
Bondareva et al. (2010) Do size effects of cells influence... ?
Article written together with two female students. The work with erythrocytes, initiated by this article, led to an unexpected result, reflected in our theses with Bondareva from 2011. Bondareva A. A., Makhnij T. I., Shabanov D. A. Do size effects of cells of diploid and triploid fish affect...
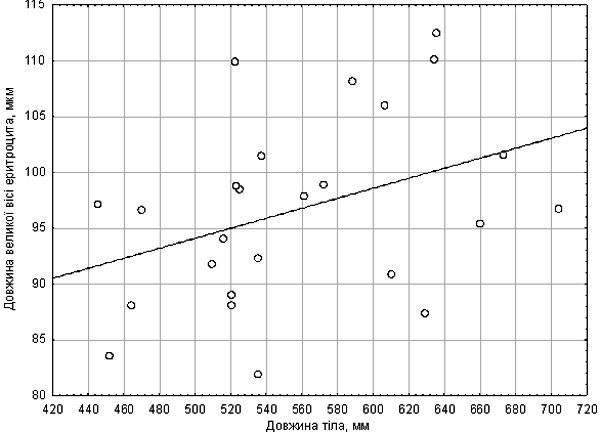
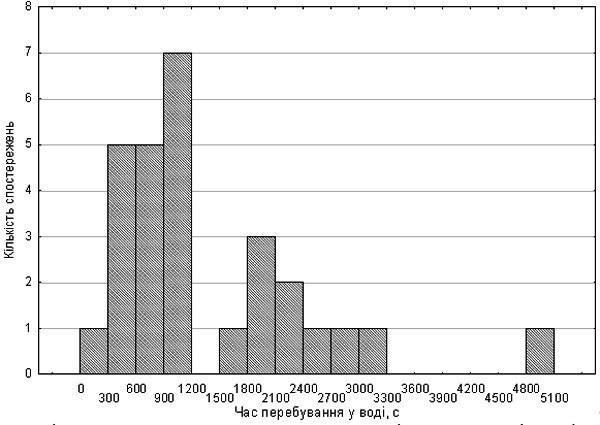
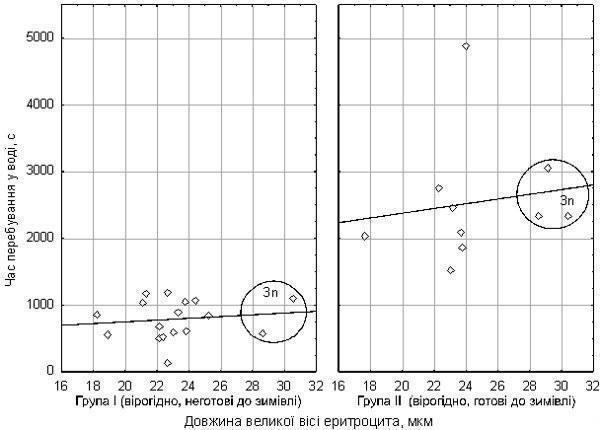
{ "translated_text": "Bondaryeva A.A., Makhnii T.I., Shabanov D.A. // Biology and Valiology. – Issue 12 – Kharkiv: HDPU, 2010. pp. 10-15. DO DIMENSIONAL EFFECTS OF DIPLOID AND TRIPLOID EDIBLE FROGS CELLS INFLUENCE THEIR RESISTANCE TO HYPOXIA? Bondaryeva A.A., Makhnii T.I., Shabanov D.A. V. N. Karazin Kharkiv National University Diploids and triploids of Pelophylax esculentus co‑inhabiting the Siversko‑Donets green frog diversity centre constitute a suitable model for studying the influence of cell size on physiological processes. The hypothesis that triploids differ from diploids in hypoxia resistance was tested. This hypothesis was not confirmed. It was shown that frog erythrocyte size is positively correlated with body size. In early autumn a significant increase in hypoxia resistance is recorded in some frogs, presumably related to their physiological preparation for overwintering. Key words: edible frogs, erythrocytes, hypoxia, triploidy, dimensional effects of cells. Introduction. The hybridogenic complex of green frogs (Pelophylax esculentus complex) includes two parental species (the marsh frog, Pelophylax lessonae (Camerano, 1882) and the lake frog Pelophylax ridibundus (Pallas, 1771)). By hybridising, these species produce hybrids: the edible frog, Pelophylax esculentus (Linnaeus, 1758). P. esculentus reproduces hemiclonally (semiclonally): only one of the parental genomes (chromosomal sets) is transmitted to the gametes and is passed from generation to generation without recombination (3, 6). Particular interest is evoked by the Siversko‑Donets centre of green frog diversity discovered in Eastern Ukraine. Here diploid, triploid and even occasional tetraploid P. esculentus coexist (2, 3). Triploid P. esculentus are also widespread in Western Europe, predominantly existing in population systems of a different composition (6). In triploids, cell nuclei contain one and a half times more chromatin than in diploids. It is likely that this underlies the increase in nuclear size of triploid cells and, due to preservation of the nucleocytoplasmic ratio, the increase in cell size itself. Enlarged erythrocytes of triploid P. esculentus are used for their expression‑diagnostics (6, 7). Since triploids are practically indistinguishable in body size from diploids, it can be argued that triploids are built from a smaller number of larger cells. Because hybrid frogs reproduce hemiclonally, genetic differences between diploid and triploid P. esculentus are expressed only in the number of parental genomes, not in genotype divergence. Presumably, despite differences in cell size, diploids and triploids are constructed from the same macromolecules. Cellular and tissue‑level changes in triploids compared with diploids should cause systematic disturbances in organismal functions, especially in processes where cell number and the ratio of volume to surface are important. One physiological system expected to undergo changes in triploids is the respiratory system. How