
Vegerina ta іn. (2013) Vyznachennya spivvidnoshennya diploїdiv i triploїdiv sered metamorfiv zelenyh zhab
Вегерина А. О., Мелешко Е. В., Пирина І. С., Сапожникова В. А., Бирюк О. В. Визначення співвідношення диплоїдів і триплоїдів серед метаморфів зелених жаб у Північно-Донецькому центрі різноманіття Pelophylax esculentum complex // Вісник Харківськог...
Вегерина А. О., Мелешко Е. В., Пирина І. С., Сапожникова В. А., Бірюк О. В. Визначення співвідношення диплоїдів і триплоїдів серед метаморфів зелених жаб у Північно-Донецькому центрі різноманіття Pelophylax esculentus complex // Вісник Харківського національного університету імені В. Н. Каразіна. Серія: біологія. – 2013. – Вип. 18, № 1079. – С. 107–113.
УДК: 597.851
Визначення співвідношення диплоїдів і триплоїдів серед метаморфів зелених жаб у Північно-Донецькому центрі різноманіття Pelophylax esculentus complex
А. О. Вегеріна, Е. В. Мелешко, І. С. Пиріна, В. А. Сапожникова, О. В. Бірюк
Харківський національний університет імені В. Н. Каразіна (Харків, Україна)
mykhailova.o.v@gmail.com
Перевірено гіпотезу, що частка триплоїдів серед метаморфів представників Pelophylax esculentus complex у Північно-Донецькому центрі різноманіття зелених жаб є вищою, ніж серед статевозрілих особин. Визначено плоїдність 30 метаморфів, зібраних у липні 2013 р. Встановлено, що значення середньої довжини еритроцитів у метаморфів нижче такої у статевозрілих жаб. Плоїдність визначалась шляхом підрахунку кількості хромосом у метафазних пластинках, отриманих з ендотелію кишечника. Для визначення плоїдності у разі відсутності ділячих клітин застосували Ag‑фарбування, яке дозволяє виявити ядерця в інтерфазних ядрах. Усі вивчені особини виявилися диплоїдними. З урахуванням вибірки, що була вивчена у 2012 р., один триплоїд був зареєстрований в об’єднаній вибірці з 47 молодих особин; гіпотеза, що перевірялася, не підтвердилася. Висунуто припущення про причини низької частки триплоїдів у дослідженій вибірці.
Ключові слова: Pelophylax esculentus, триплоїди, еритроцити, Ag‑фарбування, плоідність.
The determination of the ratio of diploid and triploid green frogs underyearlings from the Pelophylax esculentus complex Seversky Donets diversity center
A. O. Vegerina, O. V. Meleshko, I. S. Pyrina, V. O. Sapoznikova, O. V. Biryuk
The hypothesis has been tested that the percentage of triploids among the metamorphs of Pelophylax esculentus complex representatives in the Seversky Donets green frogs diversity center is higher than that among the mature individuals. The ploidy of 30 metamorphs collected in July, 2013 was defined. It was found that the metamorphs erythrocytes mean length is lower than that in the mature frogs. Ploidy was defined by counting chromosomes in metaphase plates derived from intestine endothelium. To determine the ploidy in the absence of dividing cells the Ag‑staining was used, which reveals the nucleoli in interphase nuclei. All the studied specimens were diploid. Given the sample studied in 2012, one triploid was registered in the combined sample of 47 young individuals; the tested hypothesis was not confirmed. The assumptions about the causes of the low percentage of triploids in the studied sample were made.
Key words: Pelophylax esculentus, triploids, red blood cells, Ag‑staining, ploidy.
