
Том 6. Амфібії. 02. Рання еволюція та викопна історія
Grzimek`s Animal Life Encyclopedia. — 2nd ed. — Farminfton Hills, MI: Gale Group, 2003. Volume 6. Amphibians. — 507 p. Early evolution and fossil history. — Р. 7-14.
Рання еволюція та викопна історія
Поява хребетних з кінцівками – стегоцефали
Корені земноводних і амніот (рептилій, птахів і ссавців) слід шукати серед стегоцефалів (тобто чотириногих хребетних з пальцями), які з’явилися близько 370 мільйонів років тому в Девоні. Група кісткових хребетних, від яких виникли чотириногі (група, що включає збережені стегоцефали), неформально відома як кісткові риби, включає дві групи хребетних лучеперих і лопастеперих. Лучепері – група, що містить майже всі кісткові риби, відомі сьогодні. Лопастепері містять три групи: сучасну латимерію, двоякодихих і чотириногих. (Автори енциклопедії дотримуються тут кладистичного підходу, за яким кожна група включає всіх своїх нащадків, і, отже, ссавці та інші чотириногі належать до лопастепери‑риб; це не єдина можлива інтерпретація. – Прим. Д.Ш.) Наші найближчі відомі родичі, які зберегли парні плавці, – це пандерихтиди. Предки пандерихтид і чотириногих не мали спинного та анального плавців, характерних для інших лопастепери‑риб, і, на відміну від них, їх череп не був розділений на передню і задню частини внутричерепним суглобом. Крім того, череп містив нові покривні кістки – лобові.
Пандерихтиди (Panderichthys і Elpistostege) були великими лопастепери‑рибами довжиною близько 39 дюймів (1 м) з пізнього Девону Європи та Північної Америки. На відміну від своїх остеолепіформних родичів (наприклад, Eusthenopteon), пандерихтиди мали масивну, сплющену голову і тіло, а також дуже виступаючий рильце з одним зовнішнім носовим отвором на краю щелепи з кожного боку черепа – цією особливістю вони поділялися з ранніми стегоцефалами. Базальні лучепері (наприклад, многопер і ільна риба), двоякодихі та чотириногі володіли легенями як дихальним органом, тому передбачається, що всі ранні кісткові риби мали легені. У лінії, що веде до сучасних лучепери, легені перетворилися на плавальний міхур (орган плавучості), але у лопастепери‑ріб легені збереглися. Завданням, що стояло перед ранніми водними предками чотириногих, була розробка механізму вентиляції легенів у наземному середовищі.
Найраніші відомі стегоцефали – це тварини помірних розмірів: Ichthyostega (61 дюйм, 1,5 м у довжину) і Acanthostega (19,7 дюйма, 0,5 м у довжину) з Гренландії та Tulerpeton (близько 26,4 дюйма, 0,67 м у довжину) з Росії. На підставі залишків інших тварин, пов’язаних з цими стегоцефалами, і особливостей їх анатомії, цілком ймовірно, що вони вели водний спосіб життя. Acanthostega і Ichthyostega, ймовірно, жили у прісній або солонуватій воді, тоді як Tulerpeton мешкав у морському середовищі. Протягом більшої частини ХХ століття вважалося, що стегоцефали виникли у прісній воді, але все більше ранніх амфібій знаходять у прибережних, ймовірно, солонуватих і морських водах. Деякі класичні пермокарбонові (Гарнетт, Гамільтон і Робінсон, Канзас, США) та девонські (Мігуаша, Квебек, Канада) місця викопних знахідок, які раніше інтерпретувалися як прісноводні, як встановлено зараз, були прибережними, лагунними, дельтовими або устьовими.
