
Бондарєва та ін. (2010) Чи впливають розмірні ефекти клітин... ?
Статья, написанная вместе с двумя студентками. Работа с эритроцитами, начатая этой статьей, привела к неожиданному результату, отраженному в наших с Бондаревой тезисах 2011 года. Бондарєва А. А., Махній Т. I., Шабанов Д. А. Чи впливають розмірні ефекти клітин диплоїдних та триплоїдних їсті...
Бондарєва А. А., Махній Т. I., Шабанов Д. А. Чи впливають розмірні ефекти клітин диплоїдних та триплоїдних їстівних жаб на їх стійкість до гіпоксії? // Біологія та валеологія. – Вип. 12 – Харків: ХДПУ, 2010. с. 10-15.
ЧИ ВПЛИВАЮТЬ РОЗМІРНІ ЕФЕКТИ КЛІТИН ДИПЛОЇДНИХ ТА ТРИПЛОЇДНИХ ЇСТІВНИХ ЖАБ НА ЇХ СТІЙКІСТЬ ДО ГІПОКСІЇ?
Бондарєва А.А., Махній Т.I., Шабанов Д.А.
Харківський національний університет ім. В.Н. Каразіна
Диплоїди та триплоїди Pelophylax esculentus, що мешкають разом у Сіверсько-Донецькому центрі різноманіття зелених жаб, є вдалою моделлю для вивчення впливу розмірів клітин на фізіологічні процеси. Перевірено припущення про те, що триплоїди відрізняються від диплоїдів за стійкістю до гіпоксії. Це припущення не підтвердилося. Показано, що розмір еритроцитів жаб позитивно пов'язаний з розміром їхнього тіла. На початку осені у деяких жаб реєструється значне збільшення стійкості до гіпоксії; це, вірогідно, пов’язано з їхньою фізіологічною підготовкою до зимівлі.
Ключові слова: їстівні жаби, еритроцити, гіпоксія, триплоїдія, розмірні ефекти клітин.
Do dimensional effects of diploid and triploid edible frogs cells influence on their resistance to hypoxia? Bondaryeva A.A., Makhnii T.I., Shabanov D.A. Diploids and triploids of Pelophylax esculentus inhabiting the green frogs diversity centre in the Siversky Donets basin are good model for study of influence of cell size on physiological processes. The hypothesis that triploids differ from diploids in their resistance to hypoxia was tested. This hypothesis was not confirmed. It was shown that the sizes of frog erythrocytes are positive related to their body sizes. In early autumn it is registered a significant increase in resistance to hypoxia in some frogs; presumably, this is related to their physiological preparation for wintering.
Key words: edible frogs, erythrocytes, hypoxia, triploidy, dimensional effects of cells.
Вступ. До гібридогенного комплексу зелених жаб (Pelophylax esculentus complex) входять два батьківських види (ставкові жаби, Pelophylax lessonae (Camerano, 1882) і озерні жаби Pelophylax ridibundus (Pallas, 1771)). Схрещуючись, ці види утворюють гібриди: їстівних жаб, Pelophylax esculentus (Linnaeus, 1758). P. esculentus розмножуються геміклонально (напівклонально): в гамети у них переходить лише один з батьківських геномів (хромосомних наборів), який передається з покоління у покоління без рекомбінації (3, 6).
Особливий інтерес викликає знайдений у Східній Україні Сіверсько-Донецький центр різноманіття зелених жаб. Тут поряд одні з одними мешкають диплоїдні, триплоїдні та навіть поодинокі тетраплоїдні P. esculentus (2, 3). Триплоїдні P. esculentus поширені і в Західній Європі, алепереважноіснують у популяційних системах іншого складу (6). У триплоїдів у ядрах клітин міститься в півтора рази більше хроматину, ніж у диплоїдів. Вірогідно, саме з цим пов’язане збільшення розмірів ядер клітин триплоїдів та, внаслідок збереження ядерно-цитоплазматичного відношення, збільшення і самих клітин. Збільшені розміри еритроцитів триплоїдних P. esculentus застосовують для їх експрес-діагностування (6, 7). Оскільки триплоїди практично не відрізняються за розміром тіла від диплоїдів, можна стверджувати, що триплоїди побудовані з меншої кількості більших за розміром клітин.