Вступ
У склад гибридного комплексу зелених жаб (Pelophylax esculentus complex) входять два батьківські види: ставочна жаба Pelophylax lessonae (Camerano, 1882) і озерна жаба Pelophylax ridibundus (Pallas, 1771), від схрещування яких утворюються міжвидові гібриди. Для позначення цих гібридів (з певною умовністю) використовується назва, аналогічна видовій — їстівна жаба Pelophylax esculentus (Linnaeus, 1758). Різні форми гібридів відрізняються плоїдністю та характером їх гаметоогенезу; у типічному випадку вони геміклонально (без рекомбінації) передають у гамети один з батьківських геномів (Plötner, 2005). Особи батьківських видів живуть і розмножуються з гібридами спільно, утворюючи геміклональні популяційні системи. Розташований у басейні р. Северський Донец регіон, що характеризується високим різноманіттям гібридних зелених жаб, у тому числі поширенням триплоїдних гібридів, був названий Північно-Донецьким центром різноманіття P. esculentus complex (Шабанов та ін., 2009).
У першій роботі, в якій було повідомлено про виявлення триплоїдних гібридів зелених жаб у цьому районі, зареєстрована частка триплоїдів серед гібридів була відносно високою і становила 30 % (Borkin et al., 2004). Досліджена у цій роботі вибірка зелених жаб включала значну кількість сеголетків. У ряді інших робіт, де досліджували статевозрілих особин, реєструвалася нижча частка триплоїдів у загальній кількості гібридів: 9 % (Mezhzherin et al., 2010) або 11 % (Козак та ін., 2012).
Описана різниця в результатах дозволяє висунути гіпотезу про те, що частка триплоїдів серед P. esculentus у Північно-Донецькому центрі різноманіття зелених жаб зменшується з віком. На користь цієї гіпотези свідчить те, що триплоїдні гібриди не є окремим таксоном і, ймовірно, не мають самостійної, незалежної від диплоїдів генеалогії. Геноми таких жаб є результатом еволюції диплоїдних представників батьківських видів, і їх триплоїдність можна розглядати як цитогенетичну аномалію. Спираючись на це, можна припустити, що відбір зменшує частку триплоїдів у старших вікових класах порівняно з молодшими.
У 2012 р. частина авторів даної роботи досліджувала вибірку з 17 цьоголіток і нестатевозрілих особин, зібраних в околицях с. Гайдари Зміївського району Харківської області. Плоїдність жаб у цій роботі визначали за допомогою методу пресованих препаратів (Михайлова та ін., 2011). Зареєстрована у цій роботі частка триплоїдів виявилася несподівано низькою: з 17 особин 1 була триплоїдною і 16 — диплоїдними.
У даній роботі ми перевіряли те саме припущення, використовуючи більшу за чисельністю вибірку і поєднуючи косвений метод визначення плоїдності з двома прямими. Косвений метод визначення плоїдності полягає у вимірюванні середнього розміру еритроцитів кожної особини. Як відомо, ядра клітин триплоїдних гібридів містять у півтора рази більше хроматину (Plötner, 2005); це призводить до збільшення розмірів ядер і клітин загалом. Межа між розмірами диплоїдних і триплоїдних еритроцитів у різних регіонах проживання триплоїдних P. esculentus може дещо відрізнятися. Раніше в ході досліджень розмірів еритроцитів у напівдорослих і статевозрілих зелених жабах з Північно-Донецького центру їх різноманіття було встановлено, що гранична довжина еритроцитів, що розділяє диплоїдів і триплоїдів, становить 26–28 μм (Бондарева та ін., 2012). Крім того, було показано, що розмір еритроцитів у неполовозрілих жаб може залежати від довжини тіла (Іскендерова та ін., 2012), а довжина еритроцитів у неполовозрілих триплоїдів може не досягати граничного розміру, встановленого для більших особин (Темников та ін., 2012). Прямими методами визначення плоїдності були кариоаналіз і дослідження ядерцевих органайзерів в інтерфазних ядрах (див. далі).
Матеріали та методи
Для перевірки висунутих гіпотез нами була досліджена вибірка з 40 метаморфів (цьоголіток, які нещодавно завершили метаморфоз) зелених жаб, відловлених у руслі р. Северський Донец в околицях с. Гайдари Зміївського району Харківської області в кінці липня 2013 р. Вибірка складалася з особин довжиною тіла від 18,5 до 33,3 мм. У відловлених метаморфів був взятий мазок крові. 8 особин з технічних причин не були досліджені. У решти жаб взяли фрагменти кишечника для кариологічних досліджень, але у 2 особин матеріал виявився непридатним для аналізу, тому їх також виключили.
