), або кінцевий результат, тобто зміни частот вже складених компонентів поліморфних систем. На практиці ж становлення нових пристосувань неможливе без стабілізації індивідуальних реакцій, що дають їм початок.
Але якщо матеріал еволюції складають лабільні реакції (модифікації), що здійснюються нормо‑утворюючими генотипами лише в ухильних умовах, то ясно, що вони, за визначенням, не підкоряються менделевим правилам. Схрещування двох елементарних морфозів норми не може дати стійкого результату, незважаючи на генетичні відмінності їх носіїв. У випадку ж схрещування такого морфозу з стійким нормальним фенотипом перший, природно, поглинається другим (при розвитку потомства в нормальних умовах). Єдина природна ситуація, де правильне менделеве розщеплення має місце (якщо виключити найгрубіші мутаційні порушення), — це схрещування варіантів норми, тобто стабілізованих фенотипів, що складають поліморфну систему. Лише в цьому випадку в епігенетичних системах гибридів виникають альтернативні шляхи стійкого розвитку (креоди), між якими здійснюється упорядкований вибір. Коротко кажучи, менделюють лише продукти каналізованого розвитку (адаптивні морфи), а не продукти його дестабілізації (морфози). Тому для епігенетичної теорії еволюції світ природної змінливості як цілісність не описується менделівськими термінами. Його складають не алелі (у яких теорія бачить лише відношення між певними типами розвитку), а лише самі фенотипи, тобто варіанти норми та їх аберації. Поведінка окремих фенотипів у схрещуваннях залежить виключно від їх приналежності до цих двох класів, що характеризують тип індивідуального розвитку, а також від самих умов розвитку. Дарвінівська невизначена змінливість — це сукупність аберацій норми і, отже, область фенотипів, що не мають упорядкованого успадкування. Різниця, описувана в менделівських алелях, — це результат відбору за альтернативними фенотипами, а не просто будь‑яке проявлення генетичної різниці.
З цих позицій корінним чином змінюється пояснення причин, чому корисні елементарні зміни не розчиняються у схрещуваннях і можуть зберігатися відбором. У історії критики дарвінізму припущення про таке розчинення відоме як «кошмар Дженкіна», і протягом усього нашого століття еволюціоністи не втомлюються повторювати, що цей аргумент усунуто відкриттям менделевих законів з їх принципом «чистоти гамет». Але ці закони універсальні для хромосом, а не для ознак і не застосовні до невизначеної фенотипічної змінливості. Справжня причина дієвості відбору полягає не в високій успадкованості і строгій дискретності його елементарних змін, а в приналежності їх до обмеженого простору аберацій, властивих даній фенотипічній нормі. У результаті ці мало стійкі самі по собі варіації сумарно повторюються у поколіннях, забезпечуючи стабільність потенційного субстрату відбору без стабільності індивідуального успадкування. Відбір на користь однієї з них веде не до збереження її «незмішуваних факторів», а лише до збільшення чисельної ролі гамет, залишених її носіями незалежно від їх індивідуальної генетичної конституції, що змінюється в кожному поколінні.{
"translated_text": "На початкових етапах процесу ці гамети здатні відтворювати відбираний фенотип лише серед інших варіантів, що складають початкову гамму аберацій; проте кожного разу їх комбінація підвищує ймовірність його подальшого здійснення. У міру продовження відбору на стійкість самовідтворення цього фенотипу (тобто в результаті переважного збереження в кожному поколінні тих його нащадків, які змогли його повторити) поступово зростає його індивідуальна успадкованість. Цей процес базується на безперервному перебудовуванні генотипу гамет у підтримуваній відбором лінії.\nДосягнута таким шляхом висока успадкованість фенотипу, заснована на його стабілізації, проявляється тепер у його переважаючій домінації при схрещуваннях з різними випадковими варіаціями, чим і демонструється його «нерозчинність». Якщо ж подібним чином із початкового спектру відбираються одразу кілька фенотипічних варіантів, то в послідовних поколіннях їх гібридизація буде приводити до все більш упорядкованого їх успадкування у визначених співвідношеннях. Таким чином, менделєвське розщеплення за фенотипами — це не причина збереження еволюційних новацій, а результат їх стабілізації в ході відбору.