
Новая модель Касума Ахмедли
Поскольку обсуждение первой модели Касума Ахмедли стало очень большим, а его новая модель дает новую отправную точку для обсуждения, выношу ее на отдельную страницу.
Оскільки обговорення першої моделі Касума Ахмедлі стало дуже великим, а його нова модель дає нову відправну точку для обговорення, виношу її на окрему сторінку. Ось його лист.
Дорогий Дмитре!
Отже, я склав нову (хіміко-кінетичну) модель, ґрунтуючись на таких припущеннях. Гермафродити мають пару гомологічних хромосом HH, що підтримують одночасно як оогенез, так і сперматогенез (не перешкоджаючи їм). У якийсь момент відбувається мутація, що додає до хромосоми H ділянку, яка блокує оогенез, і перетворює її, таким чином, на хромосому Y. Мутантні особини HY з цієї причини стають самцями. В інший момент у гермафродитів HH виникає інша мутація, що додає до хромосоми H іншу ділянку, з перетворенням її на хромосому X, яка, перебуваючи в гомологічній парі XX, блокує сперматогенез. Оскільки особини HX - як і раніше гермафродити, вони можуть народити самок XX. Таким чином, поєднання HH і HX визначають гермафродитів, HY і XY - це самці, а XX - це самки. Поєднання YY виявляється неможливим, з огляду на те, що немає ні гермафродитів, ні самок з хромосомою Y, які могли б утворити яйцеклітини з нею.
Ця модель багато в чому відрізняється від попередньої. Зокрема, у ній немає димерного субстрату, а лише мономерний S, з кожної одиниці якого може утворитися 1 хромосомна одиниця (1 на гамету, 2 на зиготу) або, відповідно, 1 одиниця якого утворюється при руйнуванні 1 хромосомної одиниці. Крім того, я замінив безперервну мутацію ступінчастою і роблю це вручну, додаючи до системи, близької до рівноважної, передбачуваного мутанта і спостерігаючи за подальшими змінами в ній (на жаль, використовувана програма розрахунку не дозволяє виконувати ступінчасті зміни автоматично). Також я прибрав процеси мітозу, як такі, що не мають прямого стосунку до розмноження. Але натомість яйцеклітини тепер утворюються шляхом включення до їхнього складу запасу субстрату, необхідного для першого поділу після запліднення. Ось як виглядає схема процесів, що відбуваються:
Таблиця 1. Схема процесів
|
N |
Процесс |
k-> |
Примечания |
|---|---|---|---|
|
Мейоз |
|||
|
1 |
HH + 4*S→ 2*SSHe |
Kho |
Оогенез |
|
2 |
HX + 4*S→ SSHe + SSXe |
Kho |
|
|
3 |
XX + 4*S→ 2*SSXe |
Kxo |
|
|
4 |
HH + 4*S→ 6*H |
Khs |
Сперматогенез |
|
5 |
HX + 4*S→ 3*H + 3*X |
Khs |
|
|
6 |
HY + 4*S→ 3*H + 3*Y |
Kys |
|
|
7 |
XY + 4*S→ 3*X + 3*Y |
Kys |
|
|
Оплодотворение |
|||
|
8 |
SSHe + H→ 2*HH |
Kf |
|
|
9 |
SSHe + X→ 2*HX |
Kf |
|
|
10 |
SSHe + Y→ 2*HY |
Kf |
|
|
11 |
SSXe + H→ 2*HX |
Kf |
|
|
12 |
SSXe + X→ 2*XX |
Kf |
|
|
13 |
SSXe + Y→ 2*XY |
Kf |
|
|
Деградация |
|||
|
14 |
HH→ 2*S |
Kd |
|
|
15 |
HX→ 2*S |
Kd |
|
|
16 |
HY→ 2*S |
Kd |
|
|
17 |
XX→ 2*S |
Kd |
|
|
18 |
XY→ 2*S |
Kd |
|
|
19 |
SSHe→ 3*S |
Kdh |
|
|
20 |
SSXe→ 3*S |
Kdx |
|
|
21 |
H→ S |
Kds |
|
|
22 |
X→ S |
Kds |
|
|
23 |
Y→ S |
Kds |
Сперматогенез, як бачите, теж іде зі споживанням тієї ж кількості субстрату, що й оогенез, але цей субстрат не включається в сперматозоїди як запас, а витрачається на виробництво більшої кількості сперматозоїдів, ніж яйцеклітин. Решта процесів, мені здається, не потребують роз'яснень.
