
Kasum Akhmedli: A Model of the Transition from Hermaphroditism to Gonochorism
After publishing the column on the transition from hermaphroditism to gonochorism, I (D.Sh.) received a letter from Kasum Akhmedli, a leading researcher at the National Academy of Sciences of Azerbaijan. He sent his model of such a transition and its detailed description. I post this letter here (with permis...
After publishing the column about the transition from hermaphroditism to separate sexes I (D.Sh.) received a letter from Kasum Akhmedli, a leading researcher at the National Academy of Sciences of Azerbaijan. He sent me his model of this transition and a detailed description of it. I am publishing this letter here (with the author's consent), hoping it will contribute to a better understanding of the evolutionary transition under discussion.
Dear Dmytro!
I decided to model the evolution of sexual reproduction, as you described in the column "The Victory of Stability over Optimality, or Why Hermaphrodites Lose to Males and Females" from 24/01/14, by means of chemical kinetics, and now I want to share the results with you. Although in my models the processes proceed under the law of mass action, and the reproducing forms chosen are extremely simple, nevertheless it seems to me they allow, in a rough approximation, judging the effects of the transition from asexual reproduction of hermaphrodites to sexual reproduction. I have no doubt that you have at your disposal tools for more precise mathematical modeling of evolutionary mechanisms, so do not judge me too harshly for using what I had at hand — the Gepasi program for calculating the kinetics of chemical reactions. The concept of the selfish gene in this approach is that, under conditions of a limited resource of the food substrate (SS), competition arises between colonies consuming this resource, in which the gene that replicates faster survives.
First I will describe the model, and then move on to the results.
Notation
• SS and S - a dimeric substrate (and its easily dimerized monomer) on which the genes of the population replicate and evolve. The amount of substrate is limited (otherwise selection cannot occur).
• HHxx - the genome of a diploid monozygous hermaphrodite of the original population.
• HMxx, HMxy, HFxx, MMxx, MMxy, FMxx, FMxy, FFxx - other diploid genomes (zygotes, replicators) that arose as a result of mutations of the original hermaphrodite and recombinations of the sexual process.
• H, M, F - genes that determine the sexual behavior of a hermaphrodite, male, or female (H supports both oogenesis and spermatogenesis, M supports only spermatogenesis, F supports only oogenesis). Gene H is dominant relative to its mutants M and F. Therefore the combinations HMxx, HMxy, HFxx define hermaphrodites, the combinations MMxy and FMxy define males, and FMxx and FFxx define females. The combination MMxx, despite the absence of a y-chromosome, is also classed as male, since such specimens, at meiosis, in the absence of genes H or F, produce only spermatozoa.
• Hxe, Mxe, and Fxe - haploid egg cells (e - denotes egg).
• Hx, Mx, My and Fx - haploid spermatozoa.
Units of measurement
• The unit of quantity for any sample is taken to be the original initial quantity of replicators with the genome HHxx.
• The unit of time is the average lifespan of the original replicator (individual) HHxx, which remains the same for mutants as well.
Model description
Table 1 gives the scheme describing the evolution of sexual reproduction in hermaphrodites.
Table 1. Scheme of reproduction and evolution processes
|
N |
Process |
k→ |
Notes |
|---|---|---|---|
|
Mutations |
|||
|
1 |
HHxx → HMxx |
Kmu |
