Kasum Akhmedli's new model
Since the discussion of Kasum Ahmedli's first model has become very extensive, and his new model provides a new starting point for discussion, I am placing it on a separate page.
Since the discussion of Kasum Ahmedli's first model became very extensive, and his new model provides a new starting point for discussion, I am presenting it on a separate page. Here is his letter.
Dear Dmytro!
So, I have compiled a new (chemical-kinetic) model, based on the following assumptions. Hermaphrodites have a pair of homologous chromosomes HH that support both oogenesis and spermatogenesis simultaneously (not interfering with them). At some point, a mutation occurs, adding to chromosome H a segment that blocks oogenesis, thus transforming it into chromosome Y. Mutant HY individuals become males for this reason. At another point, another mutation occurs in hermaphrodites HH, adding to chromosome H another segment, transforming it into chromosome X, which, when in homologous pair XX, blocks spermatogenesis. Since HX individuals are still hermaphrodites, they can produce XX females. Thus, combinations HH and HX determine hermaphrodites, HY and XY are males, and XX are females. The combination YY turns out to be impossible, since there are neither hermaphrodites nor females with chromosome Y that could produce eggs with it.
This model differs from the previous one in many respects. In particular, it has no dimeric substrate, but only monomeric S, from each unit of which 1 chromosome unit (1 per gamete, 2 per zygote) can form, or, respectively, 1 unit forms from the destruction of 1 chromosome unit. Furthermore, I replaced continuous mutation with step-wise mutation and do it manually, adding a presumed mutant to a system close to equilibrium and observing further changes in it (unfortunately, the calculation program used does not allow automatic step-wise changes). I also removed mitosis processes as having no direct relation to reproduction. But instead, eggs now form by including in their composition a reserve of substrate necessary for the first division after fertilization. Here is the scheme of the processes taking place:
Table 1. Scheme of Processes
|
N |
Process |
k-> |
Notes |
|---|---|---|---|
|
Meiosis |
|||
|
1 |
HH + 4*S→ 2*SSHe |
Kho |
Oogenesis |
|
2 |
HX + 4*S→ SSHe + SSXe |
Kho |
|
|
3 |
XX + 4*S→ 2*SSXe |
Kxo |
|
|
4 |
HH + 4*S→ 6*H |
Khs |
Spermatogenesis |
|
5 |
HX + 4*S→ 3*H + 3*X |
Khs |
|
|
6 |
HY + 4*S→ 3*H + 3*Y |
Kys |
|
|
7 |
XY + 4*S→ 3*X + 3*Y |
Kys |
|
|
Fertilization |
|||
|
8 |
SSHe + H→ 2*HH |
Kf |
|
|
9 |
SSHe + X→ 2*HX |
Kf |
|
|
10 |
SSHe + Y→ 2*HY |
Kf |
|
|
11 |
SSXe + H→ 2*HX |
Kf |
|
|
12 |
SSXe + X→ 2*XX |
Kf |
|
|
13 |
SSXe + Y→ 2*XY |
Kf |
|
|
Degradation |
|||
|
14 |
HH→ 2*S |
Kd |
|
|
15 |
HX→ 2*S |
Kd |
|
|
16 |
HY→ 2*S |
Kd |
|
|
17 |
XX→ 2*S |
Kd |
|
|
18 |
XY→ 2*S |
Kd |
|
|
19 |
SSHe→ 3*S |
Kdh |
|
|
20 |
SSXe→ 3*S |
Kdx |
|
|
21 |
H→ S |
Kds |
|
|
22 |
X→ S |
Kds |
|
|
23 |
Y→ S |
Kds |
As you can see, spermatogenesis also proceeds with consumption of the same amount of substrate as oogenesis, but this substrate is not included in spermatozoa as a reserve, but is spent on producing a greater number of spermatozoa than eggs. The other processes, it seems to me, do not require explanation.
As before, the initial amount of cells with set HH is taken as the unit of material quantity (1 P), and the average lifespan of an HH cell (that has not undergone meiosis) is taken as the unit of time (1 L).
Table 2 presents the selected values of the rate constants for the processes.
