
Касум Ахмедли: модель перехода от гермафродитизма к раздельнополости
После публикации колонки о переходе от гермафродитизма к раздельнополости я (D.Sh.) получил письмо от Касума Ахмедли, ведущего научного сотрудника Национальной академии наук Азербайджана. Он прислал свою модель такого перехода и ее детальное описание. Я выкладываю здесь это письмо (с согласи...
Після публікації колонки про перехід від гермафродитизму до роздільностатевості я (D.Sh.) отримав листа від Касума Ахмедлі, провідного наукового співробітника Національної академії наук Азербайджану. Він надіслав свою модель такого переходу та її детальний опис. Я викладаю тут цей лист (за згодою його автора), сподіваючись, що це сприятиме кращому розумінню обговорюваного еволюційного переходу.
Шановний Дмитре!
Я вирішив промоделювати еволюцію статевого розмноження, як Ви описали в колонці "Перемога стійкості над оптимальністю, або Чому гермафродити програють самцям та самкам" від 24/01/14, засобами хімічної кінетики і зараз хочу поділитися з Вами результатами. Хоча в моїх моделях процеси протікають під керуванням закону дії мас, та й розмножувані форми вибрані гранично прості, тим не менш, мені здається, вони дозволяють у грубому наближенні судити про ефекти переходу від нестатевого розмноження гермафродитів до статевого. Не сумніваюся, у Вашому розпорядженні є інструменти точнішого математичного моделювання еволюційних механізмів, тож не судіть суворо, що скористався тим, що мав під рукою - програмою обрахунку кінетики хімічних реакцій Gepassi. Концепція егоїстичного гена в цьому підході полягає в тому, що в умовах обмеженого ресурсу харчового субстрату (SS) між колоніями, що споживають цей ресурс, виникає конкуренція, в якій виживає ген, що відтворюється швидше.
Спочатку опишу модель, а потім уже перейду до результатів.
Позначення
• SS і S - димерний субстрат (і його легко димеризований мономер), на якому реплікують та еволюціонують гени популяції. Кількість субстрату обмежена (інакше не може бути добору).
• HHxx - геном диплоїдного монозиготного гермафродита вихідної популяції.
• HMxx, HMxy, HFxx, MMxx, MMxy, FMxx, FMxy, FFxx - інші диплоїдні геноми (зиготи, реплікатори), що утворилися в результаті мутацій вихідного гермафродита та рекомбінацій статевого процесу.
• H, M, F - гени, що визначають статеву поведінку гермафродита, самця або самки (H підтримує як оогенез, так і сперматогенез, M - підтримує тільки сперматогенез, F - підтримує тільки оогенез). Ген H домінантний по відношенню до своїх мутантів M і F. Тому комбінації HMxx, HMxy, HFxx визначають гермафродитів, комбінації MMxy і FMxy - самців, а FMxx і FFxx - самок. Комбінація MMxx, незважаючи на відсутність y-хромосоми, також віднесена до самців, оскільки такі зразки в мейозі, за відсутності генів H або F, виробляють тільки сперматозоїди.
• Hxe, Mxe, і Fxe - гаплоїдні яйцеклітини (e - позначає egg).
• Hx, Mx, My і Fx - гаплоїдні сперматозоїди.
Одиниці вимірювання
• За одиницю кількості будь-якого зразка прийнято вихідну початкову кількість реплікаторів з геномом HHxx.
• За одиницю часу - середня тривалість життя вихідного реплікатора (особини) HHxx, яка зберігається такою ж і для мутантів.
Опис моделі
У табл.1 наведена схема, що описує еволюцію статевого розмноження у гермафродитів.