Отримання та дослідження мазків крові проводили за опублікованою методикою (Бондарева та ін., 2012).
Для кариоаналізу досліджуваним метаморфам вводили 0,04 % розчину колхіцину внутрішньо‑черевно (по 0,04–0,08 мл на особину залежно від ваги). Через 24 години тварин знеболювали етілатом. Під час розтину вилучали фрагмент кишечника. Отриманий матеріал витримували 20 хв у гіпотонічному розчині (0,07 M KCl), а потім переносили у фіксатор Карнуа (3 частини метанолу і 1 частина крижаного оцтового розчину). Препарати виготовляли методом розкопування на нагрівальному столі при 60 °C. Для цього фрагмент тканини занурювали у 70 % розчин оцтової кислоти, внаслідок чого тканина ставала більш пухкою, і досліджуваний фрагмент розпадався на окремі клітини, утворюючи суспензію. Потім пастерською піпеткою наносили краплі діаметром до 1 см на нагреті предметні скляри. Нанесені краплі одразу забирали піпеткою, залишаючи на склі невелику кількість окремих клітин. Препарати сушили і витримували в термостаті 3 тижні при 37 °C.
Плоїдність визначали підрахунком кількості хромосом (26 для диплоїдів і 39 для триплоїдів) у щонайменше семи метафазних пластинках для кожної особини. На виготовлених препаратах застосовували Ag‑окрашування (серебріння) — метод, що дозволяє виявити та визначити положення ділянки ядерцевого органайзера (РЯО, сайти 18S + 28S рДНК) у хромосомі (Birshtein, 1984) (рис. 4). У ядрах соматичних клітин, що перебувають у інтерфазі, у диплоїдних особин внаслідок Ag‑окрашування виявляються ядерця, помітні як два чітко забарвлених ділянки (рис. 3) (Schmid, 1982). У триплоїдних особин виявляються три такі ділянки, що робить зазначений метод придатним для визначення плоїдності. Для особин, на кариологічних препаратах яких не було метафазних пластинок, придатних для аналізу, плоїдність визначали підрахунком кількості ядерцевих органайзерів у щонайменше 20 ядрах.
Ми використовували метод серебріння (Birshtein, 1984) з модифікаціями. Для фарбування приготували 30 % водний розчин AgNO₃, який перед використанням фільтрували через шприцевий фільтр Simplepure NY (пори 0,22 μм). Потім 200 мг желатину розчиняли в 10 мл теплої води. Після розчинення до желатину додавали 100 μl мурав’їної кислоти. На препарати наносили 75 μl розчину желатину, поверх якого — 150 μl розчину AgNO₃. Препарати накривали покривними склярами і поміщали у вологу світлонепроникну камеру, яку залишали в термостаті 3 хв при 60 °C, після чого витягали, промивали під проточною водою і сушили. Далі препарати фарбували у 2 % розчині барвника Гімза. Після висихання препарати мікроскопували і підраховували кількість ядерцевих органайзерів в інтерфазних ядрах.
Результати та обговорення
На підставі значення середньої довжини еритроцитів кожного метаморфа було зроблено припущення, що всі досліджувані особини є диплоїдними. Середні розміри еритроцитів метаморфів коливалися від 15,38 до 22,56 μм, що значно нижче встановленої межі між розмірами еритроцитів ди‑ і триплоїдів у 26–28 μм (Бондарева та ін., 2012). Для порівняння, середні розміри еритроцитів дорослих диплоїдних особин у нормі становлять 21–26 μм. Варто зазначити, що один неполовозрілий триплоїд, вивчений нами раніше у вибірці з 17 особин, зібраній у тому ж районі, при довжині тіла 48 мм мав середню довжину еритроцитів 23,5 μм, що перевищує середній розмір еритроцитів усіх розглянутих у цій роботі жаб, але значно менше межі, встановленої для статевозрілих особин (Темников та ін., 2012).
