
Volume 6. Amphibians. 02. Early evolution and fossil record
Grzimek`s Animal Life Encyclopedia. — 2nd ed. — Farminfton Hills, MI: Gale Group, 2003. Volume 6. Amphibians. — 507 p. Early evolution and fossil history. — pp. 7-14.
Early evolution and fossil history
The emergence of vertebrates with limbs – stegocephalians
The roots of amphibians and amniotes (reptiles, birds and mammals) should be sought among stegocephalians (i.e., tetrapod vertebrates with digits), which appeared about 370 million years ago in the Devonian. The group of bony vertebrates from which the tetrapods (the group that includes the surviving stegocephalians) arose, informally known as bony fishes, comprises two groups of gnathostome vertebrates: ray‑finned and lobe‑finned. Ray‑finned fishes are a group containing almost all bony fishes known today. Lobe‑finned fishes contain three groups: modern coelacanths, lungfishes and tetrapods. (The encyclopedia authors adopt here a cladistic approach in which each group includes all of its descendants, and therefore mammals and other tetrapods are regarded as lobe‑finned fishes; this is not the only possible interpretation. – Note D.Sh.) Our closest known relatives that retained paired fins are the panderichthyids. The ancestors of panderichthyids and tetrapods lacked dorsal and anal fins characteristic of other lobe‑finned fishes, and, unlike them, their skull was not divided into anterior and posterior parts by an intracranial joint. In addition, the skull possessed new covering bones, the frontal bones.
Panderichthyids (Panderichthys and Elpistostege) were large, approximately 39-inch (1 m) long, lobe-finned fishes from the Late Devonian of Europe and North America. Unlike their osteolepiform relatives (e.g., Eusthenopton), panderichthyids had a massive, flattened head and body, and a very protruding snout with a single external nostril at the jaw margin on each side of the skull – a feature they shared with early stegocephalians. Basal ray-finned fishes (e.g., bichirs and bowfin), lungfish, and tetrapods possessed lungs as a respiratory organ, so it is presumed that all early bony fishes had lungs. In the lineage leading to modern ray-finned fishes, the lungs evolved into the swim bladder (an organ of buoyancy), but in lobe-finned fishes, the lungs were retained. The challenge facing the early aquatic ancestors of tetrapods was the development of a mechanism for ventilating the lungs in a terrestrial environment.
The earliest known stegocephals are medium-sized animals: Ichthyostega (61 inches, 1.5 m in length) and Acanthostega (19.7 inches, 0.5 m in length) from Greenland, and Tulerpeton (about 26.4 inches, 0.67 m in length) from Russia. Based on the remains of other animals related to these stegocephals and their anatomical features, it is quite likely that they led an aquatic lifestyle. Acanthostega and Ichthyostega probably lived in fresh or brackish water, while Tulerpeton inhabited a marine environment. For most of the 20th century, it was believed that stegocephals originated in freshwater, but an increasing number of early amphibians are being found in coastal, likely brackish and marine waters. Some classic Permo-Carboniferous (Garnett, Hamilton, and Robinson, Kansas, USA) and Devonian (Miguasha, Quebec, Canada) fossil sites, previously interpreted as freshwater, have now been established as coastal, lagoonal, deltaic, or estuarine.
Early stegocephalians had many features associated with an aquatic lifestyle. All of them retained the lateral line system – a series of sensory receptors in the skin that perceive mechanical vibrations in the water, typical of fish, as well as larvae and aquatic representatives of modern amphibians. Acanthostega appears to have retained functional internal gills in addition to lungs. Both Acanthostega and Ichthyostega had a tail with a fin, similar to Panderichthys, and likely used it similarly for propulsive force during swimming. The vertebrae of these stegocephalians were poorly developed and likely incapable of supporting the animal's weight on land. Ichthyostega had a robust rib cage. The overlapping ribs provided protection for internal organs but were not sufficiently mobile to aid in lung ventilation in a terrestrial environment. The limbs were short and stout, with the forelimbs significantly larger than the hindlimbs. The limbs appear to have allowed only for limited movement and were presumably positioned more to the sides of the body than beneath it. Both forelimbs and hindlimbs were paddle-like, having six to eight digits in different species. Thus, it is likely that these Devonian vertebrates used their limbs for walking on submerged substrates, possibly in the intertidal zone or in complex environments such as mangrove swamps. The musculoskeletal system of these animals can be considered prepared for a terrestrial lifestyle. The presence of limbs, digits, pectoral and pelvic girdles, and a rib cage, which were useful in an aquatic environment, led to morphological features that their descendants could use to successfully colonize terrestrial environments.