Таблиця 1. Схема процесів розмноження та еволюції
|
N |
Процесс |
k→ |
Примечания |
|---|---|---|---|
|
Мутации |
|||
|
1 |
HHxx → HMxx |
Kmu |
Самцовая мутация |
|
2 |
HHxx → HFxx |
Kmu |
Самочная мутация |
|
3 |
HMxx → MMxy |
Kmu |
Возникновение самца |
|
4 |
HMxx → FMxx |
Kmu |
Возникновение самки |
|
Митоз |
|||
|
5 |
SS + HHxx → 2*HHxx |
Kmi |
|
|
6 |
SS + HMxx → 2*HMxx |
Kmi |
|
|
7 |
SS + HMxy → 2*HMxy |
Kmi |
|
|
8 |
SS + HFxx → 2*HFxx |
Kmi |
|
|
9 |
SS + MMxx → 2*MMxx |
Kmi |
|
|
10 |
SS + MMxy → 2*MMxy |
Kmi |
|
|
11 |
SS + FMxx → 2*FMxx |
Kmi |
|
|
12 |
SS + FMxy → 2*FMxy |
Kmi |
|
|
13 |
SS + FFxx → 2*FFxx |
Kmi |
|
|
Мейоз |
|||
|
14 |
HHxx → 2*Hxe |
Kh |
Оогенез |
|
15 |
HHxx → 2*Hx |
Kh |
Сперматогенез |
|
16 |
HMxx → Hxe+ Mxe |
Kh |
О |
|
17 |
HMxx → Hx+ Mx |
Kh |
С |
|
18 |
HMxy → Hx + My |
Kh |
С |
|
19 |
HFxx → Hxe + Fxe |
Kh |
О |
|
20 |
HFxx → Hx + Fx |
Kh |
С |
|
21 |
MMxx → 2*Mx |
Kh |
С |
|
22 |
MMxy → Mx + My |
Ks*Kh |
С |
|
23 |
FMxx → Mxe + Fxe |
Ke*Kh |
О |
|
24 |
FMxy → Fx + My |
Ks*Kh |
С |
|
25 |
FFxx → 2*Fxe |
Ke*Kh |
О |
|
Оплодотворение |
|||
|
26 |
Hxe + Hx → HHxx |
Kf |
|
|
27 |
Hxe + Mx → HMxx |
Kf |
|
|
28 |
Mxe + Hx → HMxx |
Kf |
|
|
29 |
Hxe + My → HMxy |
Kf |
|
|
30 |
Hxe + Fx → HFxx |
Kf |
|
|
31 |
Fxe + Hx → HFxx |
Kf |
|
|
32 |
Mxe + Mx → MMxx |
Kf |
|
|
33 |
Mxe + My → MMxy |
Kf |
|
|
34 |
Mxe + Fx → FMxx |
Kf |
|
|
35 |
Fxe + Mx → FMxx |
Kf |
|
|
36 |
Fxe + My → FMxy |
Kf |
|
|
37 |
Fxe + Fx → FFxx |
Kf |
|
|
Смерть |
|||
|
38 |
HHxx → SS |
Kd |
Разрушение зигот |
|
39 |
HMxx → SS |
Kd |
|
|
40 |
HMxy → SS |
Kd |
|
|
41 |
HFxx → SS |
Kd |
|
|
42 |
MMxx → SS |
Kd |
|
|
43 |
MMxy → SS |
Kd |
|
|
44 |
FMxx → SS |
Kd |
|
|
45 |
FMxy → SS |
Kd |
|
|
46 |
FFxx → SS |
Kd |
|
|
47 |
Hxe → S |
Kde |
Разрушение ооцитов |
|
48 |
Hx → S |
Kds |
Разрушение сперматозоидов |
|
49 |
Mxe → S |
Kde |
|
|
50 |
Mx → S |
Kds |
|
|
51 |
My → S |
Kds |
|
|
52 |
Fxe → S |
Kde |
|
|
53 |
Fx → S |
Kds |
|
|
54 |
2*S → SS |
Kss |
Регенерация субстрата |
Стовпець k→ - це константи швидкостей процесів. Значення констант швидкостей наведено в табл.2, а початкові умови – в табл. 3.