У вивченій вибірці залежність між розміром тіла жаб і середньою довжиною їх еритроцитів (рис. 1) виявилася незначущою (р = 0,36).
На мазках крові метаморфів спостерігали як малі овальні еритроцити, так і більші округлі клітини. Така картина може бути пов’язана з тим, що розглянуті особини лише завершили метаморфоз і, можливо, ще мали кров’яні клітини, характерні для личинок (Hollyfield, 1966) (рис. 2).
Отже, визначення плоїдності неполовозрілих особин за середньою довжиною еритроцитів виявляється ненадійним. Метод потребує додаткових досліджень і визначення межі між розмірами еритроцитів різної плоїдності у особин одного розмірного класу або однієї стадії розвитку. Водночас Ag‑окрашування кариологічних препаратів соматичних тканин дозволяє визначити плоїдність навіть за умов нестачі або відсутності метафазних пластинок.
За результатами кариологічного аналізу всі метаморфи, вивчені в цій роботі, також виявилися диплоїдними. При цьому у препаратах, отриманих від 13 особин, виявлено метафазні пластинки, що містять по 26 хромосом, що відповідає диплоїдному набору. Для решти 17 метаморфів висновок про плоїдність був зроблений на підставі наявності двох яскраво забарвлених ядерцевих органайзерів в інтерфазних ядрах (рис. 3).
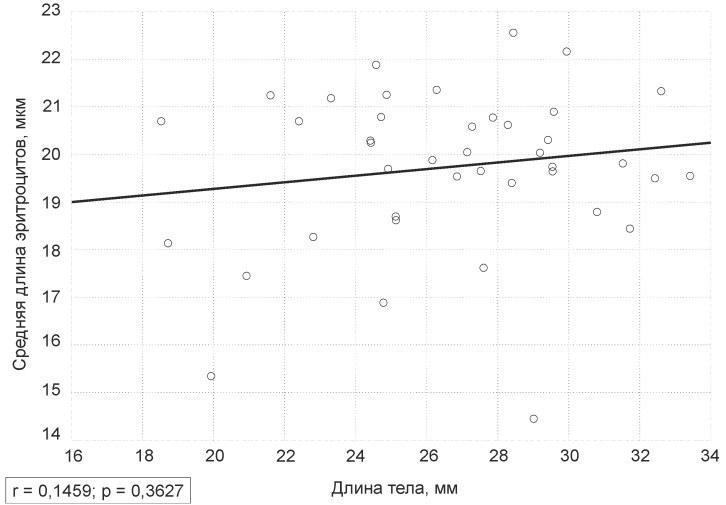
Рис. 1. Графік залежності середньої довжини еритроцитів жаб від довжини їх тіла
Fig. 1. The plot of the frogs’ average red blood cells length dependence on their body length
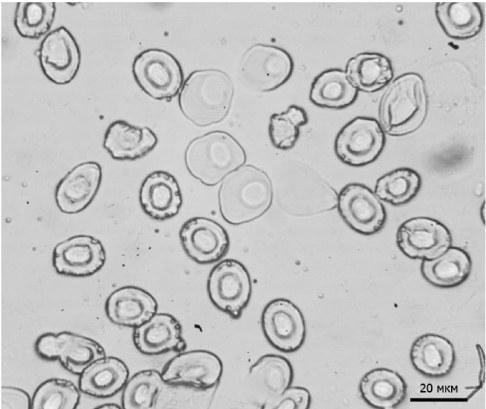
Рис. 2. Еритроцити, різні за формою та розміром, у мазку крові однієї особини
Fig. 2. The red blood cells of different shapes and sizes in the blood smear of one individual
Обговорюючи питання причин відсутності триплоїдів у вивченій вибірці, слід зазначити, що, на відміну від процитованих вище робіт (Borkin et al., 2004; Mezhzherin et al., 2010; Козак та ін., 2012), у цій роботі вибірка складалася не лише з гібридів, а й, ймовірно, з сеголетків P. ridibundus. Визначення видової приналежності метаморфів за морфологічними ознаками надзвичайно ненадійне через недостатню виразність їх діагностичних ознак. Серед сеголетків усього гібридного комплексу зелених жаб триплоїди мають зустрічатися рідше, ніж у вибірці статевозрілих гібридів.
