
Еволюція еукаріот
Саме появлення еукаріотичної клітини є одним із найзначущіших подій біологічної еволюції. Відмінність еукаріотичних організмів від прокаріотичних полягає у більш досконалій системі регуляції геному. Завдяки цьому зросла пристосованість одноклітинних організмів, їх здатність адаптуватися до змінних умов середовища без внесення успадкованих змін у геном. Завдяки можливості адаптуватися еукаріоти змогли стати багатоклітинними – у багатоклітинному організмі клітини з одним і тим же геномом, залежно від умов, утворюють зовсім різні за морфологією та функцією тканини.
Цей ароматофоз відбувся на межі Архея і Протерозою (2,6–2,7 млрд років тому), що визначили за біомаркерами – залишками стероїдних сполук, властивих лише еукаріотичним клітинам. Поява еукаріот збігається за часом з кисневою революцією.
Вважається загальновизнаним, що еукаріоти з’явилися в результаті симбіозу кількох різновидів прокаріот. Очевидно, мітохондрії походять від альфа‑протеобактерій (аеробних еубактерій), пластиди – від ціанобактерій, а цитоплазма – від невідомої архебактерії. Поки що немає загальноприйнятої теорії виникнення ядра, цитоскелету, а також жгутиків. Гіпотези виникнення життя на Землі не внесли ясності у питання про виникнення клітини. Якщо про походження прокаріот практично немає жодних правдоподібних гіпотез, що описують їхнє виникнення, то щодо походження еукаріотичних клітин існує кілька точок зору.
Основні гіпотези походження еукаріот:
1. Симбіотична гіпотеза базується на двох концепціях. За першою з цих концепцій найфундаментальніше розмежування в живій природі – це розмежування між бактеріями та організмами, що складаються з клітин з істинними ядрами – протистами, тваринами, грибами та рослинами. Друга концепція полягає в тому, що джерелом деяких частин еукаріотичних клітин була еволюція симбіозів – формування постійних асоціацій між організмами різних видів. Припускається, що три класи органел – мітохондрії, реснички та фотосинтезуючі пластиди – походять від вільно живих бактерій, які в результаті симбіозу були включені до складу клітин прокаріот‑хазяїв. Ця теорія в значній мірі спирається на неодарвінські уявлення, розвинуті генетиками, екологами, цитологами, які пов’язали Менделеївську генетику з Дарвінською ідеєю природного відбору. Вона також спирається на молекулярну біологію, зокрема дані про структуру білків і послідовність амінокислот, на мікропалеонтологію, що вивчає найраніші сліди життя на Землі, і на фізику та хімію атмосфери, оскільки ці науки мають відношення до газів біологічного походження.
2. Інвагінальна гіпотеза стверджує, що предковою формою еукаріотичної клітини був аеробний прокаріот. Усередині знаходилося кілька геномів, прикріплених до клітинної оболонки. Корпускулярні органели та ядро виникли шляхом втискування і відшнуровування ділянок оболонки з подальшою функціональною спеціалізацією в ядро, мітохондрії, хлоропласти. Потім у процесі еволюції відбулося ускладнення ядерного геному і з’явилася система цитоплазматичних мембран. Ця гіпотеза пояснює наявність у оболонках ядра, мітохондрій, хлоропластів двох мембран. Однак вона стикається з труднощами у поясненні різниць у деталях процесу біосинтезу білка в корпускулярних органелах і цитоплазмі еукаріотичної клітини. У мітохондріях і хлоропластах цей процес точно відповідає такому в сучасних прокаріотичних клітинах.
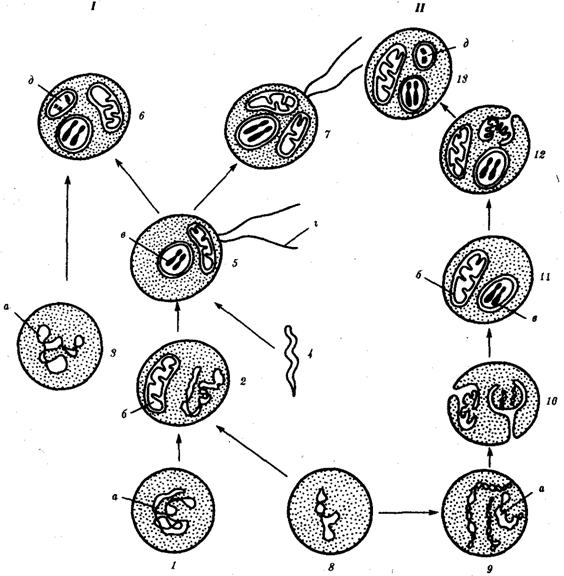
Походження еукаріотичної клітини згідно симбіотичної (I) та інвагінальної (II) гіпотез:
1 — анаеробний прокаріот (клітина‑хазяїн), 2 — прокаріоти, що мають мітохондрії, 3 — синьо‑зелена водорість (презумптивний хлоропласт), 4 — спірохетоподібна бактерія (презумптивний джгутик), 5 — примітивний еукаріот з джгутиком, 6 — рослинна клітина, 7 — тваринна клітина з джгутиком, 8 — аеробний прокаріот (презумптивна мітохондрія), 9 — аеробний прокаріот (клітина‑родоначальниця згідно гіпотези II), 10 — інвагінування клітинної оболонки, що дало ядро і мітохондрії, 11 — примітивний еукаріот, 12 — втискування клітинної оболонки, що дало хлоропласт, 13 — рослинна клітина; а — ДНК прокаріотичної клітини, б — мітохондрія, в — ядро еукаріотичної клітини, г — джгутик, д — хлоропласт.
