
Чи є губки найдавнішими багатоклітинними тваринами на Землі?
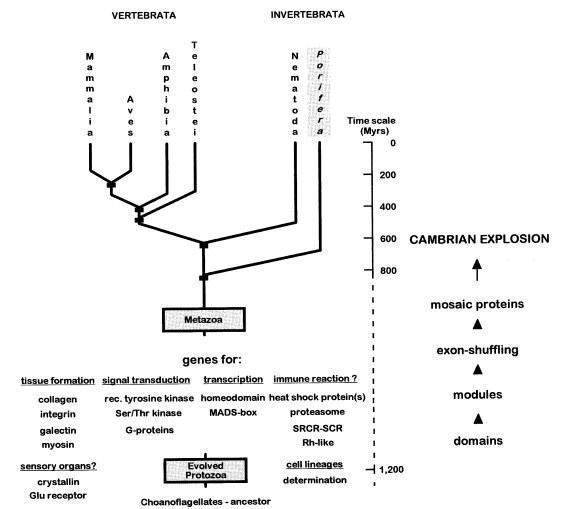
Губки (Porifera) – одна з найдавніших нині живих груп тварин і, можливо, найдавніших тварин, що належать до Metazoa. Високий поліморфізм, викликаний тотипотентністю всіх клітин губок, примітивна будова: відсутність нервової та, ймовірно, м’язової тканини; відсутність світлочутливих органів і клітин (крім хрестоподібних клітин на деяких стадіях ембріогенезу); слабка спеціалізація тканин, наприклад, багато функцій сполучної тканини у складніших тварин виконують лише кілька типів клітин мезоглеї – усе це дозволяє судити про примітивність даної групи. Але також губки володіють добре розвинутим колагеновим матриксом, скоротильною здатністю (оскулярною і тотальною), складними молекулярними комплексами рецепторів на поверхні клітин (тирозин-кіназні, інтегринові), елементами сенсорної системи (метаботропічні глутамінові рецептори, кристалліни), імунною системою (імуноглобуліноподібні молекули, Rh‑система, цистеїнові клітини‑утиліти), що класифікує губки як справжні Metazoa [6].
Проблема «укорінення» всієї групи багатоклітинних тварин є предметом дискусій, і послідовність відгалуження від загальної гілки багатоклітинних губок (Porifera), гребневиків (Ctenophora), пластинчастих (Placozoa), стрекаючих (Cnidaria) і билатерій (Bilateria) достовірно невідома, окрім того, що книдарії та билатерії є найближчими родичами. Інші групи є самостійними і давніми щодо усіх багатоклітинних.
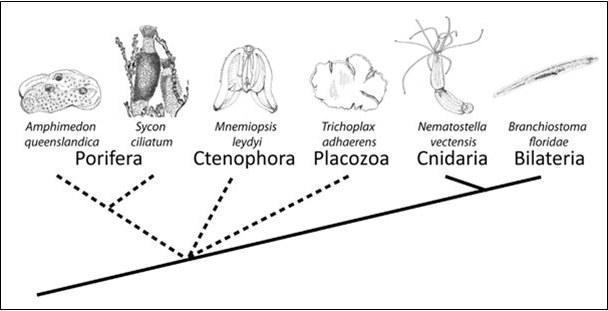
Основна проблема полягає в тому, що з’ясування філогенетичних зв’язків ранніх Metazoa і історії утворення багатоклітинних у цілому ускладнюється через давність таких еволюційних подій. Щодо губок, навіть тут α‑таксономія повністю не відома, так само і філогенетичні відносини всередині групи, отже, навіть якщо ми приймаємо гіпотезу, що саме губки стоять на початку багатоклітинності і дають початок іншим групам, то цю роль може претендувати і окремий клас, наприклад Calcarea (що показано вченими з центру океанології в Марселі). Також їх дані свідчать про те, що клас Hexactinellida у губок не лише морфологічно, фізіологічно, а й філогенетично сильно відрізняється від демоспонгій і вапнякових губок[2].
Дослідження в галузі молекулярної філогенетики дають різні уявлення про те, яке місце губки займають на дереві Metazoa, причиною цього є вибірковість порівнюваних груп. Існує також велика залежність топології дерева від обраної аутґрупи, яка буде укорінювати дерево, а також від статистичних моделей субституцій сайтів у ДНК (по суті, від цього залежить вимір швидкості накопичення мутацій і відображається у силі дивергенції дерева та довжині його гілок).