does the blood of triploid P. esculentus function, which, compared with diploids, carries enlarged erythrocytes with reduced specific surface? One indicator of respiratory system performance is adaptation to hypoxia. Differences in hypoxia tolerance have been shown for fish in which diploids and triploids occur (e.g., Oncorhynchus mykiss and Scophthalmus maximus). Triploid fish exhibit reduced erythrocyte numbers, increased cell size and decreased hemoglobin concentration, leading to lowered hypoxia tolerance (4, 8). Hypoxia adaptation in fish is also linked to erythrocyte size. In the scorpionfish (Scorpaena porcus L.) an increase in erythrocyte volume, nuclei and nucleocytoplasmic ratio under prolonged hypoxia has been demonstrated, resulting from cell swelling (1). Thus, comparing physiological traits of diploid and triploid P. esculentus is pertinent. We began by studying frog hypoxia tolerance. Materials and Methods. The aim of our work was to compare hypoxia tolerance in diploid and triploid P. esculentus and to determine its dependence on linear erythrocyte dimensions. The study was conducted on two groups of frogs collected from the floodplain of the Siverskyi Donets River in Zmiiv district, Kharkiv region. The first group comprised 25 individuals collected in mid‑July 2010; the second group comprised 28 individuals collected at the end of August the same year. Both sexes were represented in each group. For each animal a blood smear was prepared. A drop of blood obtained from a frog’s toe was placed on a slide and spread into a thin layer with the edge of another slide. Smears were dried and photographed at high magnification using a USB‑camera microscope attached to a computer. Under the same conditions, a micrometer slide was photographed. Measurements of the major axis of 15–20 erythrocytes were performed on the photographs using PDF‑XChange Viewer, and results were converted to micrometres. Each individual was subjected to acute exogenous hypoxia. To block cutaneous oxygen exchange, frogs were placed in degassed water. For degassing, tap water was boiled and then cooled to room temperature in sealed containers. In the first experimental series, frogs were placed in a sealed 200 ml jar filled to the brim with water. In the second series, 500 ml jars were used, keeping frogs away from the surface with a mesh. The measure of hypoxia tolerance was the duration the animal remained active under the described conditions. Transition to inactivity was defined by cessation of movements and eye rolling. Immediately thereafter the jar was opened and the animal removed to air. In all cases, a few seconds after skin contact with air the frog opened its eyes and assumed a normal posture. Measurements were statistically processed using Statistica 8.0. The first frog sample was fairly uniform in erythrocyte size (21.85 ± 1.83 µm), indicating it consisted solely of diploids. The mean time spent in water was 342 s, with no significant deviations. In this sample a correlation between erythrocyte size and body length was established (Fig. 1). The Spearman correlation coefficient for these parameters is R = 0.42 (p = 0.036). No effect of erythrocyte size on hypoxia tolerance was recorded. Figure 1. Relationship between erythrocyte size and body length (first frog sample) The second sample comprised frogs with more variable erythrocyte sizes (17.65–30.51 µm). Five probable triploids (erythrocyte size > 26 µm) were identified (6). In the