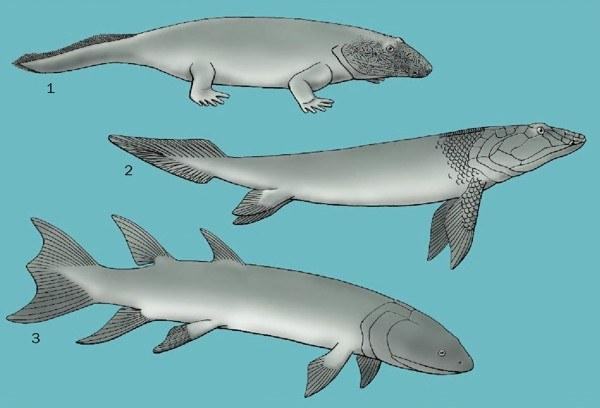
Early fossils: 1. Ichthyostega, 2. Panderichthys, and 3. Eusthenopteron
Archaic amphibians and other early stegocephalians
There are three groups of early stegocephalians: stem tetrapods, amphibians, and reptiliomorphs. Stem tetrapods include all stegocephalians that appeared before the divergence between amphibians and reptiliomorphs, a divergence that eventually led to the appearance of Lissamphibia and Amniota (reptiles, birds, and mammals), respectively. Stem tetrapods include all known Devonian stegocephalians and possibly many Carboniferous and Permian ones as well. There is considerable uncertainty regarding the relationships of many of these taxa, because the phylogeny that was long accepted by most paleontologists was overturned in the late 1990s. Nevertheless, it is clear that the major evolutionary split of stegocephalians occurred in the Late Devonian and Early Carboniferous, and that all major lineages (including amphibians and reptiliomorphs) existed in the Early Carboniferous. However, reptiliomorphs were not represented in the fossil record until the Late Carboniferous.
Diversity of post-Devonian stegocephalians
Baphetiids (formerly known as loxomatids) include five genera of presumably aquatic stegocephalians from the Mississippian and Pennsylvanian (340–305 Ma) of Europe and North America. These fossils are known mainly from skulls, which are usually broad and flat with a strange orbit shape reminiscent of a keyhole. Baphetiids may be relatives of basal tetrapods.
Three genera of crocodilomorphs, the collosuchids, which were roughly contemporaneous with the bafetids, are important because they were once thought to be closely related to the temnospondyls. However, the skulls of these fossils from Australia and North America lack the scaly fissure, a feature that characterizes the temnospondyls, and their absence in the collosuchids and the presence of lateral line organs suggest an aquatic lifestyle.
Temnospondyls are a large group comprising more than 150 described genera, persisting from the Mississippian to the Lower Cretaceous, roughly 200 million years. Most early temnospondyls and all Mesozoic representatives were aquatic, but others were amphibious, and some are thought to have been terrestrial. Among the later dissorophids, which attract attention because they are claimed to be closely related to modern amphibians, size ranged from less than 12 in (30 cm) to over 9.8 ft (3 m) in length. Many, if not all, dissorophids had aquatic larvae with external gills. Some larval and/or pedomorphic (i.e., retaining larval or juvenile traits in the adult) dissorophids possessed bicuspid (and possibly pedicellate) teeth like those of lissamphibians, suggesting that dissorophids, as some researchers propose, are closely linked to lissamphibians. Other paleontologists consider temnospondyls to be basal tetrapods. Temnospondyls had a large open palate (interpterygoid vacuities) that may have been involved in a buccal pumping mechanism similar to that used by all lissamphibians for lung ventilation. The stapes of most temnospondyls is thinner and oriented more laterally than in early stegocephalians. Because of this, some researchers argue that these animals possessed a tympanum (eardrum) in the otic notch (or in a groove of the squamosal) of the skull. If this hypothesis is correct, the absence of a tympanum in caecilians and caudates would be secondary (i.e., a loss). The vertebrae of early temnospondyls retained a rhachitomous structure, primitive for stegocephalians. Rhachitomous vertebrae have a large ventral, sickle‑shaped intercentrum and small paired neural pleurocentra supporting the neural arches; such vertebrae are poorly suited for a terrestrial lifestyle. In Mesozoic temnospondyls the vertebral column was consolidated into a stereospondyl type, characterised by a large intercentrum and small pleurocentra, sometimes cartilaginous. The functional reason for this vertebral strengthening remains unclear, because most, if not all, Mesozoic temnospondyls were exclusively aquatic.