Показано, що якщо вибирати більш віддалену від багатоклітинних аутґрупу, наприклад гриби (як частину монофілетичної групи Opisthokonta), можливо підтвердження альтернативної гіпотези, що Placozoa знаходяться в корені багатоклітинності, тоді як вибір більш близькородинної групи хоанофлагеллат (Choanoflagellata) дає добре підтримувану bootstrap‑методом топологію, що підтверджує гіпотезу про походження всіх багатоклітинних тварин від предка‑губки [9]. Такий вибір може бути більш доцільним, оскільки хоанофлагеллати відносять до близької щодо багатоклітинних групи не лише завдяки спільним морфологічним ознакам (наприклад, часто порівнюються з хоаноцитами губок[4]), а й підтверджується філогенетичними методами[3,7].
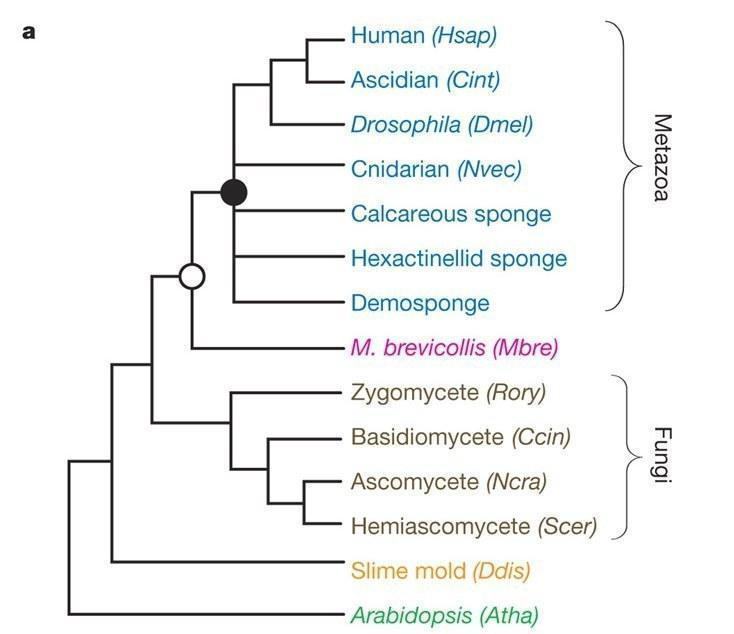
В цілому ж ряд молекулярних філогенетичних досліджень показують, що саме губки стоять у основі Metazoa, при цьому використовувалися різні методи пошуку оптимальних топологій (наприклад, максимальна парсимонія, neighbor‑joining) а також різні моделі субституцій нуклеотидів[2,9].
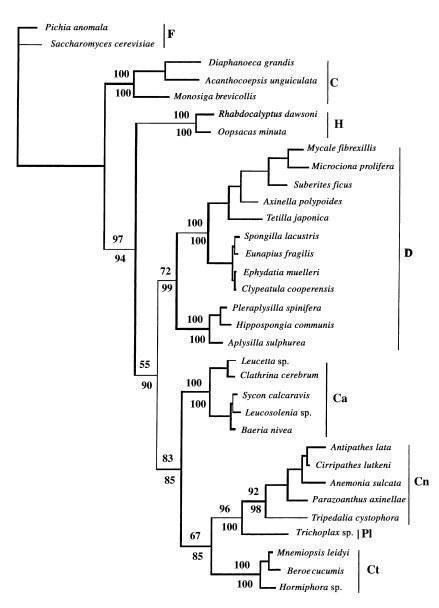
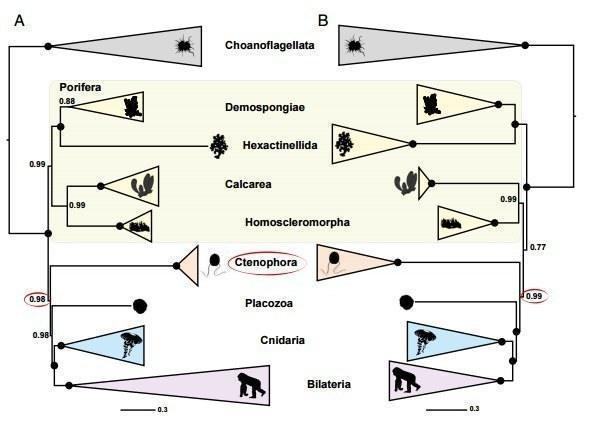
Щодо порівняльної фізіології: ще в кінці 90‑х років вчені з університету у Ерфурті провели вивчення деяких рецепторних молекул, колагену, білків імунної системи губок та їх ДНК‑послідовностей, використовуючи моделі субституцій амінокислот, що також передбачає губок як найдавніших представників багатоклітинних. На підставі аналізу дається датування розходження губок і хоанофлагеллат, і ці оцінки збігаються з іншими методами визначення часу появи губок (ранній кембрій). Цікаво, що передбачається, що можливість багатоклітинності, диференціації клітин і подальший кембрійський вибух у цілому стали можливими після ініціації еволюційно значущого механізму альтернативного сплайсингу (перестановки екзонів у процесі транскрипції) 1000 млн років тому. Порівняння послідовностей, що кодує білок‑кіназу С у цій роботі, вказує на Calcarea як найближчого родича Eumetazoa і відокремлює Hexactinellida від інших губок[6]. Тут також можна вказати на «конкуренцію» гіпотез про первинність гребневиків і губок, і тут виграють губки, оскільки мають ще більш примітивну скоротливу систему (ми знаємо лише про можливу участь у цьому ендопінакоцитів[8]), ніж гребневики. Порівняння синапсів і нейромедіаторів гребневиків з таковими у билатерій, правда, також виявляє суттєві відмінності, але тут може грати роль конвергентна еволюція нервової системи [10].
А що, якщо припустити існування предка‑не‑губки, що мав ще простіше будову, при цьому мав відносно диференційоване багатоклітинне тіло? Це припущення дійсно має підстави, проте виникає ряд проблем, викликаних необхідністю палеонтологічних даних або неможливістю наразі змоделювати такого предка. Таким чином, це питання можна залишити відкритим. А що, якщо такий предок був складнішим?