Таблиця 2. Константи швидкостей
|
Константи швидкості |
Примечания |
|
|
Kmu |
1.00E-04 |
К.С. мутации |
|
Kmi |
0.1 |
К.С. митоза |
|
Kh |
1 |
К.С. мейоза у гермафродитов |
|
Ks |
1 |
Ускорение сперматогенеза M |
|
Ke |
1 |
Ускорение оогенеза F |
|
Kf |
1.00E+04 |
К.С. оплодотворения |
|
Kd |
1 |
К.С. разрушения репликаторов |
|
Kde |
0.05 |
К.С. разрушения ооцитов |
|
Kds |
0.05 |
К.С. разрушения сперматозоидов |
|
Kss |
100 |
К.С. регенерации субстрата |
Таблиця 3. Початкові умови
|
Початкові умови |
|
|
|
10 |
|
|
1 |
|
Усі інші |
0 |
 0
0Той факт, що одиницею часу обрано середню тривалість життя реплікаторів, відображається в тому, що в процесах 36-43 константа швидкості руйнування для них усіх встановлена Kd=1. Початкове значення чисельності гермафродитів
При заплідненні в кожному акті беруть участь 1 яйцеклітина та 1 сперматозоїд.
Поки геном містить домінантний ген H, реплікатор (HHxx, HMxx, HFxx) у мейозі здатний як до оогенезу, так і до сперматогенезу і є гермафродитом. Мутація (1) призводить до виникнення гена M, під керуванням якого (а значить за відсутності в геномі генів H або F) втрачається здатність до оогенезу. Аналогічно, мутація (2) призводить до виникнення гена F, під керуванням якого (за відсутності в геномі генів H або M) втрачається здатність до сперматогенезу.
Обговорення результатів моделювання
Модель 1
Як 1-у модель розглянуто найпростіший випадок, коли жодних мутацій немає (Kmu=0), і єдиним реплікатором у системі є вихідний гермафродит HHxx. На рис.1 наведені графіки розвитку рівноваги в цьому випадку.
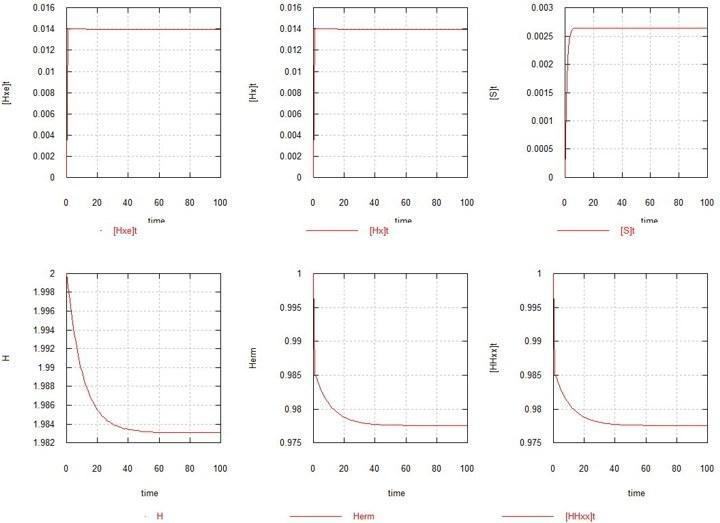
Рис. 1. Розвиток стаціонарного стану в популяції гомозиготного гермафродита.
H – сумарний вміст гена H у системі. Herm – кількість зигот HHxx.
Як видно і, як і слід було очікувати, рівновага (точніше, стаціонарний стан) досягається за кілька поколінь. Кількість зигот знижується на пару %, унаслідок досягнення гаметами стаціонарної кількості по 1.4% відносно зигот кожна. Кількість субстрату SS (не показано) практично не змінюється (близько 10).
Модель 2
Розглянемо тепер що відбувається, коли обидві мутації (M і F) вмикаються. Коефіцієнти Ks та Ke приймемо рівними 1, тобто сперматогенез у М-самців та оогенез у F-самок ідуть з тими ж швидкостями, що і у вихідного гермафродита. Ми повинні очікувати, що оскільки в цій ситуації в самок відбувається втрата 50% фертильності, гермафродити повинні витіснити мутантів, що з'являються. Однак, проти цих очікувань, відбувається те, що зображено на рис. 2.