Male mutation |
|
2 |
HHxx → HFxx |
Kmu |
Female mutation |
|
3 |
HMxx → MMxy |
Kmu |
Emergence of a male |
|
4 |
HMxx → FMxx |
Kmu |
Emergence of a female |
|
Mitosis |
|||
|
5 |
SS + HHxx → 2*HHxx |
Kmi |
|
|
6 |
SS + HMxx → 2*HMxx |
Kmi |
|
|
7 |
SS + HMxy → 2*HMxy |
Kmi |
|
|
8 |
SS + HFxx → 2*HFxx |
Kmi |
|
|
9 |
SS + MMxx → 2*MMxx |
Kmi |
|
|
10 |
SS + MMxy → 2*MMxy |
Kmi |
|
|
11 |
SS + FMxx → 2*FMxx |
Kmi |
|
|
12 |
SS + FMxy → 2*FMxy |
Kmi |
|
|
13 |
SS + FFxx → 2*FFxx |
Kmi |
|
|
Meiosis |
|||
|
14 |
HHxx → 2*Hxe |
Kh |
Oogenesis |
|
15 |
HHxx → 2*Hx |
Kh |
Spermatogenesis |
|
16 |
HMxx → Hxe+ Mxe |
Kh |
O |
|
17 |
HMxx → Hx+ Mx |
Kh |
S |
|
18 |
HMxy → Hx + My |
Kh |
S |
|
19 |
HFxx → Hxe + Fxe |
Kh |
O |
|
20 |
HFxx → Hx + Fx |
Kh |
S |
|
21 |
MMxx → 2*Mx |
Kh |
S |
|
22 |
MMxy → Mx + My |
Ks*Kh |
S |
|
23 |
FMxx → Mxe + Fxe |
Ke*Kh |
O |
|
24 |
FMxy → Fx + My |
Ks*Kh |
S |
|
25 |
FFxx → 2*Fxe |
Ke*Kh |
O |
|
Fertilization |
|||
|
26 |
Hxe + Hx → HHxx |
Kf |
|
|
27 |
Hxe + Mx → HMxx |
Kf |
|
|
28 |
Mxe + Hx → HMxx |
Kf |
|
|
29 |
Hxe + My → HMxy |
Kf |
|
|
30 |
Hxe + Fx → HFxx |
Kf |
|
|
31 |
Fxe + Hx → HFxx |
Kf |
|
|
32 |
Mxe + Mx → MMxx |
Kf |
|
|
33 |
Mxe + My → MMxy |
Kf |
|
|
34 |
Mxe + Fx → FMxx |
Kf |
|
|
35 |
Fxe + Mx → FMxx |
Kf |
|
|
36 |
Fxe + My → FMxy |
Kf |
|
|
37 |
Fxe + Fx → FFxx |
Kf |
|
|
Death |
|||
|
38 |
HHxx → SS |
Kd |
Destruction of zygotes |
|
39 |
HMxx → SS |
Kd |
|
|
40 |
HMxy → SS |
Kd |
|
|
41 |
HFxx → SS |
Kd |
|
|
42 |
MMxx → SS |
Kd |
|
|
43 |
MMxy → SS |
Kd |
|
|
44 |
FMxx → SS |
Kd |
|
|
45 |
FMxy → SS |
Kd |
|
|
46 |
FFxx → SS |
Kd |
|
|
47 |
Hxe → S |
Kde |
Destruction of oocytes |
|
48 |
Hx → S |
Kds |
Destruction of spermatozoa |
|
49 |
Mxe → S |
Kde |
|
|
50 |
Mx → S |
Kds |
|
|
51 |
My → S |
Kds |
|
|
52 |
Fxe → S |
Kde |
|
|
53 |
Fx → S |
Kds |
|
|
54 |
2*S → SS |
Kss |
Substrate regeneration |
The column k→ is the rate constants of the processes. The values of the rate constants are given in Table 2, and the initial conditions - in Table 3.
Table 2. Rate constants
|
Rate constant |
Notes |
|
|
Kmu |
1.00E-04 |
R.C. of mutation |
|
Kmi |
0.1 |
R.C. of mitosis |
|
Kh |
1 |
R.C. of meiosis in hermaphrodites |
|
Ks |
1 |
Acceleration of spermatogenesis M |
|
Ke |
1 |
Acceleration of oogenesis F |
|
Kf |
1.00E+04 |
R.C. of fertilization |
|
Kd |
1 |
R.C. of replicator destruction |
|
Kde |
0.05 |
R.C. of oocyte destruction |
|
Kds |
0.05 |
R.C. of spermatozoa destruction |
|
Kss |
100 |
R.C. of substrate regeneration |
Table 3. Initial conditions
|
Initial conditions |
|
|
|
10 |
|
|
1 |
|
All others |
0 |
 0
0The fact that the average lifespan of replicators was chosen as the time unit is reflected in that, in processes 36-43, the destruction rate constant for all of them is set to Kd=1. The initial value of the number of hermaphrodites
During fertilization, each act involves 1 egg cell and 1 spermatozoon.
As long as the genome contains the dominant gene H, the replicator (HHxx, HMxx, HFxx) is capable, at meiosis, of both oogenesis and spermatogenesis and is a hermaphrodite. Mutation (1) leads to the emergence of gene M, under whose control (and hence, in the absence of genes H or F in the genome) the capacity for oogenesis is lost. Similarly, mutation (2) leads to the emergence of gene F, under whose control (in the absence of genes H or M in the genome) the capacity for spermatogenesis is lost.
Discussion of modeling results
Model 1
As Model 1, the simplest case is considered, when there are no mutations at all (Kmu=0), and the only replicator in the system is the original hermaphrodite HHxx. Fig. 1 shows graphs of the development of equilibrium in this case.