Ранні стегоцефали мали багато особливостей, пов’язаних з водним способом життя. Усі вони зберігали систему бокової лінії – серію сенсорних рецепторів у шкірі, що сприймають механічні коливання у воді, типову для риб, а також личинки та водних представників сучасних амфібій. Acanthostega, здається, зберегла функціональні внутрішні жабри на додачу до легенів. І Acanthostega, і Ichthyostega мали хвіст з плавником, схожим на Panderichthys, і, ймовірно, так само використовували його для забезпечення рухової сили під час плавання. Хребетні цих стегоцефалів були погано розвиненими і, ймовірно, нездатними підтримувати вагу тварини на суші. Ichthyostega мала міцну грудну клітку. Перекриваючі ребра забезпечували захист внутрішніх органів, але не були достатньо рухливі, щоб сприяти вентиляції легенів у наземному середовищі. Кінцівки були короткими і товстими, передні кінцівки значно більші за задні. Кінцівки, здається, могли лише до обмежених рухів і, за припущеннями, розташовувалися радше збоку тіла, ніж під ним. І передні, і задні кінцівки були ластоподібними, маючи від шести до восьми пальців у різних видів. Таким чином, ймовірно, що ці девонські хребетні використовували кінцівки для ходіння по зануреному субстрату, можливо, у приливній зоні або в складних середовищах, таких як мангрові болота. Опорно‑руховий апарат цих тварин можна розглядати як підготовлений до наземного способу життя. Наявність кінцівок, пальців, грудного та тазового поясів і грудної клітки, які були корисними у водному середовищі, зумовила морфологічні ознаки, які їх нащадки змогли використати для успішного освоєння наземного середовища.
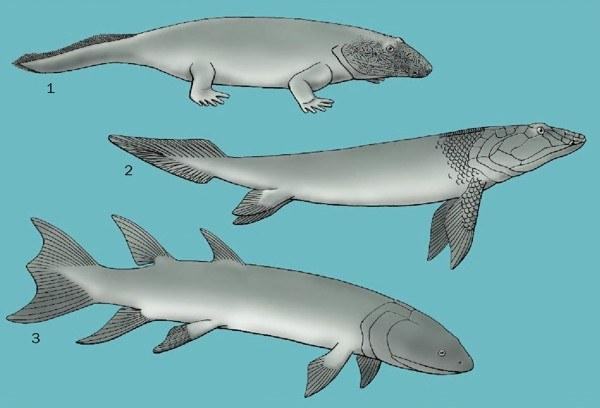
Ранні викопні: 1. Ichthyostega, 2. Panderichthys і 3. Eusthenopteron
Архаїчні амфібії та інші ранні стегоцефали
Є три групи ранніх стегоцефалів: стовбурові тетраподи, амфібії та рептиліоморфи. Стовбурові тетраподи включають усіх стегоцефалів, які з’явилися до дивергенції між амфібіями та рептиліоморфами, дивергенції, яка в кінцевому підсумку привела до появи лиссамфібій і амніот (рептилій, птахів і ссавців) відповідно. Стовбурові тетраподи включають усіх відомих девонських стегоцефалів і, можливо, також багато карбонових і пермських з них. Існує значна невизначеність щодо відносин багатьох з цих таксонів, бо філогенія, яка довгий час приймалася більшістю палеонтологів, була спростована в кінці 1990‑х. Тим не менш, ясно, що основний еволюційний розподіл стегоцефалів відбувся в кінці Девону і на початку Карбону, і що всі основні лінії (включаючи амфібії та рептиліоморфів) існували в нижньому Карбоні. Однак рептиліоморфи не були представлені в палеонтологічному реєстрі до верхнього Карбону.
Різноманіття пост‑девонських стегоцефалів
Бафетиди (раніше відомі як локсоматиди) включають п’ять родів передбачувано водних стегоцефалів з Міссісіпі та Пенсильванії (340‑305 млн років тому) Європи та Північної Америки. Ці викопалини відомі в основному завдяки черепам, які зазвичай широкі і плоскі зі своєрідною формою орбіти, схожою на замкову щілину. Бафетиди можуть бути родичами стовбурових тетрапод.
Три роду крокодилоподібних колостеїдів, які приблизно були сучасниками бафетид, важливі, бо колись вважалися тісно пов’язаними з темноспондилами. Проте черепа цих викопалих в Австралії та Північній Америці позбавлені лускової щілини, особливості, що характеризує темноспондили, і їх відсутність у колостеїдів та наявність органів бокової лінії свідчать про водний спосіб життя.