У районі, де були зібрані метаморфи, співвідношення представників P. ridibundus і P. esculentus близьке до 1:1 (Д. А. Шабанов, особисте повідомлення; Москалев та ін., 2012). Об’єднавши вибірки, вивчені у роботі 2012 р. (Темников та ін., 2012) і в цьому дослідженні, ми можемо встановити, що в об’єднаній групі з 47 сеголетків один триплоїд виявився, а диплоїдними — 46.
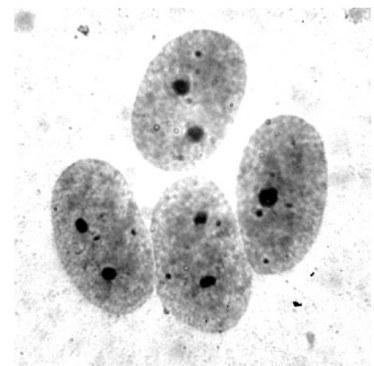
Рис. 3. Епітеліоцити тонкого кишечника з Ag‑окрашеними ядерцевими органайзерами
Fig. 3. Small intestine epithelial cells with silver nitrate stained nucleoli

Рис. 4. Метафазна пластинка (26 хромосом) з Ag‑окрашеними РЯО
Fig. 4. Metaphase plate (26 chromosomes) with silver nitrate stained NORs
Отриманий результат дає підставу на даному етапі дослідження відхилити гіпотезу, що перевірялась у цій роботі. Щоб узгодити низьку частку триплоїдів, зареєстровану в нашому дослідженні, з результатами публікацій, де вказується значно вища частка триплоїдів серед P. esculentus, необхідно припустити дію якихось неучтених нами факторів. Досліджена нами сукупність метаморфів або суттєво відрізняється за часткою входять у її склад триплоїдів, або суттєво відрізняється за співвідношенням P. ridibundus і P. esculentus від вивчених раніше вибірок старших особин.
Для пояснення отриманого результату можна висунути два припущення.
1. Можливо, серед метаморфів і сеголетків частка триплоїдів може бути не більшою, а, навпаки, меншою, ніж у старших вікових класах. Відомо, що при схрещуванні гібридів, клонально передаючих гамети одного й того ж батьківського виду, відокремлюються особини цього батьківського виду. Це явище назвали гібридолізом. Однак диплоїдні особини, що виникають внаслідок гібридолізу, виявляються маложиттєздатними і гинуть до досягнення статевої зрілості (Plötner, 2005). Якщо цей процес відбувається у вивченій нами популяційній системі досить часто, він може призводити до збільшення частки триплоїдів у старших вікових класах за рахунок селективної загибелі певної частини диплоїдних особин (представників батьківських видів, що відокремилися внаслідок схрещування гібридів). Оскільки значна частина гамет, що виробляються P. esculentus у вивченій популяційній системі, належить P. ridibundus (Боркін та ін., 2005), суттєва частка гібридолізних особин, що належать до цього виду, може гинуть після метаморфози.
2. Отриманий результат може бути наслідком нерівномірного розподілу по руслу сеголетків, що належать до батьківського виду або до триплоїдних гібридів. Раніше було показано, що розподіл статевозрілих жаб по території русла не є випадковим, не залежить від їх генотипу. Представники різних форм і різних генотипів концентруються в різних мікромісцезнаходженнях і демонструють різну поведінку (Коршунов, 2010; Москалев та ін., 2012).
Висловлені припущення в подальшому можуть бути перевірені на більш широкому матеріалі з використанням методів, застосованих у цій роботі.
Подяки
Автори висловлюють щиру подяку доценту Д. А. Шабанову, як автору ідеї та науковому керівнику даної роботи.