Оскільки гібридні жаби розмножуються геміклонально, генетичні відміни диплоїдів і триплоїдів P. esculentus виражаються лише у різній кількості геномів батьківських видів, а не у генетичній дивергенції генотипів. Вірогідно, попри відміни у розмірі клітин, ди- та триплоїди побудовані з однакових макромолекул. Зміни триплоїдів на клітинному та органо-тканинному рівнях в порівнянні з диплоїдами мають викликати закономірні порушення у функціях організму, особливо у тих процесах, для яких важливими є кількість клітин та співвідношення їх об’єму та поверхні. Однією з фізіологічних систем, функціонування яких в триплоїдів має зазнавати змін, є система газообміну. Як функціонує кров триплоїдів P. esculentus, яка, порівняно з диплоїдами, несе збільшені еритроцити зі зменшеною питомою поверхнею? Одним з показників функціонування системи газообміну є пристосування до гіпоксії.
Різниця у стійкості до гіпоксії показана для риб, у яких зустрічаються диплоїди та триплоїди (як для Oncorhynchus mykiss та Scophthalmus maximus). У триплоїдів риб знижена кількість еритроцитів, збільшені їхні розміри та зменшена концентрація гемоглобіну. Це викликає зниження стійкості до гіпоксії (4, 8). Пристосування до гіпоксії у риб пов’язане й з розмірами еритроцитів. Для скорпен (Scorpaena porcus L.) показано збільшення об’єму еритроцитів, їх ядер та ядерно-цитоплазматичного відношення за умов довготривалої гіпоксії. Це відбувається внаслідок свелінгу (набрякання клітин) (1).
Таким чином, актуальними є порівняння фізіологічних особливостей ди- та триплоїдних P. esculentus. Ми почали з вивчення стійкості жаб до гіпоксії.
Основна частина. Метою нашої роботи було порівняння стійкості до гіпоксії у ди- та триплоїдних P. esculentus та визначення залежності цієї стійкості від лінійних розмірів еритроцитів. Дослід проводили на двох групах жаб, зібраних в заплаві ріки Сіверський Донець в Зміївському районі Харківської обл. Перша група складалася з 25 особин, що були зібрані в середині липня 2010 р, друга група складалася з 28 особин, зібраних наприкінці серпня того ж року. В обох групах були наявні як самки, так і самці.
Для кожної тварини було зроблено мазок крові. Краплю крові, отриману з пальця жаби, наносили на предметне скло та розмазували тонким шаром ребром іншого предметного скла. Мазки висушували та фотографували під великим збільшенням мікроскопу USB-камерою, з’єднаною з комп’ютером. За тих же умов сфотографували об’єкт-мікрометр. Вимірювання великої вісі 15–20 еритроцитів проводили на фотографіях з використанням програми PDF‑XChange Viewer. Результати вимірювань перераховували у мікрометри.
Кожну особину поміщували в умови штучної гострої екзогенної гіпоксії. Щоб заблокувати обмін кисню через шкіру жаб, їх поміщували у дегазовану воду. Для дегазації відстояну водопровідну воду кип’ятили, а потім остуджували до кімнатної температури у закритих ємностях. У першій серії експериментів жаб поміщували у закриту банку ємністю 200 мл, наповнену водою до кришки. У другій серії дослідів використовували банки ємністю 500 мл, утримуючи жаб на відстані від поверхні за допомогою сітки. Мірою стійкості до гіпоксії був час перебування тварини в описаних умовах в активному стані. Момент переходу у неактивний стан визначали по припиненню рухів жаби та закочуванню її очей. Одразу після цього банку відкривали і витягували тварину на відкрите повітря. У всіх випадках через кілька секунд після контакту шкіри з повітрям жаба відкривала очі та приймала нормальну позу. Результати вимірювань статистично обробляли за допомогою програми Statistica 8.0.