\n* * *\nПодані уявлення про епігенетичний механізм еволюційного процесу дозволяють оцінити сутність понять, що лежать в основі синтетичної теорії еволюції. Аналіз останньої ускладнюється тим, що з часом все більше зростає розрив між її фактичними уявленнями і тими, які приймаються декларативно, без використання в її побудовах. Очевидно, що предметом розгляду має бути саме перша категорія понять.\nВихідним тут є положення, що субстратом відбору є мозаїчні менделєвські алельні фактори, ототожнювані з хромосомними локусами. Еволюція — це зміна генетичного складу популяції, і її швидкість визначається швидкістю заміщення алелей (Dobzhansky, 1947; Грант, 1980). Для хромосомних генів передбачається упорядковане фенотипічне вираження, оцінюване кількісно як відносний внесок у пристосованість. Стійкість фенотипів або ознак розглядається чисто преформістськи як властивість контролюючих їх генів, і тому її пояснення не входить до завдань теорії. Поняття стійкості має тут лише надіндивідуальний сенс і виражає або фіксацію певних частот алелей (генетичний рівноваг), або, у більш прямому значенні, гомозиготизацію за тим чи іншим з них, тобто досягнення алелем 100%-ної частоти в популяції. Останнє відбувається за рахунок усунення альтернативних алелей — в результаті відбору або ж генетичного дрейфу (при обмеженні розмірів популяції), коли випадкові відхилення частот від рівноваги можуть набувати незворотного характеру. Відповідно предметом розгляду теорії є не організація генотипу та керованого ним індивідуального розвитку, а організація генофонду популяції.\nПоняття генетичної обумовленості та успадкованості (стійкості) ототожнюються розгляданою теорією, тобто будь-яка генетична зміна розуміється як фенотипічно успадкована, хоча і визнається, що її ефект може спотворюватися генотипічними та зовнішніми впливами. Тому успадковані зміни розглядаються як не залежні від природного відбору; останній лише оперує ними, а не створює їх. Ненаслідкові, або модифікаційні, відхилення розуміються як шуми, що затемнюють дію генів і гальмують відбір генетичних змін (Дубінін, 1966а; Майр, 1968; Грант, 1980). Оскільки між останніми, за визначенням, не визнається різниці у ступені стійкості їх ефекту, то поняття фенотипічної норми і аберації позбавляються в теорії будь‑якої функціональної ролі. Для них немає інших характеристик, окрім частот відповідних фенотипів і генів у популяції. З тієї ж причини уявлення про еволюційне поглинання одних фенотипів іншими через систему схрещувань є тут цілком беззмістовним; мова може йти лише про їх заміну шляхом витіснення, що виражає заміну відповідних алелей. Це заміщення означає зменшення генетичної дисперсії популяції за пристосованістю і, відповідно, зменшення генетичного навантаження. Чим більше число генів одночасно зачіпається відбором, тим більша величина навантаження і тим повільніше мають відбуватись зміни (за умови збереження достатньої чисельності популяції).\nОскільки будь‑яка мутація розглядається теорією як алельна зміна, що контролює стійку ознаку, це означає фактичне визнання стрибкоподібної еволюції, хоча на словах її часто заперечують (Майр, 1968; Р’юз, 1977; тощо). Обмеження, накладені на сальтаціонізм, мають тут технічний, а не принциповий характер. Там, де між морфами або расами встановлюється просте алельне різниця, це вважається вказівкою на виникнення однієї форми з іншої шляхом одиничної мутації (Dobzhansky, 1947, с. 50, 190; Грант, 1980, с. 88, 174); там же, де передбачається множина таких різниць (наприклад, між послідовними видами), еволюційне перетворення розглядається як послідовність мутацій (Dobzhansky, 1947, с. 52). Саме мутація виступає тут як творча сила, тоді як творча роль відбору зводиться лише до гомозиготизації рецесивних генів, що виявляє нові фенотипи, або до комбінування генних ефектів, або, нарешті, до створення оптимального генотипічного середовища для вираження гена. Еволюція розглядається тут як діалог між середовищем і генами (Новинський, 1978).