Embolomers were 12 inches (30 cm) to 9.8 feet (3 m) long and were primarily aquatic and semi-aquatic predators. Embolomer fossils are found from the Upper Carboniferous to the Triassic, and they are known from Europe, North America, and Russia. Their vertebrae consist of cylindrical intercentra and pleurocentra. Because they had a massive notochord, it is thought that embolomers lacked a tympanic cavity, but the notochord may have transmitted low-frequency ground vibrations and sounds propagating in water. Embolomers are important because they were previously thought to be related to amniotes. However, many scientists now consider them to be another group of stem tetrapods.
Seymouriamorphs are another group of stem tetrapods that were previously thought to be closely related to amniotes. This relatively small (12 genera) but widespread group is known from the Permian of North America, Europe, and Asia, and is represented by larvae with external gills and adults (about 3 feet [90 cm] in length). The absence of lateral line organs and gills in adults suggests they were terrestrial. Adult seymouriamorphs had long, slender stapes, suggesting they may have possessed a tympanic cavity, and a rib structure that implies the ability for costal ventilation of the lungs.
Diadectomorphs represent eight genera from the Upper Carboniferous and Lower Permian. They were formerly considered to be amniotes, but are now considered to be closely related to them. These animals reached lengths of 6.5 feet (2 m). In addition to some predatory or piscivorous forms, diadectomorphs include some of the earliest herbivorous stegocephalians, the diadectids.
The assemblage of small amphibians (most less than 12 inches [30 cm] long) includes five groups (Aistopoda, Nectridea, 'microsaurs', Adelospondyli, and Lysorophia) of evolutionary grade, informally known as 'lepospondyls.' These animals are known from the Early Carboniferous to the Late Permian. Some appear to have lived exclusively in aquatic environments (e.g., many Nectridea, Adelospondyli), whereas others (e.g., the 'microsaurs' Pantylus and Tuditanus) were apparently amphibian or terrestrial. Most lepospondyl amphibians lacked or had only a small otic notch or squamosal embayment (e.g., Adelospondyli), thus they lacked a tympanic membrane. If these amphibians include the closest known relatives of lissamphibians, then the ancestor of frogs and salamanders probably lacked a tympanic membrane, and the tympanum of anurans may have appeared only in the Triassic. The group's name derives from the structure of their vertebrae, which are dominated by large, cylindrical pleurocentra that fuse with the neural arches at an early stage of development, as occurs in lissamphibians. Some lepospondyl amphibians retain a small, crescent-shaped intercentrum, but others lose it, as do most lissamphibians.
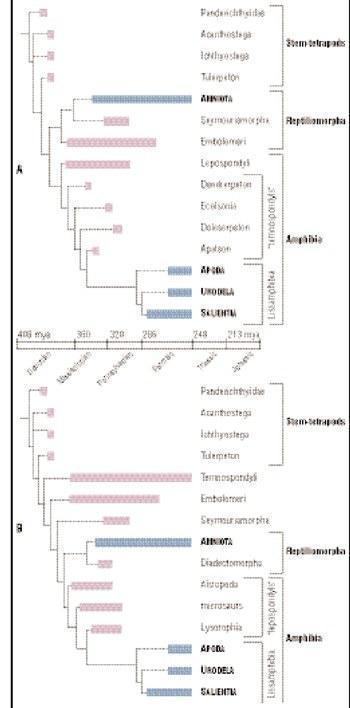
A and B: Possible phylogenetic trees of amphibians
Lissamphibia
The oldest known lissamphibians, a group that includes caecilians, salamanders, caudates and their fossil relatives, date to the Triassic, about 250 Ma. Fossils of this group are extremely scarce. In fact, during the Triassic, which lasted about 37 million years, only two lissamphibian species are known, both closely related to caudates. Not all paleontologists agree that lissamphibians constitute a natural group, but most consider it monophyletic. Regardless of the chosen phylogenetic relationships, there is a gap of several tens of millions of years between the sister group (i.e., dissorophids or lysorophids [? lepospondyls? – Note D.Sh.]) and the earliest known lissamphibians. Starting in the Jurassic (206 Ma) fossil lissamphibians become more common, but the best‑represented groups are those that lived in aquatic environments.
