
Студентські роботи після практики II курсу – 2014
Artjukh O. A., Pereverzeva Ya. V., Brusianskyi I. I. Oпис брюшних і спинних узорів червоночеревих жабок Vakulenko M. V., Meleško E. V., Khazeyeva A. A. Дослідження структури популяційної системи зелених жаб (Pelophylax esculentus complex) Нижнього Добрицького ставку Зміївського району Харківської області.
Студентські роботи після практики II курсу – 2008
Студентські роботи після практики II курсу – 2009
Студентські роботи після практики II курсу – 2010
Студентські роботи після практики II курсу – 2011
Студентські роботи після практики II курсу – 2012 (I частина)
Студентські роботи після практики II курсу – 2012 (II частина)
Студентські роботи після практики II курсу – 2013
Студентські роботи після практики II курсу – 2015
Студентські роботи після практики II курсу – 2016
Студентські роботи після практики II курсу – 2017
Опис черевних і спинних візерунків червоночеревних жабок
Артюх О. А., Переверзева Я. В., Брусянський І. І.
Харківський національний університет імені В. Н. Каразина, біологічний факультет, кафедра зоології та екології тварин, пл. Свободи, 4, м. Харків, Україна, 61077
e‑mail: neagame123@gmail.com
Формалізований опис візерунків червоночеревних жабок Bombina bombina (Linnaeus, 1761) може використовуватись з метою індивідуального маркування особин і для фенетичного вивчення популяцій (визначення родинних зв’язків, меж популяцій та їх груп). Матеріалом для роботи послужили 23 статевозрілих і 1 ювенільна особина, зібрані в поймі річки Північного Донця в околицях біологічної станції біологічного факультету ХНУ імені В. Н. Каразина в Національному природному парку «Гомільшанські ліси» у першій декаді липня 2014 р.
Для отримання якісних зображень дорзального і вентрального візерунка жабок притискали до скла офісного сканера білою ганчіркою, після чого проводили сканування. Вивчені особини випущені в місця збору.
У досліджуваній вибірці сканограм рисунка ми виділили по 3 ознаки (кожна з яких могла приймати кілька станів) з вентральної і з дорзальної сторони для кожної особини. Для дорзальної сторони були виділені такі ознаки: ознака 1 – спинні плями бобовидної форми (0 – відсутність бобовидних плям; 1 – 1 бобовидна пляма; 2 – 2 плями; 3 – 3 плями; 4 – 4 плями), ознака 2 – зелений колір на спині (0 – немає; 1 – є), ознака 3 – плями в області голови (1 – Y‑подібна пляма; 2 – рідкісна Y‑подібна пляма; 3 – X‑подібна пляма; 4 – горизонтальна смуга між очима; 5 – рідкісна смуга між очима, від 2 до 4 плям). З вентральної сторони були виділені такі ознаки: ознака 4 – яскраво‑червоні плями в зоні поясу верхніх кінцівок (1 – 8 плям; 2 – менше 8 плям), ознака 5 – плями на брюхові та голові (1 – дрібні; 2 – великі; 3 – дрібні і великі), ознака 6 – зона переходу від брюшка до нижніх кінцівок (1 – 2 витягнуті плями; 2 – 3 плями; 3 – 4 і більше 4‑х плям).
Кодування візерунка особини складається з двох груп (для дорзальної і вентральної сторони тіла), кожна з яких складається з трьох чисел, що відображають стани виділених нами ознак. Наприклад, код дорзального візерунка 1.1.5 означає, що жабка за 1‑й ознакою має 1 бобовидну пляму, за 2‑й ознакою – зелений колір спини, а за 3‑й – рідкісну смугу між очима (від 2 до 4 плям). Для вентральної сторони код 1.1.2 має таку розшифровку: за 4‑й ознакою – наявність плям у зоні поясу верхніх кінцівок, за 5‑й ознакою – дрібні плями на брюхові та голові, за 6‑й – 3 плями в зоні переходу від брюшка до нижніх кінцівок. Відповідно до цих кодів слід організувати комп’ютерну базу сканограм або фотографій з можливістю формування підборок наявних зображень, що володіють певним станом будь‑якої з використаних ознак. Остаточну ідентифікацію слід проводити шляхом порівняння двох зображень або зображення і досліджуваної особини.
Розроблена нами методика може бути застосована для індивідуального маркування або вивчення локальної популяційної структури жабок.