\nЦі редукціоністські принципи теорії, зокрема відбір генів і наділення їх коефіцієнтами пристосованості, все менше задовольняють її прихильників і вже давно характеризуються як «надмірні спрощення» (Dobzhansky, 1947, с. 106). Але звідти не випливає великої ясності у питанні, що ж у такому випадку становить справжній субстрат відбору. Як субстрат вказуються генотипи і генетичні комбінації (Шеппард, 1970), або лише фенотипи (Майр, 1968), або все це разом (Грант, 1980). Усе це скоріше відображає намір відійти від старих уявлень, ніж справжнє відходження від них. Тому не дивно, що механізм рухомого відбору досі зводять до заміщення одного алеля іншим (Грант, 1980, с. 144). За визнанням R. Lewontin (Lewontin, 1970), сучасна популяційна генетика, на якій базується синтетична теорія, все ще виходить з уявлення про чисто мозаїчне комбінування генів, навіть не враховуючи їх організацію в групи зв’язку.\nНайбільш ясним вказанням на збереження редукціоністського розуміння суті відбору є характер уявлень про результат його дії, притаманний синтетичній теорії. Будь‑який ефективний відбір має приводити до гомогенізації свого субстрату, і, передбачаючи відбір алелей, ми, природно, маємо очікувати (у межі) їх переходу у гомозиготний стан. Але саме це і передбачається синтетичною теорією до сьогоднішнього дня (Шеппард, 1970; Айала, 1981; Солбриг, Солбриг, 1982). Відповідність реальної картини цим очікуванням (гетерогенність природних популяцій) вважається вказівкою на наявність факторів, що діють, «незважаючи на сили відбору» (Айала, 1981, с. 51). Навіть коли робляться спроби пояснити гетерогенність відбором комплексів модифікаторів, що вирівнюють пристосованість для максимуму варіантів генетичного фону (Майр, 1974), мова все одно йде про збереження суми генів, відповідальної за цей ефект.\nКосвенною ознакою такого розуміння відбору є і сам факт появи нейтральної гіпотези еволюції (Kimura, 1968), висунутої як альтернатива синтетичній теорії. Одним із її підстав є, як відомо, наявність природного поліморфізму за ізоферментами, що вказує на множинність відповідних алелей, яка розглядається тут як щось, що не узгоджується з ідеєю відбору. Іншими словами, завдяки синтетичній теорії дарвінівський відбір настільки міцно ототожнився в умах еволюціоністів з відбором алелей, що відсутність останнього виглядає для них рівнозначною відсутності відбору взагалі і змушує їх говорити про «недарвінську» еволюцію. Це яскраво показує, що, незважаючи на формальне визнання синтетичною теорією відбору за фенотипами, «її ідеали залишились прежніми» (Новинський, 1978).\nТим же незмінним залишається для синтетичної теорії і визнання фіксованих співвідношень між геном і ознакою (формально відкинутих хромосомною генетикою). Підкоряти природну змінність менделєвським правилам — це і означає визнати, що закони мейотичної рекомбінації хромосомних локусів мають однозначне вираження у змінах фенотипів — хоча б воно і затемнювалося зовнішнім або генотипічним середовищем. Далі, оскільки тиск відбору вимірюється виключно за зміною частоти форм у поколіннях (Шеппард, 1970), очевидно, що всі судження про зміну генетичного складу на цій основі можливі лише за наявності чіткого відповідності між генами і фенотипами. Точно так само і уявлення про закріплення алелей (шляхом відбору або дрейфу) як джерела еволюційних новацій передбачає ту ж саму зв’язок.\nЗ точки зору епігенетичної концепції вихідним пунктом усіх цих побудов є зведення цілісних властивостей організму, створених природним відбором, до первинних характеристик геномних одиниць, що існують незалежно від відбору. Це виражається не лише у наданні генам внеску в пристосованість і стійкого фенотипічного ефекту, а передусім у самому розумінні гена. Менделєвський фактор, який об’єктивно виражає різницю двох створених відбором варіантів організації генотипу (що контролюють відповідні креоди), ототожнюється тут з ділянкою хромосоми, тобто різниця у властивостях двох систем переноситься на їх елементи. Локус, що визначає перемикання креодів у даній системі