Список літератури
Бондарева А. А., Бібік Ю. С., Самило С. М., Шабанов Д. А.Цитогенетичні особливості еритроцитів зелених жаб з Північно-Донецького центру різноманіття Pelophylax esculentus complex // Вісник Харківського національного університету імені В. Н. Каразіна. Серія: біологія. – 2012. – Вип. 15 (№ 1008) – С. 116–123. /Bondareva A.A., Bibik Yu.S., Samilo S.M., Shabanov D.A. Tsitogeneticheskiye osobennosti eritrotsitov zelenykh lyagushek iz Seversko-Donetskogo tsentra raznoobraziya Pelophylax esculentus complex // Vіsnyk Kharkіvs'kogo natsіonal'nogo unіversytetu іmenі V.N.Karazіna. Serіya: bіologіya. – 2012. – Vyp.15 (№1008) – S. 116–123.
Боркін Л.Я., Зіненко А.І., Коршунов А.В. та ін. Масова поліплоїдія в гибридогенному комплексі Rana esculenta (Ranidae, Anura, Amphibia) на Сході України // Мат. I конф. Українського Герпетологічного Товариства – К.: Зоомузей ННПМ НАНУ, 2005. – С. 23–26. /Borkin L.Ya., Zinenko A.I., Korshunov A.V. i dr. Massovaya poliploidiya v gibridogennom komplekse Rana esculenta (Ranidae, Anura, Amphibia) na Vostoke Ukrainy // Mat. I konf. Ukrains'kogo Gerpetologіchnogo Tovarystva – K.: Zoomuzey NNPM NANU, 2005. – S. 23–26.
Іскендерова Н.Е., Павлюк О.В., Чебукина М.В. Залежність розмірів еритроцитів від розмірів зелених жаб (Pelophylax esculentus complex) // «Біологія: від молекули до біосфери». Мат. VII Міжнар. конф. мол. науковців. – Х.: ФОП Шаповалова Т.М., 2012 – С. 256–257. /Iskenderova N.Ye., Pavlyuk O.V., Chebukina M.V. Zavisimost' razmerov eritrotsitov ot razmerov zelenykh lyagushek (Pelophylax esculentus complex) // «Bіologіya: vіd molekuly do bіosfery». Mat. VII Mіzhnar. konf. mol. naukovtsіv. – Kh.: FOP Shapovalova T.M., 2012 – S. 256–257.
Козак Н.А., Гладкова Ю.Д., Кулимова М.Д. Визначення частки триплоїдів серед Pelophylax esculentus у НПП «Гомольські ліси» та його околиць // «Біологія: від молекули до біосфери». Мат. VII Міжнар. конф. мол. науковців. – Х.: ФОП Шаповалова Т.М., 2012 – С. 261–262. /Kozak N.A., Gladkova Yu.D., Kulymova M.D. Opredeleniye doli triploidov sredi Pelophylax esculentus v NPP «Gomol'shanskiye lesa» i yego okrestnostyakh // «Bіologіya: vіd molekuly do bіosfery». Mat. VII Mіzhnar. konf. mol. naukovtsіv. – Kh.: FOP Shapovalova T.M., 2012 – S. 261–262.
Коршунов А.В. Екологічні закономірності розподілу Pelophylax esculentus complex у біотопах басейну верхнього течіння річки Північний Донец. Автореф. дис. … к.б.н./03.00.16 – екологія. – Дніпропетровський національний університет імені Олеся Гончара. – Дніпропетровськ, 2010. /Korshunov A.V. Ekologicheskiye zakonomernosti raspredeleniya Pelophylax esculentus complex v biotopakh basseyna verkhnego techeniya reki Severskiy Donets. Avtoref. diss. … k.b.n./03.00.16 – ekologiya. – Dnepropetrovskiy natsional'nyy universitet imeni Olesya Gonchara. – Dnepropetrovsk, 2010.