Summary. The subject of work is Bombina bombina (Linnaeus, 1761). The material has been collected in the vicinities of the biological station of the Kharkiv National University named by V. N. Karazin in the National park "Gomilshansky Forests" and consist of 23 mature individuals and 1 juvenile.There was developed a method of encoding. The code consists of three numbers, each number indicates a certain state of the trait.
Автори висловлюють подяку науковому керівнику доценту Д. А. Шабанову.
Артюх О. А., Переверзева Я. В., Брусянський І. І. Опис черевних і спинних візерунків червоночеревних жабок // «Біологія: від молекули до біосфери». Матеріали IX Міжнародної конференції молодих учених (18–20 листопада 2014 р., м. Харків, Україна). – Харків: ФОП Шаповалова Т. М., 2014. – С. 122.
Дослідження структури популяційної системи зелених жаб (Pelophylax esculentus complex) Нижнього Добрицького ставку Зміївського району Харківської області
Вакуленко М. В., Мелешко Є. В., Хазеева А. А.
Харківський національний університет імені В. Н. Каразина, біологічний факультет, пл. Свободи, 4, м. Харків, Україна
e‑mail: mashavmv@rambler.ru
Групу зелених жаб (Pelophylax esculentus complex) складають Pelophylax lessonae (Camerano, 1882), Pelophylax ridibundus (Pallas, 1771) і їх геміклональний гібрид Pelophylax esculentus (Linnaeus, 1758). Спільно живі та розмножуються представники цього комплексу утворюють геміклональні популяційні системи (ГПС). У даній роботі вивчено склад ГПС зелених жаб Нижнього Добрицького ставку (територія НПП «Гомільшанські ліси», Зміївський р‑н, Харківська обл.), для якої раніше була зафіксована висока частка триплоїдних гібридів (Бондарева, 2012; Мелешко тощо, 2014). Дослідження цієї ГПС з використанням маркування (шляхом ампутації пальців) ведуться з 2011 р. Завдання роботи – вивчення складу та оцінка чисельності зазначеної ГПС, а також порівняння отриманих даних з результатами досліджень 2013 р.
Вивчена випадкова вибірка, зібрана 7 липня 2014 р. Жабок ловили руками в темний час доби з використанням ліхтарика. Видову та статеву приналежність визначали за комплексом морфологічних ознак (Коршунов, 2010). Плоїдність визначали за розміром еритроцитів (Бондарева, 2012). Кров’яні мазки фотографували під великим збільшенням мікроскопа USB‑камерою, довжину еритроцитів вимірювали за допомогою програми PDF XChangeViewer, отримані результати обробляли в Statistica 8.0. Чисельність вивченої ГПС оцінювали за Пітерсоном як відношення об’єму вибірки 2014 р. до частки зібраних особин, маркованих у 2013 р., з урахуванням поправки Бейлі (метод маркування і повторного відлову).
Досліджено 51 особину, з яких P. esculentus – 46 екз. (90 %), а P. ridibundus – 5 (10 %). Серед P. esculentus виявлено 35 самців (76 %), 8 самок (17 %) і 3 незрілі особини (7 %). Частка триплоїдних особин склала 8 % від усієї вибірки (4 особини: 2 самці, 1 самка і 1 незріла особина). Серед P. ridibundus виявлено 4 самки і 1 незріла особина. Порівняно з результатами відлову 2013 р. (Михайленко тощо, 2013), відносна частка триплоїдних гібридів значно (p = 0,02) зменшилася. 2 з 76 маркованих у 2013 р. особин потрапили у вибірку 2014 р. Розрахована чисельність ГПС склала 1317 ± 639 особин. 42 марковані особини були випущені в Нижній Добрицький ставок.
Таким чином, ГПС зелених жаб Добрицького ставку відноситься до REt‑типу, у ній переважають диплоїдні самці P. esculentus. Зафіксовано зменшення частки тріплоїдів порівняно з минулим роком (p = 0,02). Причини спостережуваних змін не можуть бути встановлені використаними в роботі методами і потребують окремого вивчення. Подальший моніторинг даної ГПС представляє інтерес у зв’язку з виявленими змінами її складу.
Summary. The hemiclonal population system of green frogs from Lower Dobristky pond (National Park “Gomilshansky lissy”) was studied. Sexual and species structure was determined. The diploid males P. esculentus prevailed in this population system (the population type is REt). The portion of parental species (P. ridibundus) is 10 %. The portion of male in sample is 69 % (35 individuals of 51). The portion of female is 23 % (12 individuals), the portion of juvenile is 8 % (4 individuals). The number of triploids was shown (p=2 %) to decrease with respect to 2013 data (Mikhajlenko et al., 2013). The estimated population size is 1317±639.