Table 2. Rate Constants
|
Processes |
Rate constants |
|
|
Hermaphrodites oogenesis |
Kho |
1.00E-04 |
|
Females oogenesis |
Kxo |
1.70E-04 |
|
Hermaphrodites spermatogenesis |
Khs |
7.00E-05 |
|
Males spermatogenesis |
Kys |
1.70E-04 |
|
Fertilization |
Kf |
1.00E+04 |
|
Zygote degradation |
Kd |
1 |
|
Hermaphrodites oocyte degradation |
Kdh |
100 |
|
Females oocyte degradation |
Kdx |
50 |
|
Spermatozoa degradation |
Kds |
100 |
I will comment on the selected values of the rate constants. The value Kd=1 follows from the convention about the unit of time. This means that 1 diploid cell is destroyed on average during time 1 L. Haploid cells are destroyed 100 times faster (in 0.01 L), which ensures a correspondingly lower concentration of gametes. For female oocytes, a twice longer lifespan was chosen than for hermaphrodite oocytes, which, presumably, is achieved by them due to saving energy not spent on spermatozoa production, and seems logical. The rate constants of meiosis and fertilization were chosen so that with the selected initial conditions, the rates of meiosis and fertilization are of the same order as the rate of zygote degradation. The rate of hermaphrodites spermatogenesis is chosen 30% lower than the rate of oogenesis in them. And the rate constant of spermatogenesis in males equals the sum of the constants of both types of meiosis in hermaphrodites, which means that both hermaphrodites and males spend the same effort on meiosis, but gives males a numerical advantage in spermatozoa. Females produce oocytes at the same rate as hermaphrodites, and the fact that they invest the same effort in this is manifested in the longer lifespan (greater viability) of their oocytes.
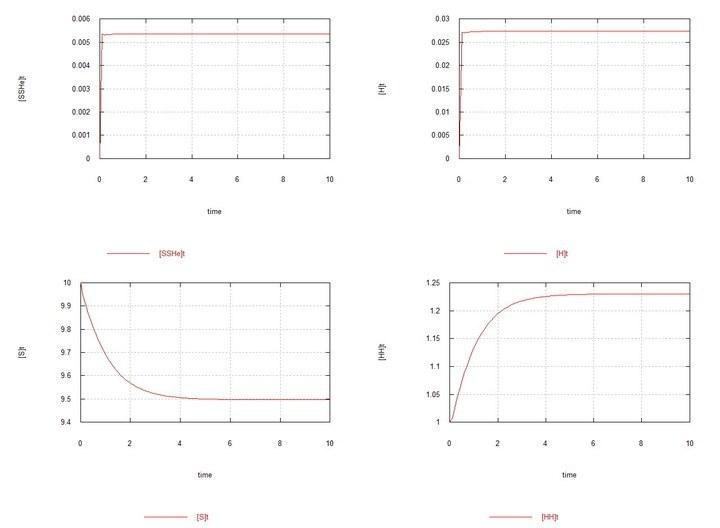
Now let us proceed to the results of the modeling.
|
S
0
|
10 |
|
HH
0
|
1 |
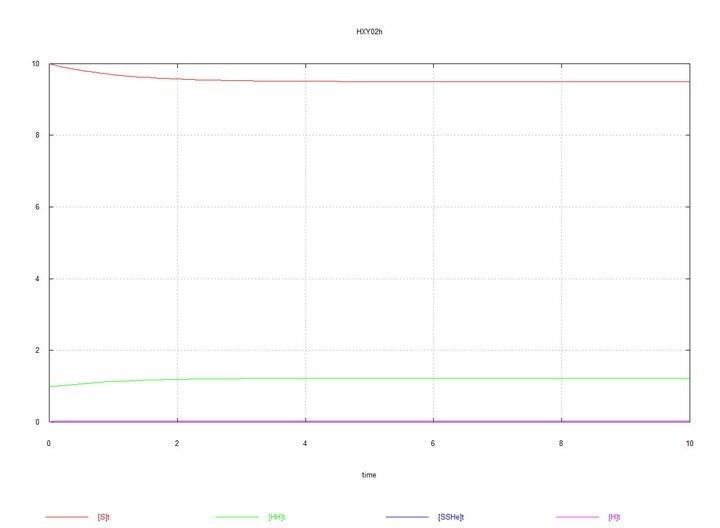
To the system of hermaphrodites, where  S
S 0 = 10 and HH0 = 1, HY0 = 0.01 is added. The system moves toward equilibrium with a predominance of hermaphrodites HH and an equilibrium concentration of males HY of about 15%.