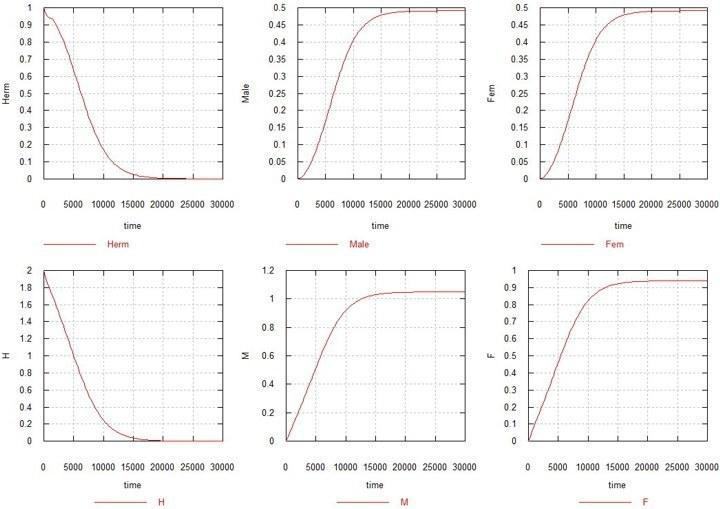
Рис. 2. Еволюція статі при «самцевій» та «самочній» мутаціях у популяції гермафродитів.
H, M і F – сумарні кількості відповідних генів у системі (з урахуванням їх наявності в усіх видах зигот та гамет). Herm – сумарна кількість гермафродитів: HHxx, HMxx і HFxx. Male – сумарна кількість самців: HMxy, MMxx, MMxy і FMxy. Fem – сумарна кількість самок: FMxx і FFxx.
Як видно з рисунка, через кілька десятків тисяч поколінь кількість гермафродитів, разом із загальним вмістом вихідного гена H, падає до нуля, і утворюється популяція, що складається виключно із самців та самок, з половинною чисельністю кожна, порівняно з вихідною чисельністю гермафродитів.
Додам (не нагромаджуючи рисунків), що варіації Ks у той чи інший бік між значеннями 0.5 і 1.5 не призводить до якісної зміни картини, дещо змінюючи лише співвідношення статей.
Модель 3
Розглянемо тепер, що відбувається, якщо відбувається тільки самцева M-мутація або тільки самоча F-мутація. Спочатку залишимо самцеву мутацію та відключимо самочу (виключимо процеси 2 і 4). Ks так само =1.
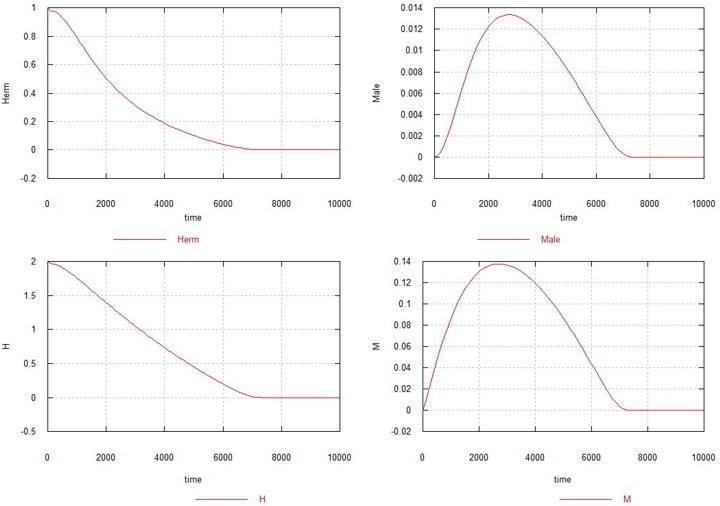
Рис. 3. Динаміка виникнення самцевої мутації в популяції гомозиготних гермафродитів. Позначення – як раніше.