Автори висловлюють подяку доценту кафедри зоології та екології тварин ХНУ імені В. Н. Каразина Д. А. Шабанову за наукове керівництво, а також А. В. Коршунову і Є. Є. Усовій за допомогу у зборі жаб.
Вакуленко М. В., Мелешко Є. В., Хазеева А. А. Дослідження структури популяційної системи зелених жаб (Pelophylax esculentus complex) Нижнього Добрицького ставку Зміївського району Харківської області // «Біологія: від молекули до біосфери». Матеріали IX Міжнародної конференції молодих учених (18–20 листопада 2014 р., м. Харків, Україна). – Харків: ФОП Шаповалова Т. М., 2014. – С. 126‑127.
Порівняння довжини та маси тіла головастиків зелених жаб (Pelophylax esculentus complex), що розвиваються в поймі Північного Донця та в Іськовому ставку
Дрогваленко Н. А., Черненко А. В.
ХНУ імені В. Н. Каразина, біологічний факультет, кафедра зоології та екології тварин, пл. Свободи, 4, м. Харків, Україна
e‑mail: Jaegernicholaus@gmail.com
Різниці в умовах проживання головастиків гібридного комплексу Pelophylax esculentus complex можуть впливати на динаміку їх росту та розвитку. У зв’язку з цим нами було досліджено розвиток головастиків, взятих з різних водойм Північно‑Донецького центру різноманітності зелених жаб.
Ми вивчили дві вибірки: 65 головастиків з Північного Донця (N 49,635; E 36,338; спіймано 01.07.14) і 53 головастика з Іськового ставку (N 49,627; E 36,284; спіймано 07.07.14). Головастиків ловили «косячими» рухами гідробіологічної сітки (комірка 7 мм) у водній рослинності на мілководді. У місцях збору нерестяться і представники батьківського виду Pelophylax ridibundus, і міжвидові гібриди Pelophylax esculentus, тому можна припустити, що досліджені нами вибірки головастиків представляли суміш різних форм представників Pelophylax esculentus complex.
У головастиків вимірювали довжину тіла (від кінчика морди до основи хвоста) автоматичним штангенциркулем (точність 0,01 мм) і масу тіла електронними вагою (точність 0,01 г). Виділяли 10 стадій розвитку задніх кінцівок (Виноградська тощо, 2010). Дисперсний аналіз проводили за допомогою програми Statistica.
У вибірках переважали головастики 7‑10‑х стадій розвитку задніх кінцівок. Головастики з пойми річки незначно (p = 0,22) перевищували головастиків зі ставку, що перебувають на тих самих стадіях розвитку, за довжиною їх тіла, і значно (p = 0,04) перевищували їх за масою тіла. Наші дані відповідають результатам аналогічної роботи, виконаної 2 роки тому (Губенко, Михайлова, 2012), що підтверджує стійкість зафіксованого ефекту.
Summary. Weight, length and developmental stage of hind limbs of tadpoles from two population of hybridogeneous comlex Pelophylax esculentus complex were studied. Tadpoles of bottomland of Siversky Donets have bigger average length and average weight.
Автори висловлюють подяку науковому керівнику Шабанову Д. А., доценту кафедри зоології та екології тварин ХНУ імені В. Н. Каразина
Дрогваленко Н. А., Черненко А. В. Порівняння довжини та маси тіла головастиків зелених жаб (Pelophylax esculentus complex), що розвиваються в поймі Північного Донця та в Іськовому ставку // «Біологія: від молекули до біосфери». Матеріали IX Міжнародної конференції молодих учених (18–20 листопада 2014 р., м. Харків, Україна). – Харків: ФОП Шаповалова Т. М., 2014. – С. 128‑129.
Порушення фертильності у міжвидових гібридів зелених жаб з Північно‑Донецького центру різноманітності Pelophylax esculentus complex
Макарян Р. М., Шейко В. П.
Харківський національний університет імені В. Н. Каразина, біологічний факультет, пл. Свободи 4, м. Харків, Україна
e‑mail: inessasheiko@mail.ru
У склад гібридного комплексу зелених жаб (Pelophylax esculentus complex) входять три основні форми. Це ставкові жаби Pelophylax lessonae (Camerano, 1882), озерні жаби Pelophylax ridibundus (Pallas, 1771) і їх геміклональні гібриди, їстівні жаби Pelophylax esculentus (Linnaeus, 1758). У ході дослідження реакції самців зелених жаб на ін’єкцію сурфагону (синтетичного аналогу гонадотропін‑рілізинг гормону луліберіна), які виконувала протягом літнього сезону 2014 р. А. А. Боброва, були виявлені самці зелених жаб, які не реагували на гормональну стимуляцію. Завдання нашої роботи полягало у порівнянні розмірів яєчок у таких особин, і у жаб, що демонстрували нормальну реакцію на гормон. Крім того, ми перевіряли наявність активних сперматозоїдів у самих яєчках P. ridibundus і P. esculentus.