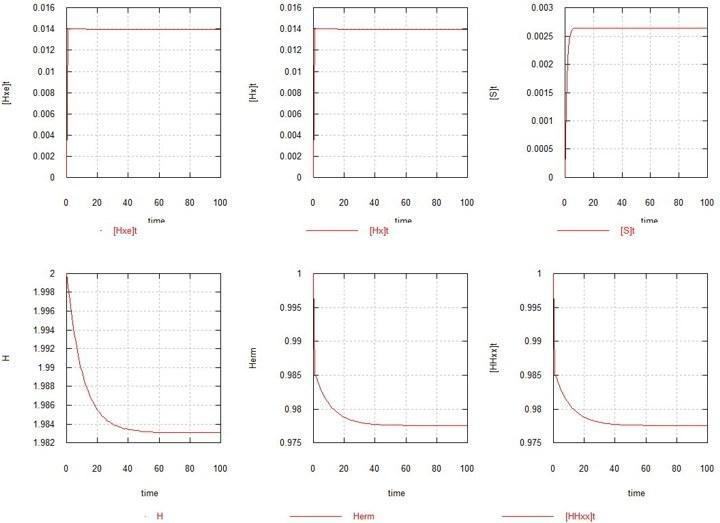
Fig. 1. Development of the steady state in a population of homozygous hermaphrodites.
H – total content of gene H in the system. Herm – number of HHxx zygotes.
As can be seen, and as should be expected, equilibrium (more precisely, a steady state) is reached within a few generations. The number of zygotes decreases by a couple of %, as a result of the gametes reaching a steady-state number of 1.4% relative to the zygotes each. The amount of substrate SS (not shown) practically does not change (about 10).
Model 2
Let us now consider what happens when both mutations (M and F) are switched on. We take the coefficients Ks and Ke equal to 1, i.e. spermatogenesis in M-males and oogenesis in F-females proceed at the same rates as in the original hermaphrodite. We should expect that, since in this situation females lose 50% of their fertility, hermaphrodites should outcompete the emerging mutants. However, contrary to these expectations, what happens is what is depicted in Fig. 2.
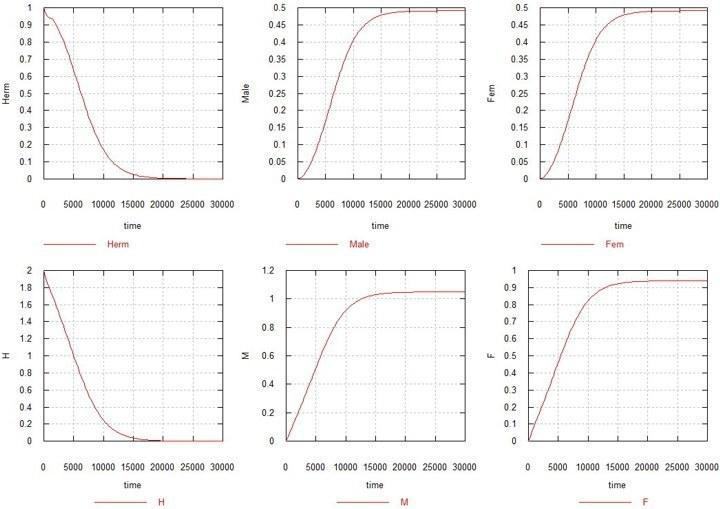
Fig. 2. Evolution of sex under "male" and "female" mutations in a population of hermaphrodites.
H, M and F – total quantities of the respective genes in the system (accounting for their presence in all kinds of zygotes and gametes). Herm – total number of hermaphrodites: HHxx, HMxx and HFxx. Male – total number of males: HMxy, MMxx, MMxy and FMxy. Fem – total number of females: FMxx and FFxx.
As can be seen from the figure, after several tens of thousands of generations, the number of hermaphrodites, along with the total content of the original gene H, falls to zero, and a population is formed consisting exclusively of males and females, each with half the number compared to the original number of hermaphrodites.
I will add (without piling on more figures) that variations of Ks in either direction between values of 0.5 and 1.5 do not lead to a qualitative change in the picture, only somewhat changing the sex ratio.
Model 3
Let us now consider what happens if only the male M-mutation occurs, or only the female F-mutation. First let us leave the male mutation and turn off the female one (exclude processes 2 and 4). Ks is likewise =1.