Перша вибірка жаб виявилася достатньо одноманітною за розміром еритроцитів (21,85 ±1,83 мкм). Судячи з цього, дана вибірка складалася лише з диплоїдів. Середній час перебування у воді становив 342 с. Значних відхилень від цього значення не спостерігалося. На цій вибірці встановлено наявність кореляції між розміром еритроцита та довжиною тіла (рис. 1). Коефіцієнт кореляції Спірмена для цих параметрів становить R = 0,42 (p = 0,036). Вплив розміру еритроцитів на стійкість до гіпоксії не зареєстрований.
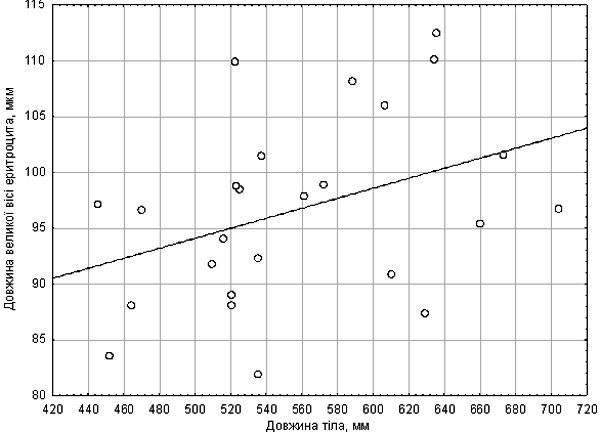
Рис. 1. Залежність розміру еритроцита від довжини тіла (перша вибірка жаб)
Друга вибірка складалася з жаб, більш різноманітних за розміром еритроцитів (від 17,65 до 30,51 мкм). У вибірці було виявлено 5 ймовірних триплоїдів (розмір еритроцитів більше 26 мкм) (6).
В розподілі за часом збереження активності в умовах штучної гіпоксії спостерігалося два піки (рис. 2). Ймовірні триплоїди демонстрували як порівняно невеликий час перебування у воді (587 с), так і дуже великий (3060 с). Як видно по рис. 2, на деяких з досліджуваних особин впливав фактор, що збільшував час витримки умов гіпоксії приблизно вдвічі. Оскількидруга серія експериментів проводилася на початку осені, вірогідно, цей фактор — фізіологічна підготовка до зимівлі.
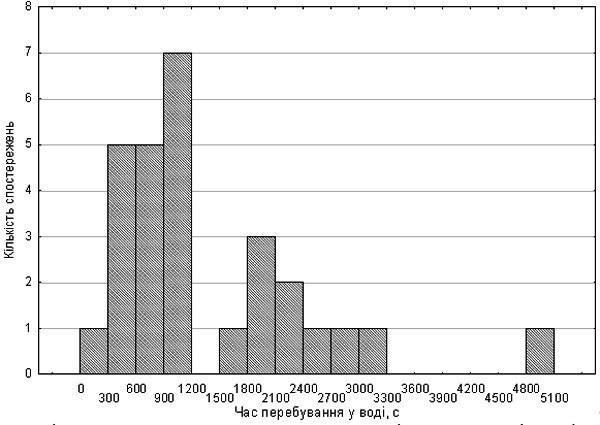
Рис. 2. Розподіл за часом збереження активності в умовах гіпоксії жаб другої вибірки
Вплив розміру еритроцитів на стійкість до гіпоксії не зареєстрований (рис. 3). Спостерігається лише слабкий та статистично незначущій зв'язок між розміром еритроцитів та часом витримки гіпоксії.