Темноспондили – це велика група з понад 150 описаних родів, які існували від Міссісіпі до нижнього Мела, протягом приблизно 200 мільйонів років. Більшість ранніх темноспондилів і всі мезозойські представники були водними, але інші були амфібиотичними, а деякі, як вважають, наземними. Серед останніх диссорофід, які заслуговують уваги, бо їх стверджують, що вони тісно пов’язані з сучасними амфібіями. Розміри темноспондилів варіювалися від менш ніж 12 дюймів (30 см) до понад 9,8 фута (3 м) у довжину. Багато, якщо не всі, диссорофіди мали водяні личинки з зовнішніми жабрами. Деякі личинки і/або педоморфні (тобто зберігаючі личинкові або ювенільні ознаки у статевому стані) диссорофіди мали двовершинні (і, можливо, педицеллярні) зуби, як у лиссамфібій, отже, диссорофіди, за думкою деяких вчених, тісно пов’язані з лиссамфібіями. Інші палеонтологи вважають темноспондилів стовбуровими тетраподами. Темноспондили мали велике відкрите небо (інтерптерігоїдні ями), які могли бути залучені до механізму щетинного насоса, схожого на той, що використовують усі лиссамфібії для вентиляції легенів. Стремечко (кістка середнього вуха) більшості темноспондилів тонше і орієнтоване більш латерально, ніж у ранніх стегоцефалів. Через це деякі дослідники вважають, що ці тварини мали тимпанум (барабанну перепонку) у вушній вирізці (або вирізці на лусковій кістці) черепа. Якщо ця гіпотеза вірна, то відсутність тимпануму у хробаків і хвостатих має бути вторинною (тобто наслідком еволюційної втрати). Хребці ранніх темноспондилів зберегли рахітоподібну будову, примітивну для стегоцефалів. Рахітоподібні хребці мали великий, вентральний, серповидний інтерцентр і невеликі парні спинні плевроцентри, що підтримували нервові дуги; такі хребці погано підходять для наземного способу життя. У мезозойських темноспондилів хребет був консолідований за стереоспондільним типом, характеризованим великим інтерцентром і невеликими плевроцентрами, які іноді були хрящовими. Функціональна причина цього зміцнення хребта залишається неясною, бо більшість, якщо не всі, мезозойські темноспондили були виключно водними.
Емболомери мали довжину від 12 дюймів (30 см) до 9,8 фута (3 м) і були в основному водними і земноводними хижаками. Викопалини емболомерів зустрічаються від верхнього Карбону до Тріасу, вони відомі з Європи, Північної Америки та Росії. Їх хребці складаються з циліндричних інтерцентра і плевроцентра. Оскільки вони мають масивне стрижень, вважається, що емболомери були позбавлені барабанної порожнини, але стрижень, можливо, передавав низькочастотні коливання землі та поширювані у воді звуки. Емболомери важливі, бо раніше вважали їх пов’язаними з амніотами. Однак тепер багато вчених розглядають їх як ще одну групу стовбурових чотириногих.
Сеймуріаморфи є ще однією групою стовбурових чотириногих, про яку раніше думали, що вони тісно пов’язані з амніотами. Ця відносно невелика (12 родів), але широко розповсюджена група відома з пермської Північної Америки, Європи та Азії, а також представлена личинками з зовнішніми жабрами та дорослими (близько 3 футів [90 см] у довжину). Відсутність органів бокової лінії та жабр у дорослих свідчить, що вони були наземними. Дорослі сеймуріаморфи мали довгі, тонкі стремена, що дозволяє припустити, що вони, можливо, мали барабанну порожнину, і будову ребер, які передбачають здатність до реберної вентиляції легенів.
Діадектоморфи представляють вісім родів верхнього Карбону і нижньої Пермі. Раніше їх вважали амніотами, тепер їх вважають тісно пов’язаними з ними. Ці тварини досягали довжини 6,5 фута (2 м). Окрім деяких хижих або рибоїдних форм, діадектоморфи включають деякі з найраніших травоїдних стегоцефалів, діадектид.