схрещувань, перетворюється у носія незмінних характеристик, збережених у ньому у всіх рекомбінаціях. Відповідно мутація локуса перетворюється в механізм створення менделєвської різниці, тобто результат історичного процесу замінюється одноактною подією. Це саме та логіка, яка змушує багатьох еволюціоністів‑ембріологів бачити у розбіжності типів онтогенезу у різних групах організмів (онтогенетичної дивергенції Бера) доказ їх стрибкоподібного походження один від одного шляхом одноактної девіації (Шишкін, 1981). Для епігенетичної концепції реальний природний світ менделєвських співвідношень — це лише сукупність стабілізованих фенотипів, що характеризують поліморфну систему виду і об’єднана вільним схрещуванням. З цієї точки зору модель еволюційних подій, прийнята синтетичною теорією, виходить з перенесення на невизначену змінність властивостей адаптивної поліморфної системи, тобто сирого матеріалу еволюції, який замінюється її продуктами — мозаїкою стабілізованих форм. Відбір тут «вводиться в дію» в умовах, коли його робота вже виконана. Уявленню про зміну частот алелей відповідає в природних популяціях лише одна реальність — зміна концентрацій стійких морф, що у схрещуваннях дають менделєвські розщеплення.\nТому виходити з конкуренції алелей як двигуна еволюції — означає оперувати лише адаптивними нормами, тоді як невизначені аберації, викликані порушеннями розвитку і що становлять справжній субстрат відбору, у цьому випадку взагалі усуваються з розгляду. Тільки за такого обмеження матеріалу схрещування варіантами норми або штучно стабілізованими формами (чисті лінії) має місце ситуація, коли фенотипи не поглинають один одного, а лише змінюють частоти у відповідь на короткочасну дію відбору. І якщо прийняти, що стійкі різниці цих фенотипів, виражені в менделєвських факторах, створюються мутаціями, а не відбором, то на долю останнього не залишиться нічого, окрім поширення або знищення фенотипічних результатів мутації. У такому випадку дія відбору дійсно не повинна принципово відрізнятись від результатів випадкових частотних флюктуацій (дрейфу) алельних фенотипів, і це пояснює причини самого виникнення ідеї дрейфу.\nОтже, при порівнянні двох викладених систем поглядів на природу еволюційного процесу виявляється їх діаметральна різниця. Якщо для синтетичної теорії відбір — це перерозподіл алельних факторів, що виникають поза ним і розуміються як стани хромосомних локусів, то для епігенетичної теорії результатом відбору, використовуючи ту ж мову, є створення і розмивання алельних властивостей фенотипів у процесі зміни адаптивних норм. Іншими словами, алелі створюються і знищуються відбором. Якщо, наприклад, два фенотипи, описані менделєвським аналізом як гомозиготи АА і аа, у схрещуваннях дають третій фенотип Аа, що має селективну перевагу, то для синтетичної теорії його фіксація в природі не означає нічого більшого, ніж встановлення балансу алелей, при якому чисельність таких гібридних форм врівноважується елімінацією менш пристосованих гомозигот. Навпаки, для епігенетичної теорії відбір на користь фенотипу Аа означає його стабілізацію щодо двох інших, тобто таку перебудову його генотипу і системи розвитку, в результаті чого він все ефективніше поглинає у схрещуваннях вихідні варіанти, а отже, і перестає закономірно розщеплюватись на форми АА і аа. Разом із каналізацією його розвитку і розмиванням (у нащадків) альтернативних креодів, що відповідають початковим чистим лініям, втрачається і сама можливість інтерпретації цього фенотипу в термінах алелей А і а. Якщо шукати аналогії цим поглядам у генетичному мисленні, можна сказати, що ідея «відбору на користь фенотипу кращого з гомозигот», прийнята фішерівською гіпотезою еволюції домінантності (Fisher, 1930), розширюється тут у уявлення про відбір на домінування кращого фенотипу взагалі.