Як і раніше, за одиницю кількості матеріалу (1 P) прийнято початкову кількість клітин з набором HH, а за одиницю часу (1 L) - середню тривалість життя клітини HH (яка не зазнала мейозу).
У табл. 2 наведено вибрані значення констант швидкостей процесів.
Таблиця 2. Константи швидкостей
|
Процеси |
Константи швидкості |
|
|
Оогенез гермафродитов |
Kho |
1.00E-04 |
|
Оогенез у самок |
Kxo |
1.70E-04 |
|
Сперматогенез у гермафродитов |
Khs |
7.00E-05 |
|
Сперматогенез у самцов |
Kys |
1.70E-04 |
|
Оплодотворение |
Kf |
1.00E+04 |
|
Деградация зигот |
Kd |
1 |
|
Деградация ооцитов гермафродитов |
Kdh |
100 |
|
Деградация ооцитов самок |
Kdx |
50 |
|
Деградация сперматозоидов |
Kds |
100 |
Таблиця 3. Початкові умови
|
|
10 |
|
|
1 |
 0
0Прокоментую вибрані значення констант швидкостей. Значення Kd=1 випливає з угоди про одиницю часу. Це означає, що 1 диплоїдна клітина руйнується в середньому за час 1 L. Гаплоїдні клітини руйнуються в 100 разів швидше (за 0.01 L), що забезпечує відповідно нижчу концентрацію гамет. Для ооцитів самок вибрано вдвічі більший час життя, ніж для ооцитів гермафродитів, що, імовірно, досягається ними за рахунок економії сил, не витрачуваних на виробництво сперматозоїдів, і виглядає логічно. Константи швидкостей мейозу й запліднення вибрані так, щоб при вибраних початкових умовах швидкості мейозу й запліднення були того ж порядку, що й швидкість деградації зигот. Швидкість сперматогенезу гермафродитів вибрана на 30% нижче швидкості оогенезу в них же. А константа швидкості сперматогенезу в самців дорівнює сумі констант обох видів мейозу в гермафродитів, що означає, що й гермафродити, і самці витрачають на мейоз однакові сили, але дає самцям чисельну перевагу сперматозоїдів. Самки виробляють ооцити з такою ж швидкістю, що й гермафродити, а те, що вони вкладають при цьому стільки ж сил, проявляється в більшій тривалості життя (більшій життєздатності) їхніх ооцитів.
Перейдемо тепер до результатів моделювання.