Сукупність малих амфібій (більшість менше 12 дюймів [30 см] у довжину) включає п’ять груп (аїстоподи, некридиї, «мікрозаври», аделогириніди та лісорофіди) еволюційного рівня, неофіційно відомого як «лепоспонділи». Ці тварини відомі з нижнього Карбону до верхньої Пермі. Деякі, здається, жили виключно у водному середовищі (наприклад, багато некридиї, аделогириніди), тоді як інші (наприклад, «мікрозаври» Pantylus і Tuditanus) очевидно були амфібиотичними або наземними. Більшість лепоспондільних амфібій не мали або мали лише невелику вушну вирізку або западину лускової кістки (наприклад, аделогириніди), таким чином, вони не мали барабанної перепонки. Якщо ці земноводні включають найближчих відомих родичів лиссамфібій, то предок хробаків і саламандр, ймовірно, не мав барабанної перепонки, і тимпанум безхвостих, можливо, з’явився лише у Тріасі. Назва групи походить від структури їх хребців, у яких домінують великі, циліндричні плевроцентри, що зливаються з нервовими дугами на ранніх стадіях розвитку, як це відбувається у лиссамфібій. У деяких лепоспондільних амфібій зберігається невеликий серповидний інтерцентр, але у інших він втрачається, як і у більшості лиссамфібій.
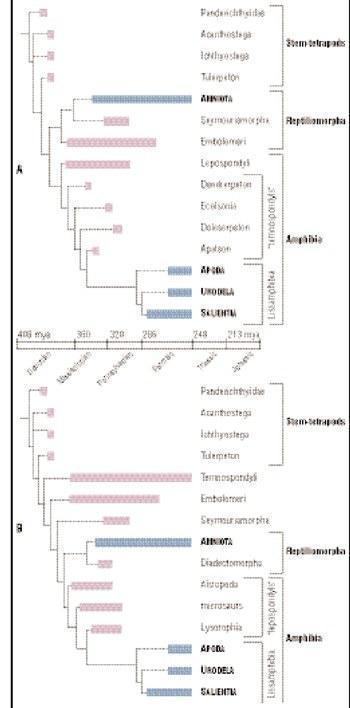
A і B: Можливі філогенетичні дерева амфібій
Лиссамфібії
Найстаріші з відомих лиссамфібій, групи, що включає хробаків, саламандр, безхвостих і їх викопних родичів, датуються Тріасом, близько 250 млн років тому. Викопалини цієї групи надзвичайно скудні. Насправді, у Тріасі, який тривав близько 37 мільйонів років, відомо лише два види лиссамфібій, і обидва вони тісно пов’язані з безхвостими. Не всі палеонтологи погоджуються, що лиссамфібії є природною групою, але більшість неонтологів вважає її монофілетичною. Незалежно від обраних філогенетичних відносин, існує розрив у кілька десятків мільйонів років між сестринською групою (тобто диссорофідами або лісорофідами [? лепоспонділами? – Прим. Д.Ш.]) і першими відомими лиссамфібіями. Починаючи з Юрського періоду (206 млн років тому) викопні лиссамфібії стали більш поширеними, проте найкраще представлені групи, які жили у водному середовищі.
Salientia, стрибуни (безхвості та Triadobatrachus)
Triadobatrachus, найстаріша викопна лиссамфібія, походить із нижнього Тріасу Мадагаскару. Як відомо з одного зразка, практично повного і зчленованого скелета (4,2 дюйма [10,6 см] у довжину), він був тісно пов’язаний з безхвостими амфібіями. Він має багато спільних особливостей черепа з жабами та жабами, але відрізняється від них довшим тулубом, менш спеціалізованим тазовим поясом, короткими кінцівками і коротким хвостом. Інший, трохи молодший викопний, Czatkobatrachus із нижнього Тріасу Польщі, заснований на кількох ізольованих кістках, і, таким чином, про його схожість або філогенетичне положення мало що можна сказати. Починаючи з нижньої Юри, відомі викопні, що належать до кожного з сучасних відділів. Проте знахідки хробаків і загальна якість викопних хвостатих є надзвичайно скудними, на відміну від безхвостих.