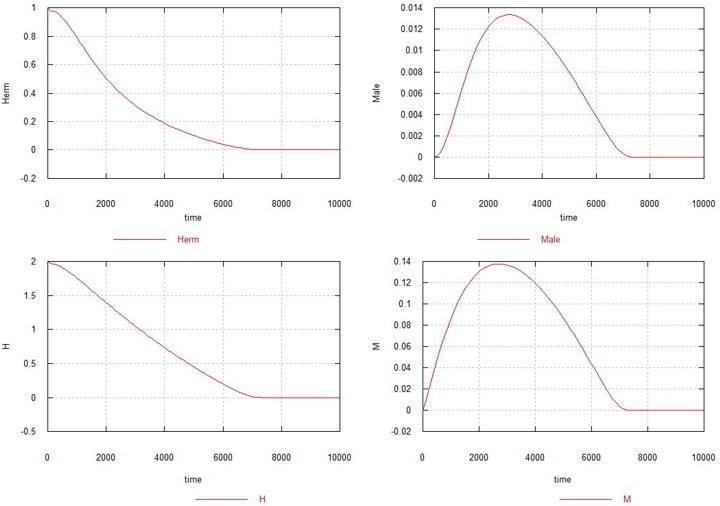
Fig. 3. Dynamics of the emergence of the male mutation in a population of homozygous hermaphrodites. Notation – as before.
Here it turns out that if only the male mutation arises, and it is not followed by the female mutation, then the mutation spreads like a disease, and within a few thousand generations the entire population dies out. Increasing Ks also does not help stabilize the population.
Model 4
A similar situation arises in the case where only the "female" mutation occurs. Let us put process 2 back and exclude processes 1 and 3.
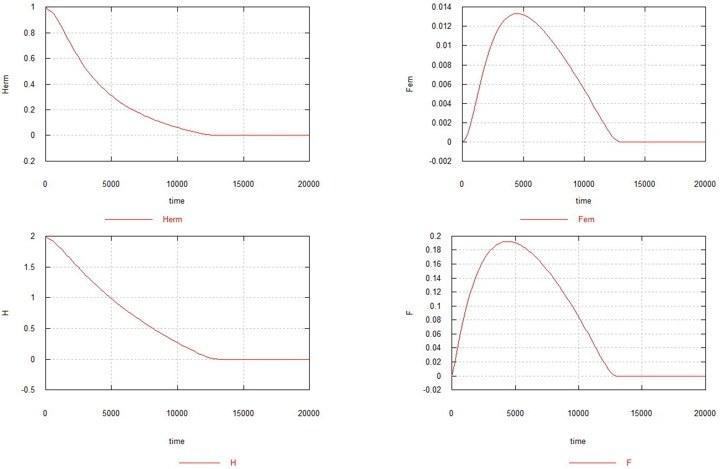
Fig. 4. Dynamics of the emergence of the female mutation in a population of homozygous hermaphrodites. Notation – as before.
As can be seen from the figure, the female mutation in the absence of the male one also leads to the complete extinction of the population, though somewhat more slowly. Increasing Ke to 1.5 in this case (see Fig. 5) leads to the hermaphrodite population not dying out, but sharply shrinking (to 0.7) and a very small steady-state number of females remaining - 0.005. The content of genes H and F stabilizes at the level of 1.6 (the original content was 2.0) and 0.14, respectively.
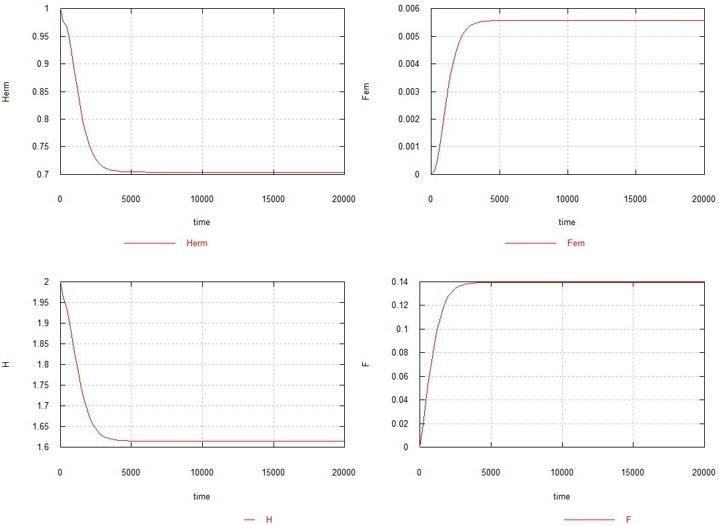
Fig. 5. The conditions are the same as in Fig. 4, with the difference that Ke=1.5.
Conclusions
Contrary to intuitive expectation, if in a population of hermaphrodites the male and female mutations occur synchronously, or with not too great a delay of either one after the other, then, despite the loss of 50% fertility in females and males, the hermaphrodite population over time turns into a population with separate sexes. However, if only one of these mutations occurs, and the other never comes about, this has a detrimental effect on the whole population and reduces its numbers (let us not say it dies out, since the model is crude, and life is a tenacious thing).