Раунд 1
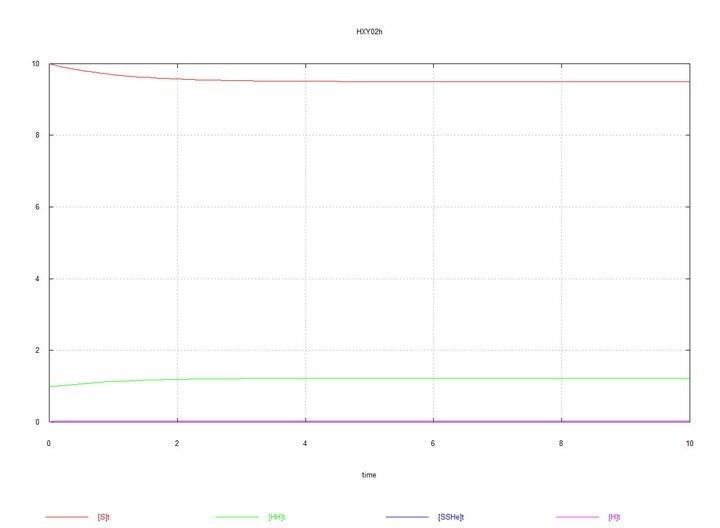
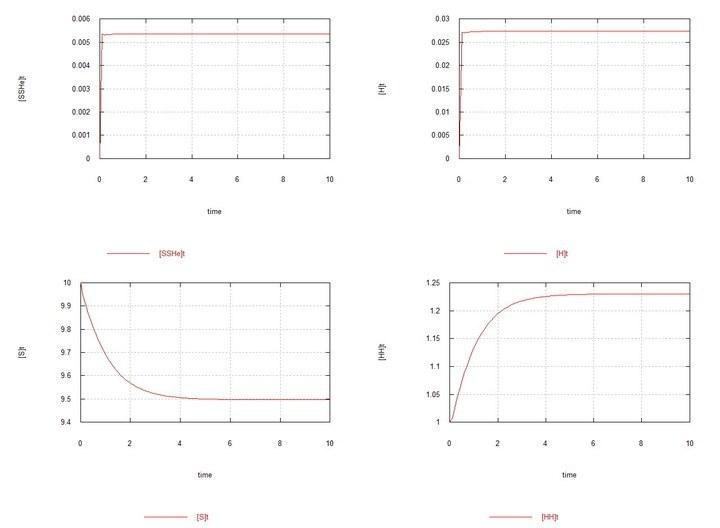
Рис. 1 (Fig01h) і Рис. 2 (Fig02h) – Рівновага в системі гермафродитів HH.
Графіки системи, що складається з самих гермафродитів HH, наводжу тільки з метою початкового контролю – щоб бачити рівноважні концентрації зигот і гамет, що встановлюються в системі.
Раунд 2
У систему гермафродитів, де
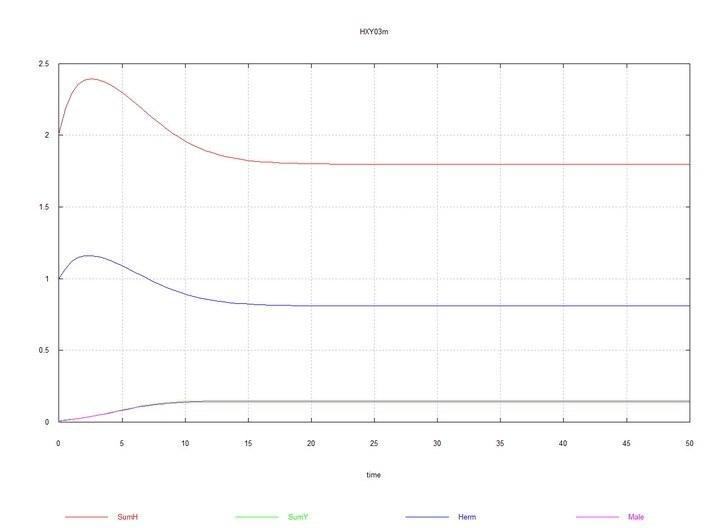
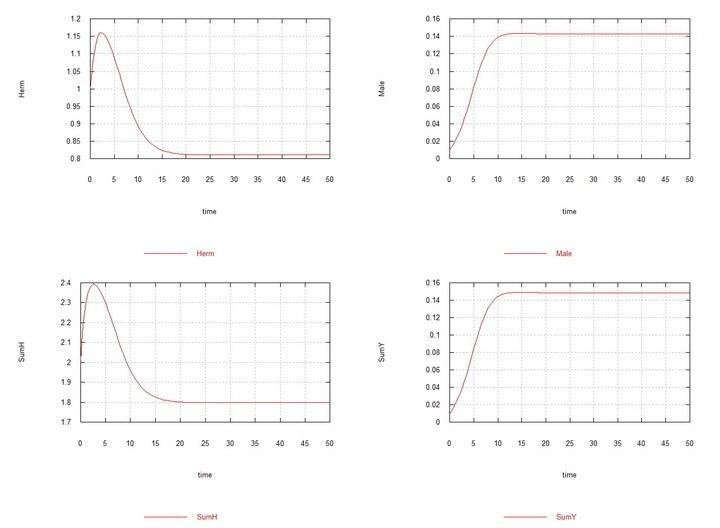
Рис. 3 і 4. (Fig03m і Fig04m) – Кінетичні криві при додаванні в середовище гермафродитів 1% мутантів-самців HY. SumH = 2*
Тут, як було сказано вище й показано в табл. 2, константа швидкості сперматогенезу самців Kys прийнята рівною 1.7e-4. Якщо її залишити рівною 1e-4, тобто такою ж, як у гермафродитів, то самці й хромосома Y невдовзі повністю витісняються гермафродитами HH.