\nПідсумкове порівняння двох еволюційних концепцій приводить до висновку, що перша з них (епігенетична) по суті значно більш «генетична», ніж та, з якою пов’язують це визначення. Такі фундаментальні емпіричні узагальнення генетики, як стійкість норми («дикого типу») у порівнянні з абераціями і здатність до їх поглинання; порушення менделєвської успадкованості, виражені в поняттях експресивності і пенетрантності; гетерогенність будь‑яких класів фенотипів; залежність ознак від генотипу в цілому; вплив відбору на їх домінування — все це, як ми бачили, належить до основоположних понять епігенетичної теорії, але не знаходить вираження в мові протилежної концепції. Синтетична теорія виходить на практиці не стільки з реальних досягнень хромосомної генетики, скільки з абстракцій раннього менделізму, що спиралося на однозначну відповідність генів і ознак і ототожнення еволюційних новацій із заміною генів.\nЦі висновки сьогодні вже не здаються несподіваними і знаходять своє підтвердження у все посилюваній тенденції до критичної переоцінки основ синтетичної теорії. Генетики все частіше характеризують її лише як «корисну абстракцію» (Камшилов, 1979, с. 141), що являє собою «тимчасове і спрощене» будування (Солбриг, Солбриг, 1982, с. 78); бачити в ній дійсний синтез генетики і дарвінізму — означає «видавати бажане за дійсність» (Бабков, 1981, с. 413). Визначення еволюції як зміни генетичного складу популяції нині розглядається як редукціоністське (Майр, 1981); у основі зміни ознак пропонується бачити перебудову генотипів, а не просто заміну генів (Майр, 1968). До числа примусових спрощень теорії еволюціоністи‑генетики відносять сьогодні всі головні її принципи — відбір алелей, оцінку їх за пристосованістю та однозначність відповідності генотипа і фенотипа; аналіз генетичних змін за фенотипичним складом вважається довільним і не враховує закономірностей індивідуального розвитку (Солбриг, Солбриг, 1982, с. 77, 78, 154, 263). Використовувані теорією моделі дії відбору на частоти локуса з двома алелями вважаються «повністю нереалістичними» (Там же, с. 258). Зростає і скептицизм щодо ролі генетичного дрейфу як фактора еволюції (Шеппард, 1970; Солбриг, Солбриг, 1982). Навпаки, раніше відмова від еволюційної ролі модифікаційних змін замінюється тепер її формальним визнанням (Майр, 1968, 1974), хоча, як справедливо зазначалося раніше, це рівносильно «перегляду основних положень генетичної концепції еволюції популяцій» (Дубінін, 1966а, с. 373). Ще одним свідченням відходу теорії від своїх фундаментальних принципів є її прагнення відмовитися від ідеї генетичного навантаження (Майр, 1974; Грант, 1980; Мейнард‑Смiт, 1981; Галл, 1980), викликане труднощами поєднання її з фактами генетичного поліморфізму всередині природних популяцій. Але разом з ідеєю навантаження неминуче усувається і єдине, запропоноване теорією пояснення механізму рухомого відбору як зменшення дисперсії генетичного складу популяції за пристосованістю. Поняття відбору на користь кращих алелей таким чином втрачає зміст. Усі ці тенденції об’єктивно відображають одне й те саме — невідповідність мови синтетичної теорії уявленням про цілісний організм як предмет природного відбору. При очевидній відсутності упорядкованих (преформованих) співвідношень між геномними одиницями та властивостями організму результати відбору за фенотипами принципово не можуть описуватись у термінах генних частот. Одним із косвених і незалежних визнань цього факту є думка про заміну генів хромосомними континуумами як майбутньої основи побудов популяційної генетики (Lewontin, 1970).\nЧим більше висувається поправок, спрямованих на модернізацію синтетичної теорії, тим важче визначити, що ж становить її сутність. Теорія не може залишатись самою собою, відмовляючись від власних основ. Складається враження, що для неї насправді немає іншого логічного фундаменту, окрім послідовного преформістського редукціонізму, що пов’язує гени з ознаками, і що поза ним вона перетворюється на еклектичне будування."
}Очевидно, саме тому, незважаючи на всі поправки, вона залишається для критиків концепцією, що абстрагується від проблеми цілісності організму і його індивідуального розвитку, і в якості такої вважається неприйнятною для пояснення еволюції, принаймні на рівні її макрохарактеристик (Bertalanffy, 1969; Но, Saunders, 1979; Alberch, 1980, 1982; Wake et al., 1983; Maderson et al., 1982; Блюменфельд, 1974; Новинський, 1978).