Михайлова О.В., Кечеджи А.Е., Шабанов Д.А. Вивчення сперматогенезу у диплоїдних Pelophylax esculentus (Amphibia, Anura) за допомогою кариоаналізу в роздавлених препаратах // Праці Українського герпетологічного товариства. – 2011. – № 3. – С. 120–127. /Mikhaylova O.V., Kechedzhi A.E., Shabanov D.A. Izucheniye spermatogeneza u diploidnykh Pelophylax esculentus (Amphibia, Anura) pri pomoshchi karioanaliza v razdavlennykh preparatakh // Pratsі Ukrains'kogo gerpetologіchnogo tovarystva. – 2011. – №3. – S. 120–127.
Москалев В.Б., Каширін О.О., Меламед Р.Г. Залежність складу вибірок зелених жаб від місця і способу їх збору // «Біологія: від молекули до біосфери». Мат. VII Міжнар. конф. мол. науковців. – Х.: ФОП Шаповалова Т.М., 2012 – С. 271–272. /Moskalyov V.B., Kashirin O.O., Melamed R.G. Zavisimost' sostava vyborok zelenykh lyagushek ot mesta i sposoba ikh sbora // «Bіologіya: vіd molekuly do bіosfery». Mat. VII Mіzhnar. konf. mol. naukovtsіv. – H.: FOP Shapovalova T.M., 2012 – S. 271–272.
Темніков А.В., Вегеріна А.О., Мелешко Є.В., Михайлова О.В. Вивчення плоїдності та розмірів еритроцитів у неполовозрілих представників Pelophylax esculentus complex // «Біологія: від молекули до біосфери». Мат. VII Міжнар. конф. мол. науковців. – Х.: ФОП Шаповалова Т.М., 2012 – С. 276–277. /Temnikov A.V., Vegerina A.O., Meleshko Ye.V., Mikhaylova O.V. Izucheniye ploidnosti i razmerov eritrotsitov u nepolovozrelykh predstaviteley Pelophylax esculentus complex // «Bіologіya: vіd molekuly do bіosfery». Mat. VII Mіzhnar. konf. mol. naukovtsіv. – H.: FOP Shapovalova T.M., 2012 – S. 276–277.
Шабанов Д.А., Коршунов О.В., Кравченко М.О. Які ж зелені жаби населяють Харківську область? Термінологічний і номенклатурний аспекти проблеми // Біологія та валеологія. – Вип. 11. – Харків: ХДПУ, 2009. – С. 116–125. /Shabanov D.A., Korshunov O.V., Kravchenko M.O. Yakі zh zelenі zhaby naselyayut' Kharkіvs'ku oblast'? Termіnologіchnyy і nomenklaturnyy aspekty problemy // Bіologіya ta valeologіya. – Vyp.11. – Kharkіv: KhDPU, 2009. – S. 116–125.
Borkin L. J., Korshunov A. V., Lada G. A., Litvinchuk S. N., Rosanov J. M., Shabanov D. A., Zinenko A. I. Mass occurrence of polyploid green frogs (Rana esculenta complex) in Eastern Ukraine // Russian Journal of Herpetology, 2004. – Vol. 11, No 3. – P. 194–213.
Birstein V.J. Localization of NORs in karyotypes of four Rana species // Genetica. – 1984. – № 64. – P. 149–154.
Hollyfield J.G. Erythrocyte replacement at metamorphosis in the frog Rana pipiens // Journal of Morphology. – 1966. – № 119. – P. 1–6.
Mezhzherin S.V., Morozov‑Leonov S.Yu., Rostovskaya O.V. et al. The ploidy and genetic structure of hybrid population of water frogs Pelophylax esculentus complex (Amphibia, Ranidae) of Ukraine fauna // Cytology and Genetics. – 2010. – Vol. 44, № 4. – P. 212–216.
Plötner J. Die westpaläarktichen Wasserfrösche. – Bielefeld: Laurenti‑Verlag, 2005. – 161 S.
Schmid M. Analysis of the structure and variability of NORs in Anura // Chromosoma. – 1982. – № 87. – P. 327–344.