Недавні дослідження, опубліковані командою австралійських біологів, проведені на базі Міжнародного центру морської молекулярної біології при Бергенському університеті у сфері порівняння транскрипційних факторів ембріогенезу трьох класів губок, передбачають, що відносно примітивна будова тіла губок може бути пов’язана з втратою генів унаслідок еволюції від складнішого предка, який дав початок губкам і кишечнополостним. Такий гіпотетичний спільний предок мав би більш багатий набір генів у порівнюваних генних сімействах (у даному випадку T‑боксні гени, Sox‑гени, гени класу ANTP, Six‑гени, Pax‑гени, GATA та Smad‑гени), які збереглися лише частково для кожного класу губок. Філогенетичний аналіз також показав, що всі гени цих сімейств демонструють схожість з найближчою групою – стрекаючими. Автори також зазначають, що така втрата генів відбувалася в ході еволюції неодноразово у різних таксономічних групах (наприклад, у тих самих кишечнополостних (Hydra) або навіть у хребетних (Tunicata))[1]. Великим плюсом на підтримку гіпотези є те, що розглянуті гени іноді відіграють ключову роль в ембріогенезі губок (наприклад, під час екскурвації кальцієвих губок активно експресується ген Brachyury, який, що цікаво, бере участь і у формуванні осевого скелету у хордових), отже, такі події, як втрата генів цих сімейств або дуплікація, дійсно впливають на хід еволюції і є інформативними для висунутої гіпотези. Водночас це може свідчити лише про те, що багато генів просто змінювалися або модифікувалися в геномі з ходом еволюції, або навіть просто змінилася функція гена, і це не є наслідком втрати генів у складнішого предка. У будь‑якому випадку без палеонтологічних даних це залишається лише гіпотезою.
А що з палеонтологічними даними? Свіженьке дослідження 2016 року від вчених Массачусетського технологічного університету не лише підтверджує гіпотезу про губки як найдавніші багатоклітинні, а й збільшує їх «давність» до докембрійського періоду (близько 640 млн років). Недостатність традиційних викопних губок може бути компенсована великою кількістю «молекулярних» викопних (мінералізованих відкладень). У цьому дослідженні це були відкладення стеролу 24‑ізопропілхолестану, для якого був ідентифікований ген, що відповідає за синтез ліпіду – SMT‑ген. Далі використовувалися методи порівняльної геноміки, за допомогою яких було показано двох можливих донорів ліпіду: водоростей і губок, при цьому методом молекулярних годин (датування еволюційних подій) було показано, що у губок він з’явився раніше, а саме 640 млн років тому[5].