Очевидна неможливість поєднати дарвінівське уявлення про природний відбір із спрощеннями мутаціонізму та раннього менделізму викликає не лише прагнення обмежити роль цих спрощень шляхом перегляду синтетичної теорії. Поряд з цим спостерігаються протилежні спроби — вирішити виникаюче протиріччя за рахунок дарвінізму. Так, М. Д. Голубовський (1981), майже повторюючи слова Йогансена про усунення генетикою основ дарвінівської теорії (Філіпченко, 1977, с. 191), вважає, що кожен крок у розвитку вчення про спадковість означав послідовне обмеження постулатів селекціонізму. За його думкою, немає дарвінівської невизначеної змінливості — її замінила сувора упорядкованість розщеплень, обмежена менделєвськими правилами і не залежна від відбору. Немає творчої ролі відбору, бо він може зберігати лише те, що створюється мутаціями (як один із прикладів наведено індустріальний меланізм у Biston betularia). Тому немає і поступовості еволюційних змін — її місце мають зайняти мутаційні стрибки, оцінювані відбором. Це поклик повернутися до уявлень де Фріза і Йогансена не здається випадковим; він відображає, хоча й у своєрідній формі, чітке розуміння ілюзорності того, що сьогодні називають синтезом дарвінізму і генетики.
Справжня асиміляція досягнень генетики дарвінівським вченням не може будуватись на абсолютизації виявлених нею закономірностей (як і взагалі будь‑яких форм біологічної упорядкованості) у якості універсальних факторів, що діють поза зв’язком з природним відбором. Навпаки, їх слід трактувати як результати відбору, що не існують поза ним. Завданням еволюційної теорії є не лише викриття біологічних законів, а й визначення умов їх виконання, тобто усвідомлення їх відносності. Тому, говорячи про еволюційне пояснення даних генетики, ми повинні мати на увазі весь реальний спектр накопичених нею фактів, а не лише суму правил, що використовуються для гибридологічного аналізу чистих ліній. Єдиною прийнятною основою такого пояснення виглядає епігенетична теорія Шмальгаузена‑Уоддингтона, що розглядає спадковість як вираження стійкості індивідуального розвитку, створеного природним відбором. Можна стверджувати, що саме з цією теорією, що ставить у центр уваги властивості цілісної системи розвитку організму і прагне до опису фактів ембріології і генетики однією мовою, пов’язані перспективи майбутнього розвитку дарвінізму.
ЛІТЕРАТУРА
Агаєв М. Г. Експериментальна еволюція. Л.: Вид-во ЛГУ, 1978.
Айала Ф. Механізми еволюції // Еволюція. М.: Мир, 1981. с. 33–65.
Астауров Б. Л. Генетика статі // Актуальні питання сучасної генетики. М.: Вид-во МГУ, 1966. С. 65–113.
Астауров Б. Л. Homo sapiens et humanus — людина з великої літери // Новий світ. 1971. № 10. С. 214–229.
Бабков В. В. Системний стиль у вивченні природного відбору // Системні дослідження. М.: Наука, 1981. С. 404–419.
Балкашина Є. І. Випадок спадкового гомеозису. Геноваріація aristopedia у Drosophila melanogaster // Журн. експеримент. біології. 1928. Т. 4. вип. 2.
Балкашина Є. І., Ромашов Д. Д. Генетичний склад популяцій Drosophila // Біол. журн. 1935. Т. 4, № 1. С. 81–106.
Бауер Е. С. Теоретична біологія. М.; Л.: ВІЕМ, 1935. 205 с.
Белоусов Л. В. Джерела, розвиток і перспективи теорії біологічного поля // Фізичні та хімічні основи життєвих явищ. М.: Вид-во АН СССР, 1963. С. 59–117.
Белоусов Л. В. Цілісні та структурно‑динамічні підходи до онтогенезу // Журн. общ. біології. 1979. Т. 40, № 4. С. 514–529.
Белоусов Л. В., Чернавський Д. С. Нестабільність і стабільність у біологічному морфогенезі // Онтогенез. 1977. Т. 8, № 2. С. 99–114.
Блюменфельд Л. А. Фізичні аспекти біологічної еволюції // Філософія в сучасному світі. Філософія і теорія еволюції. М.: Наука, 1974. С. 56–71.
Бляхер Л. Я., Воронцова М. А., Ліознер Л. Д. Каузально‑аналітичний метод у вченні про індивідуальний розвиток // Тр. Ін‑та експеримент. морфогенезу. 1935. Т. 3. С. 223–239.