Salientia, jumpers (anurans and Triadobatrachus)
Triadobatrachus, the oldest fossil lissamphibian, is from the Early Triassic of Madagascar. Known from a single specimen, a virtually complete and articulated skeleton (4.2 inches [10.6 cm] long), it was closely related to the anurans. It shares many cranial features with frogs and toads but differs from them in its longer trunk, less specialized pelvic girdle, short limbs, and short tail. Another, slightly younger fossil, Czatkobatrachus from the Early Triassic of Poland, is known from a few isolated bones, and thus little can be said about its similarity or phylogenetic position. Beginning in the Early Jurassic, fossils are known belonging to each of the modern orders. However, the finds of salamanders and the general quality of fossil caudates are extremely sparse, in contrast to the anurans.
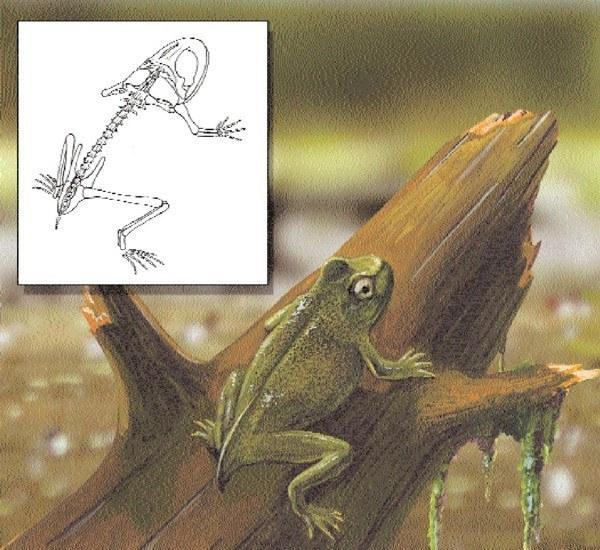
Fossil Triadobatrachus and artistic reconstruction of a living individual
Triadobatrachus is considered a close relative of the anurans (frogs and toads). Together, these animals form the taxonomic group known as Salientia. The oldest known anurans, Prosalirus and Vieraella from the Lower Jurassic of Arizona (USA) and Argentina, respectively, lived at approximately the same time as Eocaecilia, a stem caecilian. Each of these small-sized (2 inches [50 mm] and 1.25 inches [30 mm] snout-to-vent length, respectively) anurans differs from the larger Triadobatrachus by the presence of a short rod, the urostyle, absence of a tail, and long hind limbs in which the forearm bone is modified and has formed additional limb segments. Unlike Triadobatrachus, these anurans were, of course, capable of jumping, the typical mode of locomotion of anurans.
One genus, Eodiscoglossus, is represented by Middle Jurassic remains from Great Britain. This rather large anuran (3.25 inches [80 mm] snout-to-vent length) closely resembles modern discoglossoids, especially species of the genus Discoglossus.

Artistic reconstruction of a living representative of Prosalirus bitis
In the Late Jurassic, fossil remains of anurans become more diverse taxonomically and geographically. Eobatrachus and Comobatrachus are known from Wyoming (USA), and Enneabatrachus from Wyoming and Utah (USA); all are somewhat similar. Notobatrachus, from several localities in Patagonia, Argentina, is one of the most important Middle Jurassic finds because it is represented by complete articulated skeletons of many individuals, including juveniles and subadults. This anuran was large, reaching a snout-to-vent length of 5.7 inches (14.5 cm). It possessed several primitive features, including free ribs in several vertebrae, a poorly developed sacrum, a relatively short pelvic girdle, and stout, relatively short hind limbs. This anuran is considered the ancestor of all modern anurans. Two other taxa, Callobatrachus and Mesophryne, have been described from Jurassic/Cretaceous deposits of China; they were formerly allied with the discoglossoids, but their familial relationships are unknown. The oldest fossil pipid is Rhadinosteus from the Late Jurassic of Utah.
In the Cretaceous, anurans became widespread. Among the most important Early Cretaceous finds are the discoglossoid Eodiscoglossus from Spain and various pipids—Thoraciliacus, Cordicephalus, and ShomroneIIa (larvae)—from Israel. Middle/Late Cretaceous anurans include gobibatids from Mongolia and Uzbekistan, pipids from Argentina (Saltenia and Avitabatrachus) and Niger (Pachybatrachus), and leptodactylids (Baurubatrachus and Estesius) from Brazil and Bolivia, respectively.