0 = 10 and HH0 = 1, HY0 = 0.01 is added. The system moves toward equilibrium with a predominance of hermaphrodites HH and an equilibrium concentration of males HY of about 15%.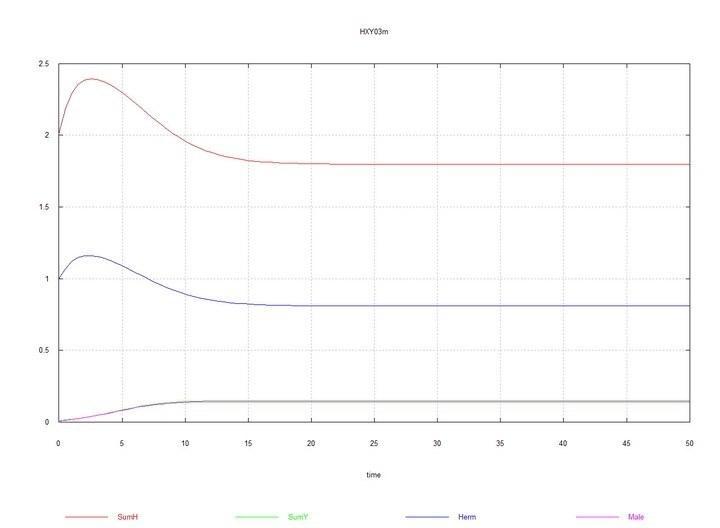
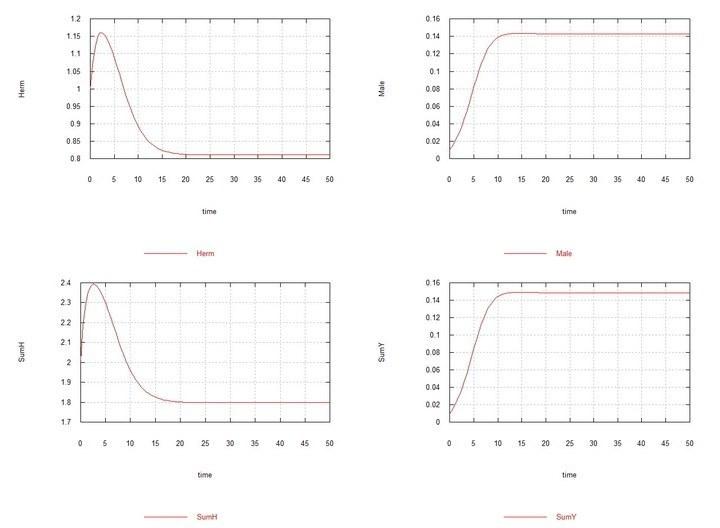
Figs. 3 and 4. (Fig03m and Fig04m) – Kinetic curves when adding 1% mutant males HY to the hermaphrodite medium. SumH = 2*HH + HY + SSHe + H; SumY = HY + Y; Herm = HH; Male = HY.