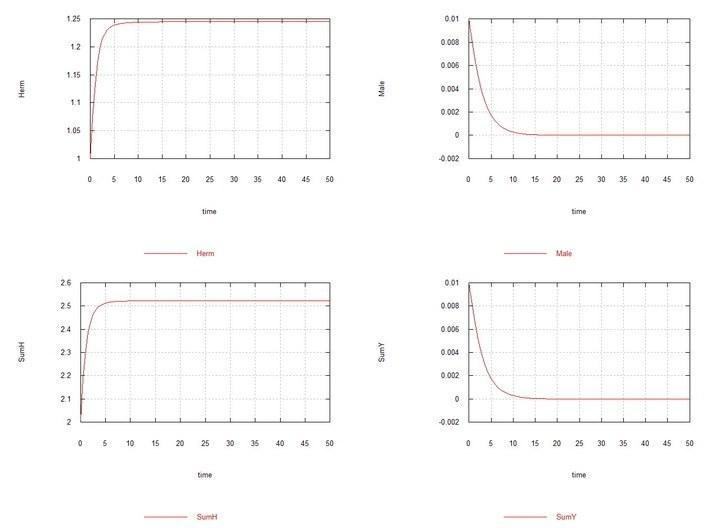
Рис. 5. (Fig09m) – Умови аналогічно Fig04m з тією відмінністю, що Kys вибрано рівним 7e-5, тобто константи швидкості сперматогенезу в гермафродитів HH і самців HY однакові. Демонструється витіснення самців за цих умов.
Раунд 3
У систему гермафродитів, де
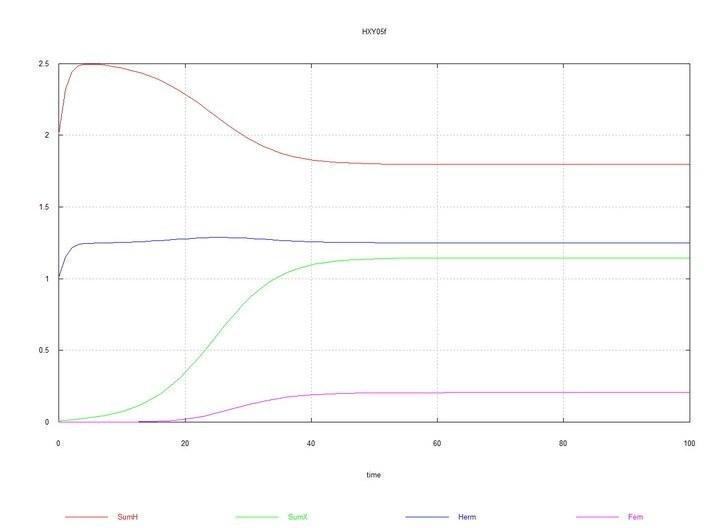
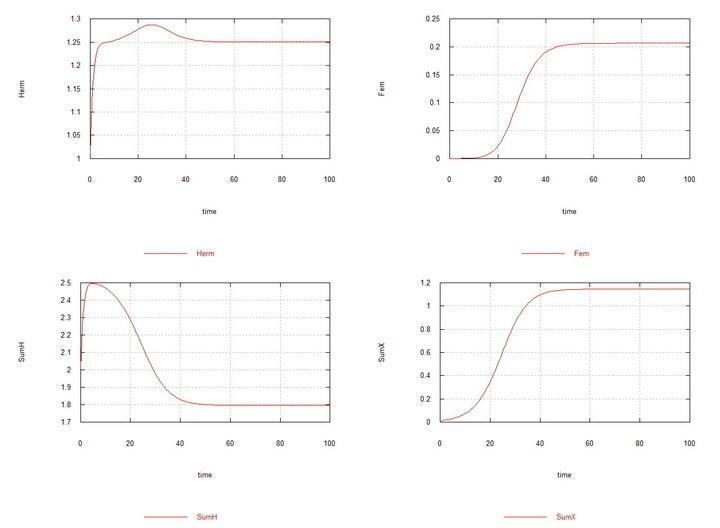
Рис. 6 і 7. (Fig05f і Fig06f) – Кінетичні криві при додаванні в середовище гермафродитів 1% мутантів-гермафродитів HX. SumH = 2*