distribution of activity‑preserving time under artificial hypoxia, two peaks were observed (Fig. 2). Probable triploids displayed both relatively short (587 s) and very long (3060 s) survival times. As shown in Fig. 2, a factor approximately doubled the hypoxia endurance in some individuals. Since the second series of experiments was conducted in early autumn, this factor is likely physiological preparation for overwintering. Figure 2. Distribution of activity‑preserving time under hypoxia for the second frog sample The influence of erythrocyte size on hypoxia tolerance was not recorded (Fig. 3). Only a weak and statistically non‑significant relationship between erythrocyte size and hypoxia endurance was observed. Figure 3. Dependence of hypoxia‑exposure time on erythrocyte size. Two frog groups (according to our hypothesis) are shown separately Conclusions.
Despite the emergence of an unplanned factor (presumably overwintering preparation) in our experiments, we can state that no significant differences in hypoxia tolerance between diploid and triploid P. esculentus were found. We may anticipate the presence of specific compensatory mechanisms in triploids that mitigate cellular dimensional effects, warranting further investigation. The work was supported by the Fund for Fundamental, Applied and Exploratory Scientific Research of V.N. Karazin Kharkiv National University. References. 1. Parfenova I.A., Soldatov A.A. Erythrogram of circulating blood of scorpionfish under experimental hypoxia. // Marine Ecological Journal. – 2005. – Vol. IV, No.2. 2.
Shabanov D.A., Korshunov O.V., Kravchenko M.O. Which green frogs inhabit Kharkiv region? Terminological and nomenclatural aspects of the problem // Biology and Valiology. – Issue 11. – Kharkiv: HDPU, 2009. – pp. 164–125.
3. Shabanov D.A., Lytvynchuk S.N. Green frogs: life without rules or a special mode of evolution? // Nature. – 2001. – No.3. – pp. 29–36. 4. Cal R.M., Vidal S., et al. Effect of triploidy on turbot haematology. // Comparative Biochemistry and Physiology, Part A 141 (2005). 35–41. 5. George S.A., Michelle R. Lennartz. Methods for determining ploidy in amphibians: nucleolar number and erythrocyte size. // Experientia. – 1980. – pp. 687–688. 6. Plötner J. Die Westpaläarktichen Wasserfrösche. Bielefeld, 2005. 7. Polls Pelaz M., Graf J.-D. Erythrocyte size as an indicator of ploidy level in Rana kl. esculenta before and after metamorphosis. // Alytes 1988. – 7. – pp. 53–61. 8. Maxime V., Labbé L. The effect of ploidy and sexual maturation on the resistance of erythrocytes to haemolysis in rainbow trout (Oncorhynchus mykiss). // Aquaculture. – 2010. – 305. Do dimensional effects of diploid and triploid edible frogs cells influence on their resistance to hypoxia? Bondaryeva A.A., Makhnii T.I., Shabanov D.A. Diploids and triploids of Pelophylax esculentus inhabiting the green frogs diversity centre in the Siversky Donets basin are good model for study of influence of cell size on physiological processes. The hypothesis that triploids differ from diploids in their resistance to hypoxia was tested. This hypothesis was not confirmed. It was shown that the sizes of frog erythrocytes are positive related to their body sizes. In early autumn it is registered a significant increase in resistance to hypoxia in some frogs; presumably, this is related to their physiological preparation for wintering. Key words: edible frogs, erythrocytes, hypoxia, triploidy, dimensional effects of cells. " }