Here, as stated above and shown in Table 2, the rate constant of male spermatogenesis Kys is taken as 1.7e-4. If it is left equal to 1e-4, i.e., the same as in hermaphrodites, then males and chromosome Y are soon completely displaced by hermaphrodites HH.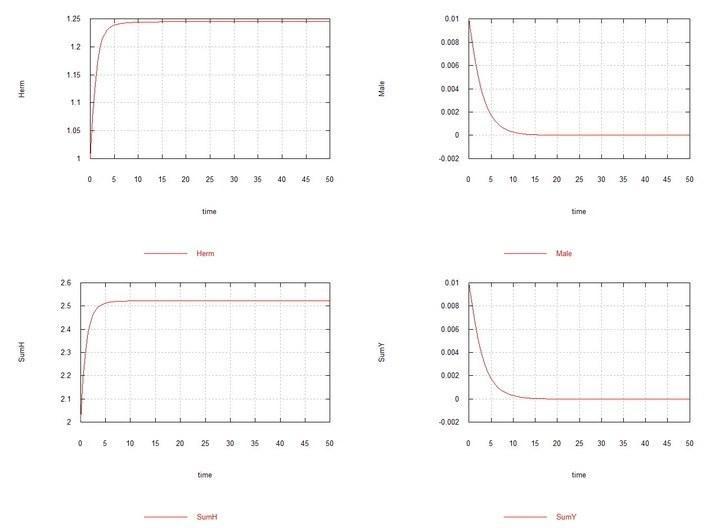
Fig. 5. (Fig09m) – Conditions are similar to Fig04m with the difference that Kysis chosen equal to 7e-5, i.e., the rate constants of spermatogenesis in hermaphrodites HHand males HYare the same. Displacement of males under these conditions is demonstrated.
To the system of hermaphrodites, where S0 = 10 and HH0 = 1, HX0 = 0.01 is added. The system moves toward equilibrium with a predominance of hermaphrodites and an equilibrium concentration of females FF of about 20%.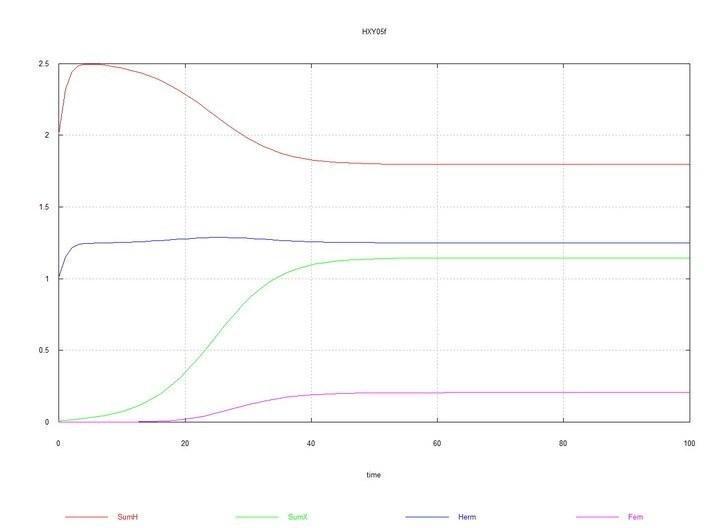
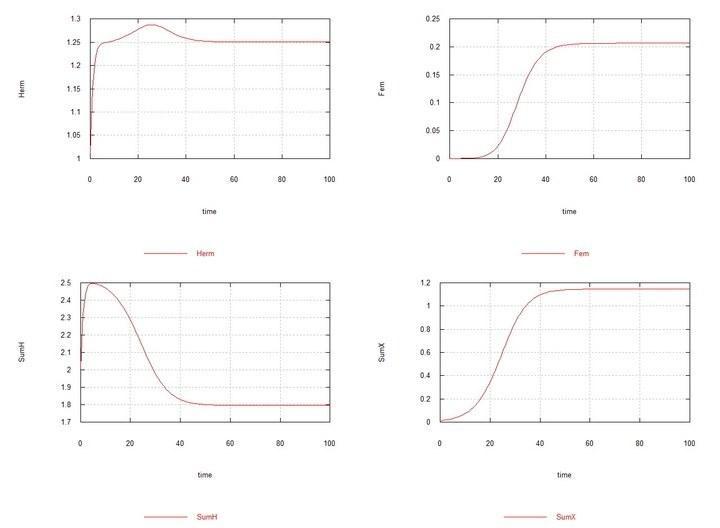
Figs. 6 and 7. (Fig05f and Fig06f) – Kinetic curves when adding 1% mutant hermaphrodites HXto the hermaphrodite medium. SumH = 2*HH + HX + SSHe + H; SumX = HX + 2*XX + SSXe + X; Herm = HH + HX; Fem = XX.