Tertiary anuran remains are too extensive to describe here. Suffice it to say that most families are represented in the fossil deposits of Europe, Africa, Asia, and North America.

Fossil frog skeleton in stone. Frogs and toads first appeared 190–160 million years ago, in the Early Jurassic.{ "translated_text": "Gymnophiona (caecilians and Eocaecilia)\n\n\tThe limbless or caecilian forms that have reached us constitute a distinctive group of limbless, snake‑like amphibians, which are terrestrial or aquatic and specialized for burrowing; they possess a tentacle on each side of the head beneath reduced eyes. Most scientists consider caecilians to be the closest to the base of the Lissamphibia (the latest finds prompt a revision of this view, assigning caecilians to the lepospondyl amphibians – note D.Sh.). The Lower Jurassic (about 204 million years ago) fossil Eocaecilia from Arizona is regarded as a stem caecilian. It differs from modern members of this group by having small, well‑ossified limbs and limb girdles, which are absent in extant caecilians. Although the eyes were larger and the skull contained more bones than in living species, the lower jaw and the jaw‑closing mechanism appear to resemble those of modern representatives. All other fossil remains associated with gymnophionans are vertebrae from the Upper Cretaceous of Sudan, the early Paleocene of Bolivia and the late Paleocene of Brazil. These fossils provide little useful information beyond the fact that modern caecilian families began to differentiate in the Late Mesozoic.\n\n[IMG_6]\n\nFossil Eocaecilia and artistic reconstruction of a living specimen\n\nTail‑bearing amphibians (salamanders and similar fossils)\n\n\tFossil tail‑bearers are known from the Middle Jurassic – Lower Cretaceous (about 180–127 million years ago) in lacustrine and lagoonal deposits of Europe, Upper Cretaceous – Eocene (about 90–35 million years ago) fluvial deposits of North America and Eocene‑Miocene (about 35–5 million years ago) brown coal of Europe. Additionally, there are remains from the Cretaceous of Bolivia, Sudan, Niger and Israel. Laccotriton, Sinerpeton, Jehohtriton and Liaoxitriton are tail‑bearers from the Upper Jurassic/Lower Cretaceous of China. With three exceptions listed below, these remains are clearly tail‑bearers, i.e., belonging to Caudata. The phylogenetic relationships of Albanerpetontidae from Middle Jurassic – Miocene deposits of the northern continents are ambiguous. This group may represent a distinct, earlier branch of the tail‑bearer line or a separate clade of Lissamphibia whose origin predates the phylogenetic divergence between tailed and tailless forms. The phylogenetic status of Ramonellus from the Lower Cretaceous of Israel is unknown. The third taxon, Triassurus, was described from the Triassic of Uzbekistan. In the latter case, if the poorly preserved remains turn out to be tail‑bearers, this group originated in the Triassic or earlier.\n\n\tThe earliest remains that can be unequivocally identified as tail‑bearers are Jurassic finds from China, the karauroid tail‑bearers Karaurus and Kokartus from Kazakhstan and Kyrgyzstan, and Marmorerpeton from England. Existing salamander groups exhibit an astonishing diversity of fossils. (1) Sirenidae: Upper Cretaceous and Paleocene of North America; Cretaceous remains of several taxa from southern continents may also belong to Sirenidae; (2) Hynobiidae: Upper Pliocene of Kazakhstan; (3) Cryptobranchidae: Paleocene of Mongolia and Russia, Upper Oligocene‑Pliocene of Europe, Upper Paleocene‑Upper Miocene of North America with Pliocene‑Pleistocene cryptobranchid sites in North America; (5) salamandroids (Ambystomatidae, Amphiumidae, Dicamptodontidae, Plethodontidae, Proteidae, Rhyacontritonidae and Salamandridae): stem salamandroids from the Lower Cretaceous of Europe; amphiumids from the Pliocene‑Pleistocene of North America; dicamptodontids from the Pliocene of Canada; salamandrids from the Upper Paleocene onward in Europe and the Upper Miocene of East Asia and the Neogene of North America. In addition, three groups of fossil tail‑bearers with uncertain affinities are known: batrachosaurids from the Middle Cretaceous – Lower Pliocene of North America and Europe; scaferpetonids from the Upper Cretaceous, Paleocene and Eocene of North America and possibly the Cretaceous of Asia; and dicamptodontid‑like tail‑bearers from the Pliocene‑Miocene of Europe." }