Ми досліджували 22 самці зелених жаб, серед яких 8 Pelophylax ridibundus і 14 Pelophylax esculentus. Тварини зібрані в околицях біостанції ХНУ імені В. Н. Каразина (Зміївський р‑н, Харківська обл.). Ми вивчили 11 P. esculentus і 2 P. ridibundus з числа тих, що не дали відповіді на гормональну стимуляцію, а також 3 P. esculentus і 6 P. ridibundus, випадково зібраних у природних місцевостях. Жабок наркозували, розривали їм головний і спинний мозок, відкривали черевну порожнину і витягали яєчка. Жабу (спинною стороною вниз) і її відокремлені яєчка розкладали на склі офісного сканера А4, після чого сканували з роздільною здатністю 600 dpi. Вимірювання довжини тіла жаби та довжини яєчок проводили за фотографіями за допомогою PDF XChangeVIEWER з використанням відсканованої мірної шкали. Для визначення наявності активних сперматозоїдів розрізали один з яєчок ножицями на кілька шматочків, які поміщали в річну воду. Через кілька хвилин отриману суспензію мікроскопували і реєстрували наявність або відсутність сперматозоїдів.
У P. esculentus яєчка невеликого розміру (середня довжина – 6 мм), яєчка у P. ridibundus більші (середня довжина – 8,2 мм). Серед 14 P. esculentus частка особин з активними сперматозоїдами склала 28,6 % (4 особини), а серед 8 P. ridibundus – 75 % (6 особин).
У P. ridibundus яєчка довжиною від 8 до 11 мм містили активні сперматозоїди, а у особин P. esculentus при довжині яєчок від 8,4 до 10 мм активні сперматозоїди не виявлені.
Ймовірно, причиною більш суттєвих порушень фертильності у P. esculentus є порушення їх гаметоогенезу внаслідок міжвидової гібридизації.
Summary. It was investigated impaired fertility in interspecific hybrids of green frogs from Donetskiy center of diversity Pelophylax esculentus complex. We identified the presence of active sperm in the suspension of the cut of the testes, as well as the size of the testes. It was found impaired fertility of P. esculentus in our results of investigations.
Автори висловлюють глибоку подяку А. А. Бобровій за наданий матеріал і допомогу в роботі, а також доценту Д. А. Шабанову за наукове керівництво і допомогу в інтерпретації результатів.
Макарян Р. М., Шейко В. П. Порушення фертильності у міжвидових гібридів зелених жаб з Північно‑Донецького центру різноманітності Pelophylax esculentus complex // «Біологія: від молекули до біосфери». Матеріали IX Міжнародної конференції молодих учених (18–20 листопада 2014 р., м. Харків, Україна). – Харків: ФОП Шаповалова Т. М., 2014. – С. 129‑130.
Імітаційне моделювання самоорганізації в біологічних системах на прикладі хорового співу зелених жаб
Малахова Д. О.
Обласна спеціалізована школа‑інтернат «Обдарованість» Харківської обласної ради
e‑mail: sunlilika@i.ua
Імітаційне моделювання може використовуватись для перевірки того, чи виникають властивості цілісних систем із передбачуваного набору властивостей її елементів. Дана робота має перевірити припущення щодо механізмів хорового співу зелених жаб.
Спів жаб часто здавався загадковим, таким, що потребує наявності «диригента» для пояснення злагодженості хору. Ми перевіряли, чи можна пояснити цю злагодженість, застосовуючи гідравлічну модель інстинктивної поведінки, яку запропонував Конрад Лоренц. Згідно з нею, реалізація певної форми поведінки пояснюється ростом відповідної мотивації (яку можна порівняти з рідиною, що накопичується або витрачається); під час реалізації цієї поведінки мотивація знижується. З наростанням мотивації відповідна форма поведінки або запускається специфічним стимулом, або починається самовільно.
Ми припустили, що поведінку кожної жаби можна описати наступними параметрами:
— В (від англ. begin), поріг заспіву (рівень мотивації, при досягненні якого жаба, яка мовчить, починає співати);
— E (від англ. end), поріг замовкання (рівень мотивації, при зниженні до якого замовкає жаба, яка співає);
— A (від англ. accumulation), швидкість зростання мотивації у особини, що мовчить;
— S (від англ. spending), швидкість зниження мотивації у особини, що співає;
— P (від англ. provocation), ефективність голосової провокації (зниження порогу заспіву, яке прискорює спів іншої жаби).