Here the constant Kdx of the rate of degradation of female oocytes SSXe is chosen equal to 50 (Table 2). However, if it is chosen equal to 100, as for hermaphrodite oocytes SSHe, then in this case too chromosome X is soon completely displaced by chromosome H.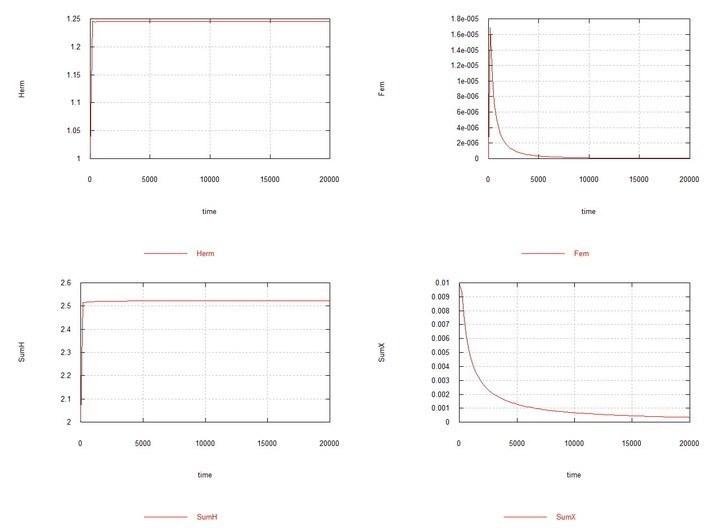
Fig. 8. Fig10f – Conditions are similar to Fig06f with the difference that Kdx is chosen equal to 100, i.e., the rate constants of oocyte degradation in hermaphrodites and females are the same. Displacement of females under these conditions is demonstrated.
To the near-equilibrium system of hermaphrodites-males with the ratio HH0 = 0.8, HY = 0.2, HX0 = 0.01 is added. Males XYand females XXgradually completely displace hermaphrodites of both types and males HY.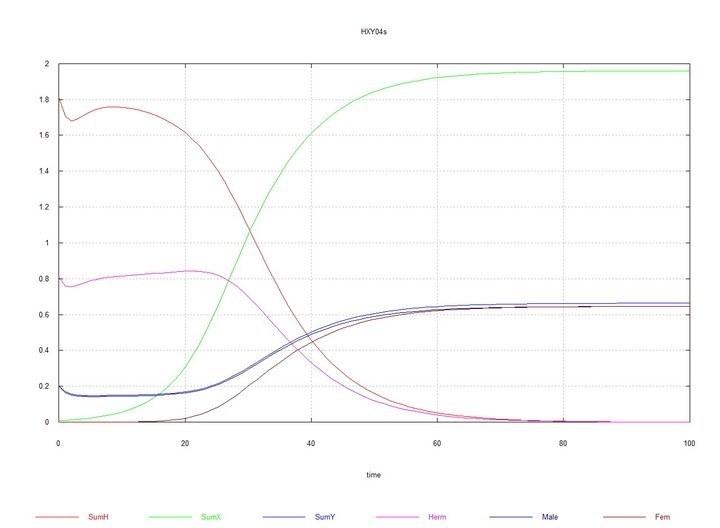
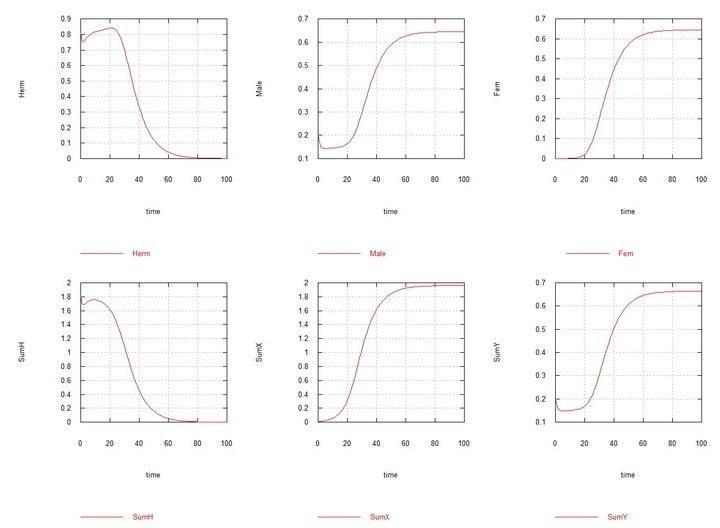
Figs. 9 and 10 (Fig07s and Fig08s) – Kinetic curves when adding 1% hermaphrodites HXto the equilibrium system of hermaphrodites (HH80%) and males (HY20%). SumH = 2*HH + HX + HY + SSHe + H; SumX = HX + 2*XX + XY + SSXe + X ; SumY = HY + XY + Y; Herm = HH + HX; Male = HY + XY; Fem = XX.