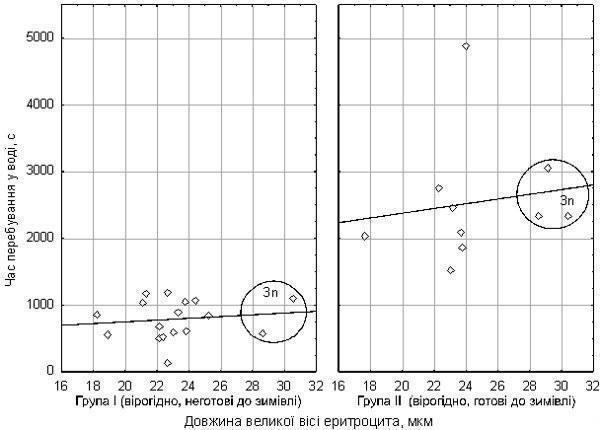
Рис. 3. Залежність часу перебування в умовах гіпоксії від розміру еритроцитів жаб. Дві групи жаб (за прийнятою нами гіпотезою) показано окремо
Висновки. Незважаючи на те, що в наших експериментах проявилася дія незапланованого фактора (вірогідно, підготовки до зимівлі), ми можемо стверджувати, що значних відмін у стійкості до гіпоксії між диплоїдами та триплоїдами P. esculentus не знайдено. Ми можемо передбачати наявність у триплоїдів специфічних механізмів компенсації розмірних ефектів клітин, які потребують подальшого вивчення.
Робота виконана за підтримки Фонду фундаментальних, прикладних та пошукових науково-дослідних робіт Харківського національного університету імені В.Н. Каразіна.
Література.
1. Парфьонова І.А., Солдатов А.А. Еритрограма циркулюючої крові скорпени в умовах експериментальної гіпоксії. // Морський екологічний журнал. – 2005. – Т. IV, №2.
2. Шабанов Д. А., Коршунов О. В., Кравченко М. О. Які ж зелені жаби населяють Харківську область? Термінологічний і номенклатурний аспекти проблеми // Біологія та валеологія. – Вип. 11. – Харків: ХДПУ, 2009. – С. 164–125.
3. Шабанов Д.А., Литвинчук С.Н. Зелені жаби: життя без правил чи особливий спосіб еволюції? // Природа. – 2001. – №3. – С. 29 – 36.
4. Cal R.M., Vidal S., et al. Effect of triploidy on turbot haematology. // Comparative Biochemistry and Physiology, Part A 141 (2005). 35 – 41.
5. George S.A., Michelle R. Lennartz. Methods for determining ploidy in amphibians: nucleolar number and erythrocyte size. // Experientia. – 1980. – р. 687 –688.
6. Plötner J. Die Westpaläarktichen Wasserfrösche. Bielefeld, 2005.
7. Polls Pelaz M., Graf J.-D. Erythrocyte size as an indicator of ploidy level in Rana kl. esculenta before and after metamorphosis. // Alytes 1988. – 7. – p. 53 – 61.
8. Maxime V., Labbé L. The effect of ploidy and sexual maturation on the resistance of erythrocytes to haemolysis in rainbow trout (Oncorhynchus mykiss). // Aquaculture. – 2010. – 305.
Чи впливають розмірні ефекти клітин диплоїдних і триплоїдних їстівних жаб на їх стійкість до гіпоксії? Бондарєва А.А., Махній Т.І., Шабанов Д.А. – Диплоїди і триплоїди Pelophylax esculentus, що спільно мешкають у Сіверсько-Донецькому центрі різноманіття зелених жаб, є вдалою моделлю для вивчення впливу розмірів клітин на фізіологічні процеси. Перевірено припущення, що триплоїди відрізняються від диплоїдів за стійкістю до гіпоксії. Це припущення не підтвердилося. Показано, що розмір еритроцитів жаб позитивно пов’язаний з розміром їх тіла. На початку осені у деяких жаб реєструється значне збільшення стійкості до гіпоксії; це, ймовірно, пов’язано з їх фізіологічною підготовкою до зимівлі.
Ключові слова: їстівні жаби, еритроцити, гіпоксія, триплоїдія, розмірні ефекти клітин.