Наявних даних поки недостатньо, щоб надати перевагу якійсь одній з гіпотез або розробити нову, яка задовольнила б більшість вчених, але в останні роки вдалося переконливо довести симбіогенетичну теорію походження еукаріотичної клітини.
Еволюційні можливості клітин еукаріотичного типу вищі, ніж прокаріотичного. Провідна роль тут належить ядерному геному еукаріот, який перевершує за розміром геном прокаріот. Важливі відмінності полягають у диплоїдності еукаріотичних клітин завдяки наявності в ядрах двох комплектів генів, а також у багатократному повторенні деяких генів.
Ускладнюється механізм регуляції життєдіяльності клітини, що проявилося у збільшенні відносної кількості регуляторних генів, заміні кільцевих «голих» молекул ДНК прокаріот хромосомами, у яких ДНК з’єднана з білками.
Аеробне дихання також послужило передумовою для розвитку багатоклітинних форм. Самі еукаріотичні клітини з’явилися на Землі після того, як концентрація O₂ в атмосфері досягла 1 % (точка Пастера). І ця концентрація – необхідна умова аеробного дихання.
Відомо, що кожна еукаріотична клітина містить геноми різного походження: у клітинах тварин і грибів це геноми ядра і мітохондрій, а в клітинах рослин – також і пластид. Невелика кільцева ДНК міститься і в базальному тілу джгутиків еукаріотичних клітин.
Згідно методу молекулярних годин еукаріоти виникли одночасно з прокаріотами. Але очевидно, що протягом значної частини історії Землі домінували прокаріоти. Перші клітини, які відповідають еукаріотичним розмірам (акритархи), мають вік 3 млрд років, проте їхня природа досі залишається неясною. Майже беззаперечні залишки еукаріот мають вік близько 2 млрд років. І лише після кисневої революції на поверхні планети склалися сприятливі умови для еукаріот (близько 1 млрд років тому).
Швидше за все, основним предком еукаріотичних клітин стали архебактерії, які перейшли до живлення шляхом поглинання харчових часток. Зміна форми клітини, необхідна для такого поглинання, забезпечувала цитоскелет, що складається з актина і міозину. Спадковий апарат такої клітини перемістився в глибину від її змінної поверхні, зберігши при цьому зв’язок з мембраною. І саме це стало причиною виникнення ядерної оболонки з ядерними порами.
Поглинені клітиною‑хазяїном бактерії могли продовжити своє існування всередині неї. Так, предками мітохондрій стала група фотосинтезуючих бактерій – пурпурні альфа‑протеобактерії. Усередині клітини‑хазяїна вони втратили здатність до фотосинтезу і взяли на себе окислення органічних речовин. Завдяки їм еукаріотичні клітини стали аеробними. Симбіози з іншими фотосинтезуючими клітинами стали причиною набуття рослинними клітинами пластид. Можливо, джгутики еукаріотичних клітин виникли в результаті симбіозу клітин‑хазяїв з бактеріями, які були здатні до вигинальних рухів.
Спадковий апарат еукаріотичних клітин був влаштований приблизно так само, як у прокаріот. Однак у зв’язку з необхідністю управління більшою і складнішою клітиною, згодом змінилася організація хромосом, а ДНК виявилася пов’язана з білками‑гістонами. Прокаріотична організація збереглася у геномах внутрішньоклітинних симбіонтів.
У результаті різних актів симбіогенезу виникли різні групи еукаріотичних організмів: еукаріотична клітина + ціанобактерія = червоні водорості; еукаріотична клітина + бактерія‑прохлорофіт = зелені водорості. Навіть хлоропласти золотистих, діатомових, бурих і криптомонадних водоростей виникли в результаті двох послідовних симбіозів, про що свідчить наявність у них 4 мембран.
Поява еукаріот була приурочена до такого періоду історії біосфери, коли умови були особливо нестабільні і непередбачувані, коли адаптивна стратегія прокаріот (швидке мутування, горизонтальний обмін генами і відбір стійких клонів) виявилася надто розтратною і недостатньо ефективною. У такій ситуації велика перевага могла отримати принципово більш універсальна і економічна адаптивна стратегія, заснована на розвитку цілеспрямованої модифікаційної змінливості.
Можливо, становлення еукаріот і розвиток у них статевого процесу зробили структуру змінливості і біорізноманіття більш дискретною і «керованою» — це мало призвести до прискореного зростання біорізноманіття і підвищення еволюційної пластичності та екологічної толерантності видів, спільнот і біоти в цілому.
Появу еукаріот можна справедливо назвати «еталонним» ароматофозом. У цій події надзвичайно яскраво проявилася загальна прогресивна спрямованість біологічної еволюції. Прогрес виразився не лише в ускладненні організації, розширенні сумарної адаптивної зони життя, зростанні біомаси і чисельності, підвищенні автономності організмів, а й у підвищенні стійкості живих систем.
На прикладі еукаріот чітко показано, що появу нових форм життя слід розглядати не як результат еволюції окремих філетичних ліній чи клад, а як закономірний і неминучий ефект розвитку систем вищого порядку — спільнот, біосфери і, можливо, всієї планети як єдиного цілого.
Використані джерела:
A. V. Markov, A. M. Kulykov. Origin of eukaryotes as a result of integrative processes in the microbial community
A. V. Markov. The problem of the origin of eukaryotes
M. V. Laryna. Hypotheses on the origin of eukaryotic cells. Emergence of multicellularity
http://batrachos.com/%D0%AD%D1%82%D0%B0%D0%BF%D1%8B_%D0%B8%D1%81%D1%82%D0%BE%D1%80%D0%B8%D0%B8_%D0%B7%D0%B5%D0%BC%D0%BD%D0%BE%D0%B9_%D0%B6%D0%B8%D0%B7%D0%BD%D0%B8
http://botan0.ru/?cat=2&id=6