Жаба, що має рівень мотивації L1 (від англ. level), мовчить, якщо вона мовчала на попередньому кроці моделі та L1 < B (або L1 < B–P, якщо співають інші жаби). Вона заспіває через такий час (кількість кроків моделі) t1, що L1 + t1×A = B (у разі, якщо співають інші жаби, L1 + t1×A = B–P). Жаба, що почала співати (наприклад, під час співів інших жаб, із рівнем мотивації L2 = B–P), замовчить через такий час t2, що L2 – t2×S = E (і у тому разі, якщо інші жаби співають, і в тиші). Зрозуміло, що ці припущення мають сенс, якщо B > E, а також P < B–E.
Ми створили модель, що відображає ці припущення, у Microsoft Office Excel. Вона імітує поведінку 28 розташованих колом жаб, на кожну з яких впливає спів 10 її найближчих сусідів. Модель розміщено у відкритому доступі: http://dspace.univer.kharkov.ua/handle/123456789/10058.
Експерименти з моделлю показали, що прийнятих припущень достатньо, щоб пояснити появу періодичного хорового співу. Ми записали хоровий спів зелених жаб, яких утримують у лабораторії популяційної екології амфібій ХНУ імені В. Н. Каразина. Для цього ми записували усі звуки у лабораторії після завершення роботи співробітників 18 жовтня 2014 р. Періодичність «включення» хору жаб склала 50 хв.Ми підібрали такі значення початкових параметрів, при яких модель продукує аналогічну динаміку хорового співу (B=56; E=18; A=1; S=24; P=2; крок моделі відповідає 1 хвилині). Існують значення початкових параметрів, при яких хор розбивається на дві частини, що співають поперемінно.
На наступному етапі нашої роботи ми плануємо зареєструвати динаміку поодинокого та хорового співу зелених жаб у природних умовах, оцінити рівень індивідуальної мінливості мотивації до співу та встановити відповідні цим даним параметри моделі.
Summary. The goal of this model is to explain the harmony in the chorus of frogs. We decided to simulate using Excel a real chorus of frogs with stewed parameters that are proportional relationship. Through experiments with the model and using the true record of singing frogs, we proved that our assumptions sufficient to explain the frequency of thisoperation.
Автор щиро дякує науковому керівнику, доценту ХНУ імені В. Н. Каразіна Д. А. Шабанову за ідею даної роботи та допомогу в її втіленні у модель.
Малахова Д. О. Імітаційне моделювання самоорганізації в біологічних системах на прикладі хорового співу зелених жаб // «Біологія: від молекули до біосфери». Матеріали IX Міжнародної конференції молодих учених (18–20 листопада 2014 р., м. Харків, Україна). – Х.: ФОП Шаповалова Т. М., 2014. – С. 130-131.
Исследование гібридного комплекса Carassius пруда в Коряковом яру (Харьковская обл., Украина)
Мирославская М. А., Сухов В. А., Шлахтер М. Л.
Харківський національний університет імені В. Н. Каразіна, кафедра зоології та екології тварин, пл. Свободи, 4, Харків, Україна
e‑mail: marina.miroslavskaya@yandex.ua
Гібридні диплоїдно‑поліплоїдні комплекси є моделлю еволюційно‑генетичних та екологічних досліджень. Вивчення гібридних комплексів карасей представляє особливу важливість також і для природоохоронної справи. Для басейну річки Північний Донец дослідження гібридних комплексів карасей раніше вже проводилися (Кулиш, 2013), за їх результатами можна зробити висновок про високу ймовірність формування унікальних поселень карасей у окремих водоймах.
Ми вивчили 63 карасі, відловлених у період з 25.06.2014 по 27.06.2014 пастками типу «конус» у ставку в Коряковому яру (Харківська обл., Зміївський р‑н, околиці с. Гайдари). Визначали 10 ознак: наявність плями на хвості, форма спинного плавника, форма анального плавника, колір плавників, форма голови, колір луски, колір очей, кількість жабрових трубочок, кількість шипиків на спинному та анальному плавниках, колір підшкірної обкладки. В залежності від стану цих ознак вивчених особин відносили до одного з морфотипів: Carassius cf. carassius або Carassius cf. gibelio на підставі опису двох видів у «Фауні України» (Мовчан, Смирнов, 1982 р.).