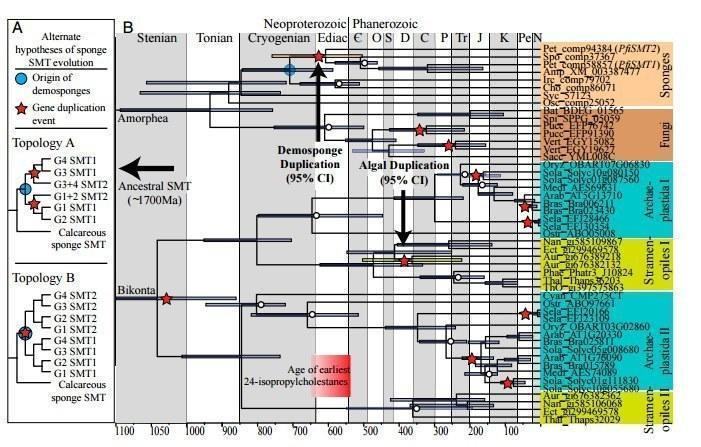
Отже, скоріш за все губки є‑таки найдавнішими багатоклітинними тваринами на Землі, але це породжує безліч додаткових питань. Наприклад, чи є вони першими представниками багатоклітинних, чи просто більш успішною групою, що дожила до наших днів? Якщо так, то якою була ця губка, адже ми маємо неймовірне різноманіття губок, аж до синцитіальних форм (скляні губки), і, можливо, навіть є підстави виділити їх в окремі типи. Принциповим є також питання вивчення їх фізіології в рамках еволюції, оскільки, наприклад, їх скоротливі здібності можуть зіграти велику роль у розумінні того, як розвивалася нервова система на початкових етапах. Відкритим залишається і питання, чи були саме губки предками книдарій і билатерій, чи просто найближчою сестринською групою, що дожила до наших днів. Я вважаю, що для цього ще довго треба збирати молекулярні, палеонтологічні та фізіологічні дані по всіх групах ранніх багатоклітинних, щоб мати можливість змоделювати гіпотетичного предка, який вперше отримав багато спеціалізованих клітин.
Джерела:
Sofia A. V. Fortunato, Marcin Adamski, Maja Adamska Comparative analyses of developmental transcription factor repertoires in sponges reveal unexpected complexity of the earliest animals // Marine Genomics. 2015 (in press)
C. Borchiellini, M. Manuel, E. Alivon, N. Boury‑Esnault, J. Vacelet and Y. Le Parco Sponge paraphyly and the origin of Metazoa // Journal of Evolutionary Biology. 2001. V. 14, Issue 1, P. 171–179.
M. Carr, B. S. C. Leadbeater, R. Hassan, M. Nelson, and S. L. Baldauf Molecular phylogeny of choanoflagellates, the sister group to Metazoa // PNAS. 2008. V. 105(43). P. 16641–16646.
James‑Clark H. Conclusive proofs on the animality of the ciliate sponges, and their affinities with the Infusoria Flagellata. Am J Sci Ser. 2. 1866. V. 42 P. 320–325.
David A. Golda, Jonathan Grabenstattera, Alex de Mendozab, Ana Riesgoc, Iñaki Ruiz‑Trillob,d, and Roger E. Summons Sterol and genomic analyses validate the sponge biomarker hypothesis // PNAS V. 113. No. 10. P. 2684–2689.
Muller W. E. Origin of Metazoa: Sponges as Living Fossils // Naturwissenschaften. 1998. V. 85(1). P. 11‑25.
Nicole King and oth. The genome of the choanoflagellate Monosiga brevicollis and the origin of metazoans // Nature. 2008. V. 451(7180). P. 783‑788.
Michael Nickel, Corina Scheer1, Jörg U. Hammel, Julia Herzen and Felix Beckmann The contractile sponge epithelium sensu lato – body contraction of the demosponge Tethya wilhelma is mediated by the pinacoderm // The Journal of Experimental Biology. 2011. V. 214. P. 1692‑1698.
Davide Pisani, Walker Pett, Martin Dohrmann, Roberto Feuda, Omar Rota‑Stabelli, Hervé Philippe, Nicolas Lartillot and Gert Wörheide Genomic data do not support comb jellies as the sister group to all other animals // Proceedings of the National Academy of Sciences. 2015. V. 112. No 50. P. 15402‑15407.
Ястребов С. А. Порівняльна геноміка змушує переглянути місце гребневиків на еволюційному дереві тварин // Елементи, 2015.