Волькенштейн М. В. Фізичний сенс нейтральної гіпотези еволюції // Журн. общ. біології. 1981а. Т. 42, № 5. С. 680–686.
Волькенштейн М. В. Біофізика. М.: Наука, 1981б. 575 с.
Галл Я. М. І. І. Шмальгаузен і проблема факторів еволюції // Історико‑біологічні дослідження. М.: Наука. 1980. Вип. 8. С. 106–123.
Гершенсон С. М. "Мобілізаційний резерв" внутрішньовидової змінливості // Журн. общ. біол. 1941. Т. 2, № 1. С. 85–107.
... (далі список літератури без змін){"translated_text":"Biological stability // Towards a Theoret. Biology / Ed. C. Waddington. Chicago: Aldine, 1970. P. 1–17.\nGurwitsch A. Ueber Determination, Normierung und Zufall in der Ontogenese // Roux' Arch. 1910. Bd. 30. S. 133–193.\nGurwitsch A. Die Vererbung als Verwirklichungsvorgang // Biol. Zentr.-BI. 1912. Bd. 32. S. 458–486.\nGurwitsch A. Ueber den Begriff des embryonalen Eeldes // Roux' Arch. 1922. Bd. 51, h. 3/4. S. 383–415.\nHaldane S. The Biochemistry of Genetics. L.: Alien and Unwin, 1954. 144 p.\nНо М. W., Sounders Р. Т. Beyond neo-Darvinism — an epigenetic approach to evolution // J: Theor. Biol. 1979. Vol. 78, N 4. P. 573–591.\nJohannsen W. Elemente der exakten Erblichkeitslehre. Aufl. 3. Jena: Fischer, 1926. 736 S.\nKettlewell H. B. D. A. resume on investigations on the evolution of melanism in the Lepidoptera // Proc. Roy. Soc. London B. 1956. Vol. 145, N 920. P. 297–303.\nKimura М. Genetic variability maintained in a finite population due to mutational production of neutraly or nearly neutral isoalleles // Genet. Res. 1968. Vol. 2. P. 247–269.\nKojima K. 1. Is there a constant fitness value for a given genotype? // Evolution. 1971. Vol. 25. P. 281–285.\nLewontin R. C. On the irrelevance of genes // Towards a Theoretical Biology // Ed. C. Waddington. Chicago: Aldine, 1970. P. 67–88.\nMaderson P. F. A. et al. The role of development in macroevolutionary change. Croup report // Life Sci. Res. Rep. 1982. N 22- P. 279–312.\nRoux W. Gesammelte Abhandlungen uber Entwicklungsmechanik der Organismen. Leipzig: Engelmann, 1895. Bd. 1. 816 S.; Bd. 2. 1075 S.\nSimpson G. G. Major Features of Evolution. N. Y.: Columbia Univ. press. 1953. 434 p.\nStandfuss М. Zur Frage des Gestaltung und Vererbung auf Grund 28-jahriger Experimente // Insektenborse. 1902. Bd. 19. S. 155–169.\nWaddington C. H. Genetic assimilation of an acquired characters // Evolution. 1953. Vol. 7. P. 118–126.\nWaddington С. Н. The Strategy of the Genes: a Discussion on Some Aspects of Theoretical Biology. L.: Alien and Unwin, 1957. 262 p.\nWaddington С. Н. Principles of Development and Differentiation. N. Y.: Macmillan, 1976. 115 p.\nWake D. В., Roth G., Wake M. Н. On the problem of stasis in organismat evolution // J. Theor. Biol. 1983. Vol. 101. P. 211–224.\nWolpert L. Mechanisms of limb development and malformation // Brit. Med. Bull. 1976. Vol. 32, N 1. P. 65–70.\nWoodger I. Н. What do we mean by «inborn»? // Brit. J. Philos. Sci. 1953. Vol. 3. P. 319.\nWright S. An intensive study of the inheritance of color // Carnegie Inst. Wash. Publ. 1916. Vol. 241, pt. 2. P. 59–160.\nWright S. Evolution organic// Encyclopaedia Britanica. L., 1964. Vol. 8. P. 915–931.\nZimmermann K. Ober Mutationen in wilden Populationen // Mitt. Zool. Mus. Berlin. 1933. Bd. 19. S. 439–452."}