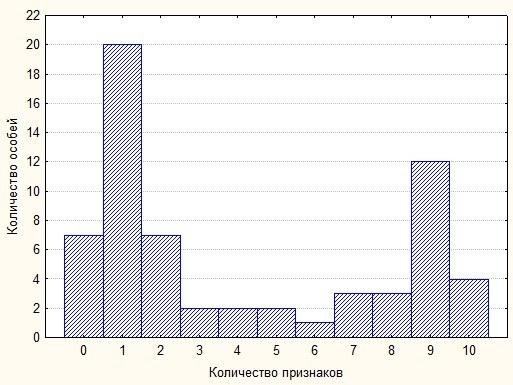
Рис. 1
На рис.1 подано зареєстрований розподіл особин за кількістю ознак, характерних для Carassius cf. gibelio (нехарактерних для Carassius cf. carassius). Розподіл виявився неперервним і двовершинним. Піки розподілу припадають на особин, що відрізняються від типового опису чистого морфотипу за однією ознакою. На підставі цього результату ми можемо висловити припущення, що в даному ставку відбувається гібридизація, що призводить до перерозподілу ознак між різними морфотипами.
Розподіл за довжиною тіла карасей у віці 3‑х років (вік визначали за кільцями на лусці) значно відрізняється від нормального (p=0,0016) і має двовершинний характер. Довжина тіла не залежить від морфотипу. Це свідчить про неоднорідність групи і дозволяє припустити, що в її склад входять швидкоростучі та повільноростучі особини або ді‑ і поліплоїди, оскільки плоїдність також може впливати на розміри організму, а утворення поліплоїдів часте явище для гібридних комплексів.
Summary. Identified 10 characteristics, depending on the state of these characteristics of the studied individuals was attributed to one of the morphotypes: Carassius cf. carassius or Carassius cf. gibelio. The distribution of the length of the body carp at the age of 3 years is significantly different from normal and has a bimodal nature, it demonstrates the heterogeneity of the group and suggests that it consists of fast‑growing and slow‑growing individuals or di‑ and polyploids.
Автори висловлюють подяку Шабанову Д. А. і Гончарову Г. Л. за наукове керівництво, відлов матеріалу та допомогу у статистичній обробці даних.
Мирославська М. А., Сухов В. А., Шлахтер М. Л. Дослідження гібридного комплексу Carassius пруда в Коряковому яру (Харківська обл., Україна) // «Біологія: від молекули до біосфери». Матеріали IX Міжнародної конференції молодих учених (18–20 листопада 2014 р., м. Харків, Україна). – Х.: ФОП Шаповалова Т. М., 2014. – С. 132-133.
Устойчивість до гіпоксії у представників Pelophylax esculentus complex
Парамонова В. В., Ковалев В. В.
Харківський національний університет імені В. Н. Каразіна, біологічний факультет, кафедра зоології та екології тварин, пл. Свободи, 4, м. Харків, Україна, 61022
e‑mail: vika_paramonova@mail.ua
Представники гібридогенного комплексу зелених жаб (Pelophylax esculentus complex) – зручний об’єкт для дослідження впливу розміру клітин на фізіологічні процеси. У Північно‑Донецькому центрі різноманіття зелених жаб міжвидові гібриди, Pelophylax esculentus (Linnaeus, 1758), представлені ді‑ і триплоїдами. Розмір еритроцитів (і, ймовірно, багатьох інших клітин) у триплоїдів збільшений, а маса і розмір тіла відповідають таковим у диплоїдів (Шабанов, Литвинчук, 2010). Збільшення клітин, зменшення їх відносної поверхні та кількості має призводити до ускладнення газообміну; цьому припущенню відповідає вища активність ферментів анаеробного розщеплення глюкози (Бондарева та ін., 2013). Ми перевіряли гіпотезу, що триплоїдні жаби мають відрізнятися від диплоїдних стійкістю до гіпоксії.
Жаб ловили в кінці червня 2014 року в околицях біостанції ХНУ імені В. Н. Каразіна в поймі р. Північний Донец і на Нижньому Добрицькому ставку (НПП «Гомольські ліси»). Ми визначили стійкість до гіпоксії 33 особин (25 самців і 8 самок): 5 триплоїдних P. esculentus, 18 диплоїдних P. esculentus, 10 – Pelophylax ridibundus (Pallas, 1771), представників батьківського виду. Належність до батьківського виду або гібридів визначали за комплексом зовнішніх ознак (Атемасова та ін., 2008), плоїдність — за розмірами еритроцитів (Бондарева та ін., 2012). Жаб розміщували в банку з дегазованою водою без доступу до повітря і визначали час до втрати активності. Після експерименту жаб випустили у місцях їхнього вилову. Результати обробляли в програмі Statistica 7 з використанням багатофакторного дисперсійного аналізу (рис. 1).