Тут константа Kdx швидкості деградації ооцитів самок SSXe вибрана рівною 50 (табл. 2). Якщо ж вибрати її рівною 100, як для ооцитів гермафродитів SSHe, то й у цьому випадку хромосома X невдовзі повністю витісняється хромосомою H.
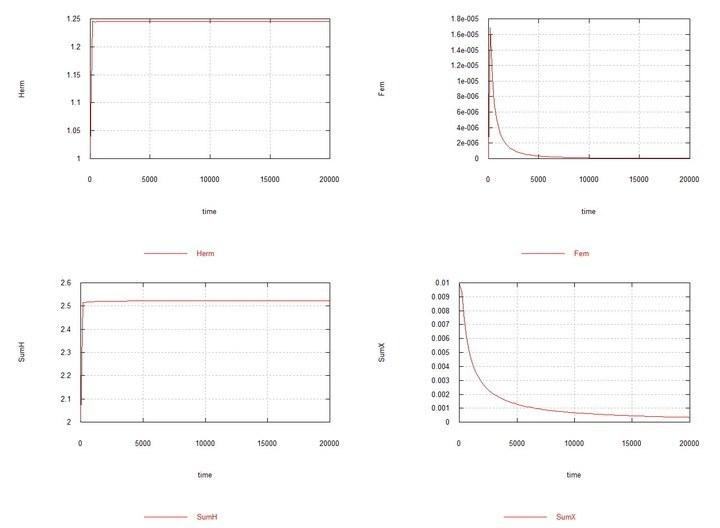
Рис. 8. Fig10f – Умови аналогічно Fig06f з тією відмінністю, що Kdx вибрано рівним 100, тобто константи швидкості деградації ооцитів у гермафродитів і самок однакові. Демонструється витіснення самок за цих умов.