Ось тут з'ясовується, що якщо виникає тільки самцева мутація, і за нею не йде самоча мутація, то мутація поширюється, як хвороба, і через кілька тисяч поколінь уся популяція вимирає. Збільшення Ks також не допомагає стабілізувати популяцію.
Модель 4
Схожа ситуація виникає і в тому випадку, коли має місце тільки «самоча» мутація. Повернемо на місце процес 2 та виключимо процеси 1 і 3.
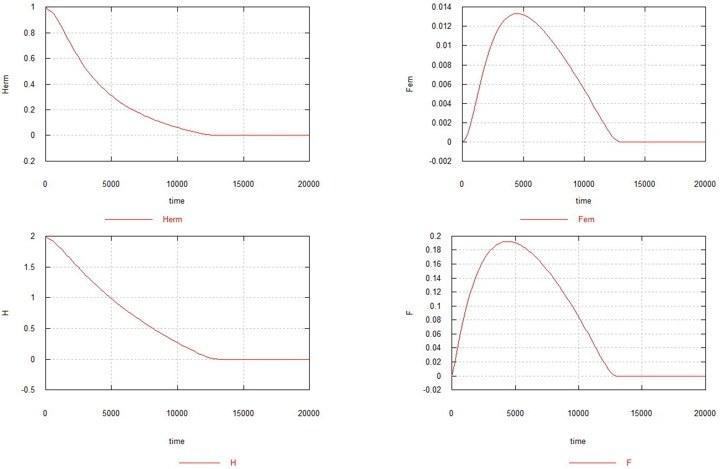
Рис. 4. Динаміка виникнення самочої мутації в популяції гомозиготних гермафродитів. Позначення – як раніше.
Як видно з рисунка, самоча мутація за відсутності самцевої також призводить до повного вимирання популяції, хоча й дещо повільніше. Збільшення Ke до 1.5 у цьому випадку (див. рис. 5) призводить до того, що популяція гермафродитів не вимирає, але різко скорочується (до 0.7) і залишається дуже невелика стаціонарна кількість самок - 0.005. Вміст генів H і F стабілізується на рівні 1.6 (вихідний вміст становив 2.0) і 0.14, відповідно.
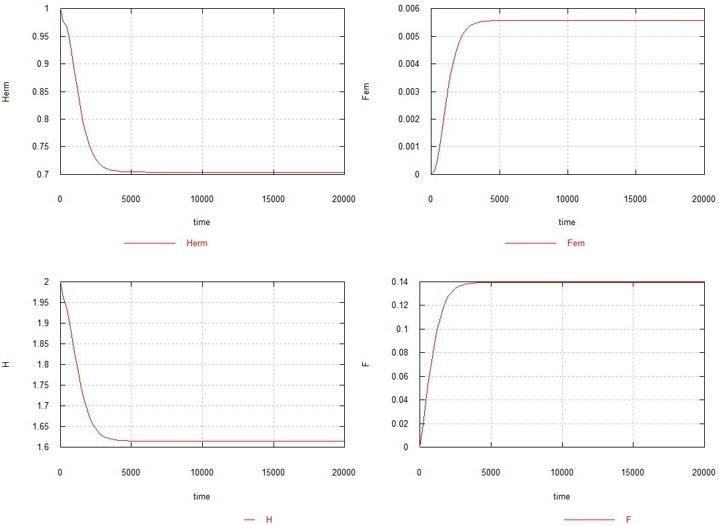
Рис. 5. Умови ті самі, що на рис. 4, з тією відмінністю, що Ke=1.5.
Висновки
Проти інтуїтивного очікування, якщо в популяції гермафродитів відбуваються самцева та самоча мутації синхронно або з не дуже великим запізненням будь-якої з них після іншої, то, незважаючи на втрату 50% плодючості в самок та самців, популяція гермафродитів з часом перетворюється на роздільностатеву популяцію. Однак, якщо відбувається тільки одна з цих мутацій, а інша так і не настає, це справляє згубний вплив на всю популяцію та знижує її чисельність (не будемо говорити, що вона гине, оскільки модель груба, а життя – чіпка штука).