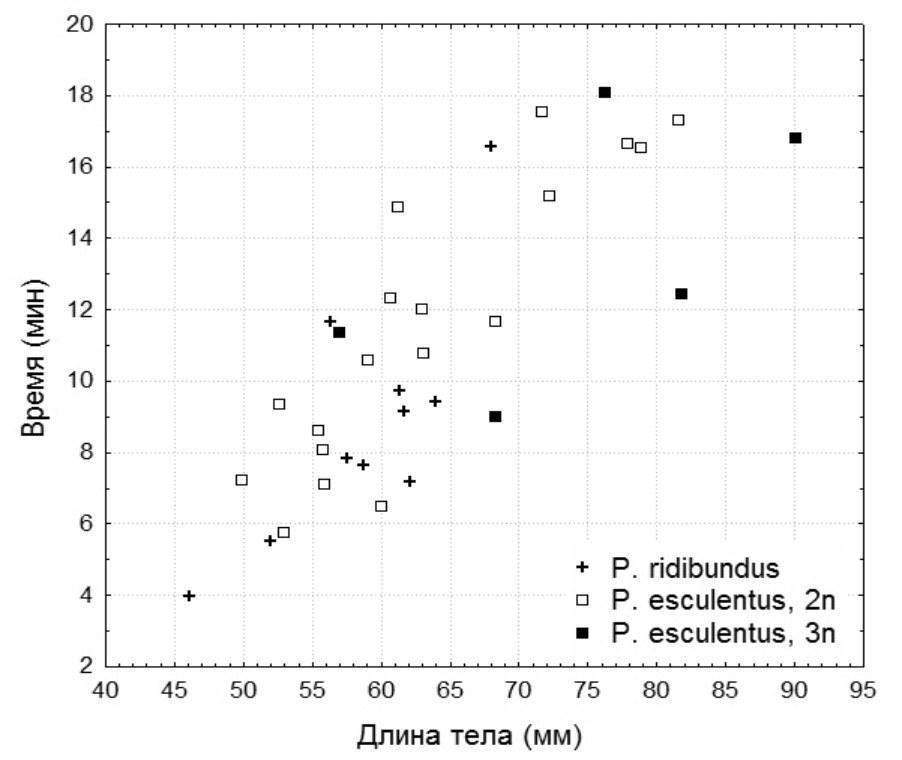
Рис. 1. Залежність стійкості до гіпоксії від довжини тіла зелених жаб. На стійкість до гіпоксії з розглянутих факторів значно впливає лише довжина тіла. При збільшенні довжини тіла збільшується стійкість до гіпоксії. Відмінності у стійкості ді‑ і триплоїдних P. esculentus, між P. ridibundus і P. esculentus (рис. 1), а також між самками і самцями незначущі.
Хоча на підставі раніше наявних даних можна припустити, що фізіологічні особливості реакції ді‑ і триплоїдних P. esculentus на гіпоксію можуть бути різними, наші результати показують, що зміна розмірів клітин у триплоїдів ефективно компенсується. Механізми такої компенсації потребують додаткового вивчення.
Summary. The influence of ploidy on the hypoxia resistance Pelophylax esculentus was investigated. The time what frog spent in degassed water to moment of loss the activity was the indicator of resistance. The results showed only one dependence: dependence of the resistance to body length.
Автори вдячні доценту Шабанову Д. А. за наукове керівництво та допомогу в роботі.
Парамонова В. В., Ковалев В. В. Устойчивість до гіпоксії у представників Pelophylax esculentus complex // «Біологія: від молекули до біосфери». Матеріали IX Міжнародної конференції молодих учених (18–20 листопада 2014 р., м. Харків, Україна). – Х.: ФОП Шаповалова Т. М., 2014. – С. 133-134.
Вивчення особливостей криків вивільнення зелених жаб із Сіверсько-Донецького центру різноманіття Pelophylax esculentus complex
Тимченко К. М., Федорова А. О.