Раунд 4
В навколо-рівноважну систему гермафродити- самці зі співвідношенням
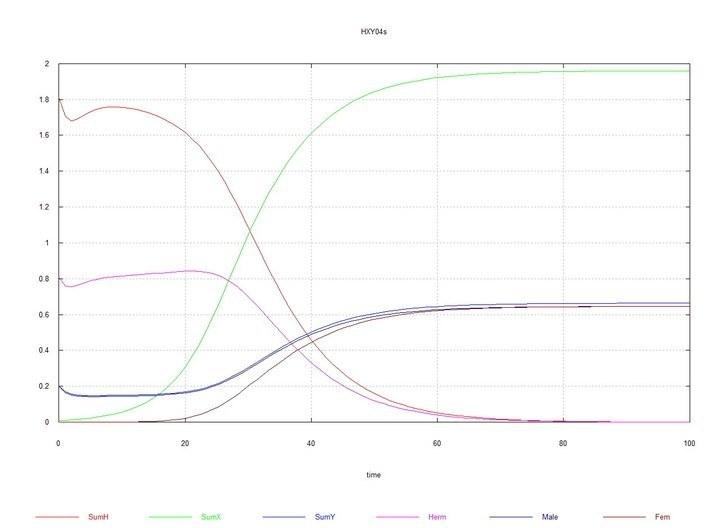
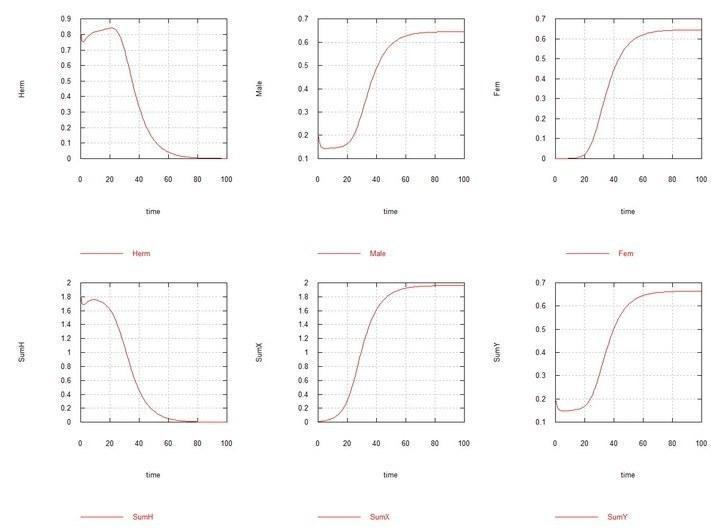
Рис. 9 і 10 (Fig07s і Fig08s) – Кінетичні криві при додаванні 1% гермафродитів HX у рівноважну систему гермафродитів (HH 80%) і самців (HY 20%). SumH = 2*
Висновки
У результаті моделювання вийшло, що поява в популяції гермафродитів HH мутанта HY, який має колишню швидкість сперматогенезу (тобто як у HH), мутанти швидко витісняються гермафродитами. Однак, якщо самці HY витрачають на сперматогенез стільки ж сил, скільки гермафродит на обидва види гамет сумарно, то встановлюється деяка рівновага, у якій, усе ж, гермафродити значно переважають у чисельності самців. Це очікувано, оскільки самці у своєму потомстві мають рівне число самців і гермафродитів, а гермафродити між собою мають ще й додаткову кількість нащадків-гермафродитів.
При появі в популяції гермафродитів HH мутантних гермафродитів HX, і наданні самкам XX такої ж здатності виробляти яйцеклітини, як і гермафродитам HH і HX, хромосома X так само швидко витісняється хромосомою H, як це відбувалося зі слабкою хромосомою Y. Але якщо самкам XX надається здатність виробляти більш життєздатні ооцити (за рахунок того, що вони витрачають на їх виробництво стільки ж сил, скільки гермафродити витрачають на виробництво обох типів гамет у сумі), то й тут встановлюється рівновага, у якій чисельно переважають гермафродити (і, як легко бачити, з тих же причин).
Однак, коли в рівноважній системі гермафродити-самці HH-HY (з переважанням, як було сказано, гермафродитів HH), з'являється мутант HX, то відбувається дивергентне зростання чисельності самців XY і самок XX, повне витіснення гермафродитів усіх видів і самої хромосоми H, і встановлюється рівновага з рівною чисельністю самців і самок.
P.S. Можливо, замість розмови про H, X, Y, як про хромосоми, слід було б говорити про них, як про алелі одного гена, як робите це Ви, але я виходив з того, що гени, що визначають чоловічу й жіночу стать – це різні гени, і ген на хромосомі Y антагоністичний до самого себе й не здатний до утворення пари YY. Утім, усе це для мене не дуже ясно, а Ви, як фахівець у цій галузі, можете судити набагато компетентніше.