As a result of modeling, it turned out that the appearance in the population of hermaphrodites HHof the mutant HYwith the same rate of spermatogenesis (i.e., as in HH) leads to the mutants being rapidly displaced by hermaphrodites. However, if males HYspend on spermatogenesis as much effort as a hermaphrodite spends on both types of gametes combined, then some equilibrium is established, in which, nevertheless, hermaphrodites significantly outnumber males. This is expected, since, males in their offspring have an equal number of males and hermaphrodites, and hermaphrodites among themselves also have an additional number of hermaphrodite offspring.
When mutant hermaphrodites HXappear in the population of hermaphrodites HH, and females XXare given the same ability to produce eggs as hermaphrodites HHand HX, chromosome Xis also as rapidly displaced by chromosome H, as occurred with the weak chromosome Y. But if females XXare given the ability to produce more viable oocytes (due to the fact that they spend on their production as much effort as hermaphrodites spend on producing both types of gametes combined), then here too an equilibrium is established, in which hermaphrodites predominate numerically (and, for the same reasons, as is easy to see).
However, when in the equilibrium system hermaphrodites-males HH-HY (with predominance, as was stated, of hermaphrodites HH), mutant HXappears, then there is a divergent increase in the number of males XYand females XX, complete displacement of hermaphrodites of all types and of chromosome H itself, and equilibrium is established with equal numbers of males and females.
P.S. Perhaps, instead of talking about H, X, Yas chromosomes, one should speak of them as alleles of a single gene, as you do, but I proceeded from the fact that the genes determining male and female sex are different genes, and the gene on chromosome Yis antagonistic to itself and unable to form a YY pair. However, all this is not very clear to me, and you, as a specialist in this field, can judge much more competently.
|
|
10 |
|
|
1 |
I will comment on the chosen values of the rate constants. The value Kd=1 follows from the convention of the time unit. This means that 1 diploid cell is destroyed on average in 1 L. Haploid cells are destroyed 100 times faster (in 0.01 L), which ensures a correspondingly lower concentration of gametes. For female oocytes, a lifespan twice as long as for hermaphrodite oocytes is chosen, which is presumably achieved by saving energy not spent on sperm production, and seems logical. The rate constants for meiosis and fertilization are chosen such that, under the chosen initial conditions, the rates of meiosis and fertilization are of the same order as the rate of zygote degradation. The rate of spermatogenesis in hermaphrodites is chosen to be 30% lower than the rate of oogenesis in them. And the rate constant for spermatogenesis in males is equal to the sum of the constants for both types of meiosis in hermaphrodites, which means that both hermaphrodites and males spend the same energy on meiosis, but gives males a numerical advantage in sperm. Females produce oocytes at the same rate as hermaphrodites, and the fact that they invest the same energy in this is reflected in the longer lifespan (greater viability) of their oocytes. Let's move on to the simulation results. Round 1 Fig. 1 (Fig01h) and Fig. 2 (Fig02h) – Equilibrium in the HH hermaphrodite system. I present the graphs of the system consisting only of HH hermaphrodites solely for initial control – to see the equilibrium concentrations of zygotes and gametes established in the system. Round 2 To the hermaphrodite system, where S 0 = 10 and HH 0 = 1, HY 0 = 0.01 is added. The system moves towards equilibrium with a predominance of HH hermaphrodites and an equilibrium concentration of HY males of about 15%. Figs. 3 and 4. (Fig03m and Fig04m) – Kinetic curves upon addition of 1% male mutants HY to the hermaphrodite environment. SumH = 2* HH + HY + SSHe + H ; SumY = HY + Y ; Herm = HH ; Male = HY . Here, as stated above and shown in Table 2, the male spermatogenesis rate constant Kys is taken as 1.7e-4. If it is kept at 1e-4, i.e., the same as for hermaphrodites, then males and the Y chromosome are soon completely displaced by HH hermaphrodites. Fig. 5. (Fig09m) – Conditions are analogous to Fig04m, with the difference that Kys is chosen as 7e-5, i.e., the spermatogenesis rate constants for HH hermaphrodites and HY males are the same. The displacement of males under these conditions is demonstrated. Round 3 To the hermaphrodite system, where S 0 = 10 and HH 0 = 1, HX 0 = 0.01 is added. The system moves towards equilibrium with a predominance of hermaphrodites and an equilibrium concentration of FF females of about 20%. Figs. 6 and 7. (Fig05f and Fig06f) – Kinetic curves upon addition of 1% HX hermaphrodite mutants to the hermaphrodite environment. SumH = 2* HH + HX + SSHe + H ; SumX = HX + 2* XX + SSXe + X ; Herm = HH + HX ; Fem = XX . Here, the oocyte degradation rate constant Kdx for female SSXe is chosen as 50 (Table 2). If it is chosen as 100, as for hermaphrodite oocytes SSHe, then in this case too, the X chromosome is soon completely displaced by the H chromosome. Fig. 8. Fig10f – Conditions are analogous to Fig06f, with the difference that Kdx is chosen as 100, i.e., the oocyte degradation rate constants for hermaphrodites and females are the same. The displacement of females under these conditions is demonstrated. Round 4 In a near-equilibrium hermaphrodite-male system with HH 0 = 0.8, HY = 0.2, HX 0 = 0.01 is added. XY males and XX females gradually completely displace hermaphrodites of all types and HY males. Figs. 9 and 10 (Fig07s and Fig08s) – Kinetic curves upon addition of 1% HX hermaphrodites to an equilibrium system of hermaphrodites (HH 80%) and males (HY 20%). SumH = 2* HH + HX + HY + SSHe + H ; SumX = HX + 2* XX + XY + SSXe + X ; SumY = HY + XY + Y ; Herm = HH + HX ; Male = HY + XY ; Fem = XX . Conclusions As a result of the modeling, it was found that when the HH hermaphrodite population is invaded by the HY mutant, which has the same spermatogenesis rate (i.e., as HH), the mutants are rapidly displaced by hermaphrodites. However, if HY males spend as much energy on spermatogenesis as a hermaphrodite spends on both types of gametes combined, then some equilibrium is established, in which hermaphrodites still significantly outnumber males. This is expected, since males in their offspring have an equal number of males and hermaphrodites, while hermaphrodites among themselves have an additional number of hermaphrodite offspring. When the HH hermaphrodite population is invaded by the HX mutant hermaphrodites, and XX females are given the same ability to produce oocytes as HH and HX hermaphrodites, the X chromosome is displaced by the H chromosome as rapidly as the weak Y chromosome was. But if XX females are given the ability to produce more viable oocytes (because they spend as much energy on their production as hermaphrodites spend on the production of both types of gametes combined), then an equilibrium is also established here, in which hermaphrodites numerically predominate (and, as is easily seen, for the same reasons). However, when a HX mutant appears in an equilibrium system of HH-HY hermaphrodites-males (with a predominance, as mentioned, of HH hermaphrodites), then a divergent increase in the number of XY males and XX females occurs, the complete displacement of hermaphrodites of all types and the H chromosome itself, and an equilibrium with an equal number of males and females is established. P.S. Perhaps, instead of talking about H, X, Y as chromosomes, it would be better to talk about them as alleles of the same gene, as you do, but I proceeded from the assumption that the genes determining male and female sex are different genes, and the gene on the Y chromosome is antagonistic to itself and incapable of forming a YY pair. However, all this is not very clear to me, and you, as a specialist in this field, can judge much more competently.