Харківський національний університет імені В. Н. Каразіна, біологічний факультет, кафедра зоології та екології тварин, пл. Свободи, 4, м. Харків, 61077
e‑mail: karisha_139@mail.ru
В околицях біостанції ХНУ імені В. Н. Каразіна (с. Гайдари, Зміївський р‑н, Харківська обл.) живуть міжвидові гібриди зелених жаб Pelophylax esculentus (Linnaeus, 1758) і представники одного з батьківських видів – Pelophylax ridibundus (Pallas, 1771). Безхвості амфібії належать до небагатьох груп тварин, при описі різноманіття яких корисні методи біоакустики. Серед криків безхвостих найбільшу складність і інформативність мають брачні заклики самців. Відмінності таких закликів описані і використовуються як у таксономічних цілях, так і для польового визначення форм жаб. Менш вивчені – крики вивільнення, що видаються представниками обох статей при спробі взяти їх у амплексус.
У даній роботі ми порівнювали крики вивільнення самців і самок P. ridibundus і P. esculentus. Для цього ми записали крики вивільнення 62 зелених жаб (8 самців і 17 самок P. ridibundus, 30 самців і 7 самок P. esculentus). Жаб утримували за тулуб з боків, записуючи їх реакцію на відео за допомогою цифрового фотоапарата. Осцилограми звукових сигналів аналізували в програмі Audacity 2.0.5. Для кожної особини аналізували по 4 циклічних крика. Для кожного циклу вимірювали загальну тривалість, тривалість активної фази, кількість імпульсів у крику, а також тривалість імпульсу, тривалість активної фази імпульса і кількість імпульсів у імпульсі на початку, в середині і в кінці крика. Аналіз результатів проводили в програмі Statistica 8.0 (рис. 1).
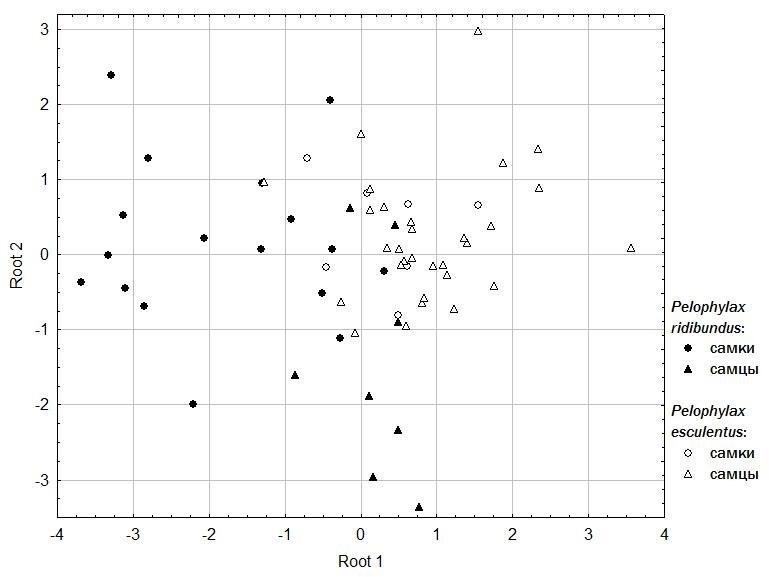
Рис. 1. Результати дискримінантного аналізу записів голосів жаб (за використаними 12 параметрами осцилограм аудіозаписів). Крики вивільнення самців і самок батьківського виду, P. ridibundus, утворюють дві слабо пересічні групи. У міжвидових гібридів, P. esculentus, статеві відмінності значно втрачені.
В цілому за результатами дискримінантного аналізу правильно класифіковано 77 % особин.
З нашої точки зору, розпочата робота потребує продовження і розвитку. Необхідно включити в аналіз крики вивільнення другого батьківського виду, Pelophylax lessonae (Camerano, 1882), продовжити роботу з пошуком характерних для різних груп жаб ознак їх криків, а також вивчення змінності цих ознак.
Summary: In the vicinity of the biological station V. N. Karazin Kharkiv National University live interspecific hybrids of green frogs Pelophylax esculentus (Linnaeus, 1758) and the representatives of one of the parental species – Pelophylax ridibundus (Pallas, 1771). The main goal of the research was to determine the differences between the releasing calls of males and females of the two species. Differences between males and females calls of P. ridibundus were identified. Differences between males and females calls of P. esculentus were not found.
Автори висловлюють подяку науковому керівнику, доценту Д. А. Шабанову.
Тимченко К. М., Федорова А. О. Вивчення особливостей криків вивільнення зелених жаб із Сіверсько-Донецького центру різноманіття Pelophylax esculentus complex // «Біологія: від молекули до біосфери». Матеріали IX Міжнародної конференції молодих учених (18–20 листопада 2014 р., м. Харків, Україна). – Х.: ФОП Шаповалова Т. М., 2014. – С. 134-135.