Викопний Triadobatrachus і художня реконструкція живої особини
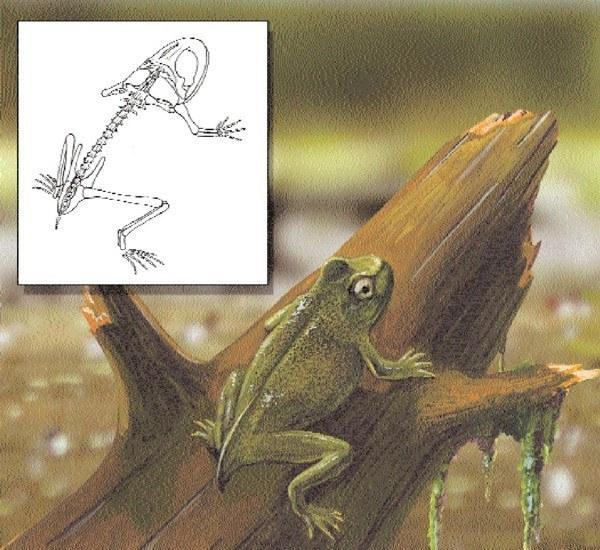
Triadobatrachus вважається близькородичним безхвостим (жабам і жабам). Разом ці тварини утворюють таксономічну групу, відому як Salientia. Найстаріші відомі безхвості, Prosalirus і Vieraella з нижньої Юри Арізони (США) і Аргентини, відповідно, жили приблизно одночасно з Eocaecilia, стовбуровим хробаком. Кожен з цих невеликих за розміром (2 дюйми [50 мм] і 1,25 дюйма [30 мм] у довжину від рила до ануса, відповідно) безхвостих відрізняється від більшого Triadobatrachus наявністю короткого стержня, уростилю, відсутністю хвоста і довгими задніми кінцівками, у яких передплічна кістка змінена і сформувала додаткові відділи кінцівки. На відміну від Triadobatrachus, ці безхвості, звичайно, були здатні до стрибків, типового способу локомоції безхвостих.
Один рід, Eodiscoglossus, представлений середньоюрськими залишками з Великобританії. Це досить велике безхвосте (3,25 дюйма [80 мм] від рила до ануса) чудово нагадує сучасних дискоглоссид, особливо види роду Discoglossus.

Художня реконструкція живого представника Prosalirus bitis
У пізній Юрі викопні залишки безхвостих стають більш різноманітними таксономічно і географічно. Eobatrachus і Comobatrachus відомі з Вайомінгу (США), а Enneabatrachus з Вайомінгу і Юри (США); усі трохи схожі. Notobatrachus, з кількох місць Патагонії в Аргентині, є однією з найважливіших середньоюрських знахідок, бо представлений повними зчленованими скелетами багатьох особин, включаючи молодих і напівдорослих. Це безхвосте було великим, досягало довжини від рила до ануса 5,7 дюйма (14,5 см). Воно мало кілька примітивних особливостей, включаючи вільні ребра у кількох хребцях, слабо розвинений криж, відносно короткий тазовий пояс і товсті, відносно короткі задні кінцівки. Це безхвосте вважається предком усіх сучасних безхвостих. Два інші таксони, Callobatrachus і Mesophryne, були описані з юрських/мелових відкладів Китаю; раніше їх пов’язували з дискоглоссидами, але їх родинні зв’язки невідомі. Найстаріший викопний піповий – це Rhadinosteus з пізньої Юри Юти.
У мелу відбулося поширення безхвостих. Серед найважливіших нижньомелових знахідок дискоглоссида Eodiscoglossus з Іспанії та різних піпових – Thoraciliacus, Cordicephalus і ShomroneIIa (личинки) з Ізраїлю. Середньо/верхньомелові безхвості включають гобіатид з Монголії та Узбекистану, піпових з Аргентини (Saltenia і Avitabatrachus) і Нігеру (Pachybatrachus) і лептодактилід (Baurubatrachus і Estesius) з Бразилії та Болівії відповідно.
Третинні залишки безхвостих надто обширні, щоб описувати їх тут. Достатньо сказати, що більшість родин представлено у викопних відкладеннях Європи, Африки, Азії та Північної Америки.

Викопний скелет жаби в камені. Жаби та жаби вперше з’явилися 190‑160 мільйонів років тому, у ранньому Юрському періоді