
Подкласс Parareptilia
Парарептилії (Parareptilia) — велика група наземних чотириногих [4], збірна, можливо штучна група [3], імовірно, незалежно від решти хребетних, що досягла рептилійного рівня організації. Також, згідно з деякими уявленнями, від парарептилій ведуть свій початок черепахи. [4]
Систематика
Термін Парарептилії (Parareptilia) запропонований Е. Олсоном (американський палеонтолог) у 1947 році. Різні класифікації дають різний склад для цієї групи. [3] Для початку можна сказати, що нині сеймуріаморф і діадектів зазвичай відносять до амфібій, а також більшість сучасних авторів зближує сеймуріаморф з пареязаврами. [1]
Керролл (американський учений, доктор наук у галузі палеонтології хребетних, що спеціалізується на палеозойських і мезозойських земноводних і плазунах) схильний розглядати парарептилій як сестринську групу амніот, поміщаючи їх вище вестлотіани. Ватсон (англійський палеонтолог, написав багато праць з еволюції викопних хребетних) розділяв найпримітивніших рептилій без скроневих вікон або котилозаврів (Cotilosauria), на три підряди: Seymouriamorpha, Diadectomorfa і Captorhinomorpha. У діадектоморф він включив діадектів і проколофонів, що мали вушну вирізку, і перейазаврів, що вторинно втратили вушну вирізку. Діадекти поміщалися в основі діадектоморфних котилозаврів, які пізніше складали ядро парарептилій при їх виділенні. [1]
За Е. Олсоном парарептилії спочатку об'єднували сеймуріаморф, діадектів, проколофонів, пайеязаврів і черепах. Сеймуріаморфи, поставлені в ранг підкласу, були з'єднані з діадектами, дві інші групи діадектоморфних котилозаврів – проколофони і пареязаври – виділялися окремо. Олсон вважав черепах ближчими до діадектів, а не до парейазаврів. Парарептилії протиставлялися еврептиліям, що включали всіх інших рептилій. Сеймуріаморфи, за Олсоном, вважалися предками діадектів. [1]
Пізніше Олсон виділяв сеймуріаморф в особливий клас Batrachosauria і розглядав як парарептилій тільки проколонофорів і пайеязаврів. Пізніше він узагалі відмовився від концепції парарептилій, з яких послідовно, услід за черепахами, були виключені сеймуріаморфи і діадекти. [1]
Ф. Хюне (німецький палеонтолог, займався вивченням викопних плазунів) об'єднав усіх парарептилій з антракозаврами у вигляді особливої гілки еволюції тетрапод – Reptiliomorpha. До цієї гілки він включає і черепах, яких він виводить від діадектоморф. У цілому він виводив усіх рептилій від рептиліоморф, але парарептилій він виділяв в окрему від рептилій гілку еволюції. [1]
Ромер (американський палеонтолог і морфолог, автор підручників з палеонтології, анатомії хребетних) розглядає сеймуріаморф разом з діадектами у складі амфібій, а проколофонів і пайеязаврів об'єднує в підряді Procolophonia ряду котилозаврів. Туди ж він відносить і міллерозаврів. [1]
М. Хітон возз'єднав сеймуріаморф з діадектами в загальному ряді Cotylosauria і переніс його в клас амфібій. Він акцентував увагу на тій обставині, що ряд спочатку був виведений на основі роду Diadectes=Empedocles. Проколофонів, парейазаврів і капториноморф. Хітон залишив у рептиліях, але виключив їх зі складу котилозаврів. Котилозаврів, виділюваних у підклас Batrachosauria він розділяє на сеймуріаморф і діадектоморф. Хітон виключає зі складу сеймуріаморф родину Discosauriscidae, представники якої вели амфібійний спосіб життя. Лантанозух у числі батрахозаврів не розглядається. У число батрахозаврів Хітон включає Nycteroleteridae. [1]
Готьє (палеонтолог Єльського університету), Клюге і Роу (професор палеонтології з Техаського університету) виділяють парарептилій, батрахозаври і діадекти перенесені назад в амфібій. Автори виключають з основного складу черепах. Першим відгалуженням від загального стовбура вони вважають звіроподібних рептилій. [1] Solenodonosaurus поміщається між сеймуріаморфами і діадектоморфами. [1]
М. Ф. Івахненко (російський палеонтолог, доктор біологічних наук) повернувся до назви і первісного обсягу групи, і переробив систематику парарептилій на основі вивчення різноманітних їх представників з верхньої пермі і тріасу. Ним описані нові багаті фауни пізньопермських і ранньотріасових проколонофорів, показана належність до лантанозухів Chalkosaurus. Івахненко включив до складу парарептилій сеймуріаморф. Також парарептилії були об'єднані з іншими рептиліями в класі Reptiliomorfa. Івахненко встановлює новий ряд Nycteroleteromorfa, у якому об'єднані ніктеролети і лантанозухи. Ніктеролети стоять в основі діадектозаврів перед діадектоморфами. У 2001 році Івахненко розбив парарептилій на 3 підряди: Seymouriamorpha, Procolonomorpha і Cheloniamorpha. [1]
Так, згідно з М. Ф. Івахненко (2001), парарептилії — у класі рептиліоморф (Reptiliomorpha). У даній схемі до складу парарептилій входять сеймуріаморфи, проколофони (надряд Procolophonomorpha) і хелоніаморфи (надряд Cheloniamorpha). До останнього надряду входять ніктеролетери, токозаври, лантанозухи, рипеязаври, парейазаври і черепахи. У цілому в цій класифікації парарептилії виглядають природною групою, що виникла від амфібійних (анамній) предків-сеймуріаморф, незалежно досягла амніотного рівня і привела в підсумку до появи сучасних черепах. [1]
Оскільки ми використовуємо систематику, складену М. Ф. Івахненко, систематика підкласу Parareptilia виглядає наступним чином:
Надряд Procolophonomorpha
Ряд Procolohponida
Підряд Nyctiphruretia
Родина Nyctiphruretidae
Підряд Procolophpnia
Родина Spondylolestidae
Родина Procolophonidae
Надряд Cheloniamorpha
Ряд Pareiasaurida
Підряд Nycteroleterina
Надродина Nycteroleteroidaea
Родина Nycteroleteridae
Підродина Nycteroleterinae
Підродина Bashkyroleterinae
Родина Tokosauridae
Родина Rhipaeosauridae
Надродина Lanthanosuchoidea
Родина Lanthaniscidae
Родина Lanthanosuchidae
Надродина Elginioidea
Родина Elginiidae
Підряд Pareiasaurina [2]
Родина Bradysauridae
Родина Pareiasauridae
Час існування
Якщо враховувати тільки вимерлі форми парарептилій то час їх існування можна віднести до таких періодів як тріас (252,17 - 201,3 млн років тому) і перм (298,9 - 252,2 млн років тому) [1, 4]. Але якщо враховувати той факт що сучасні черепахи належать до цієї групи, то їх існування триває і донині.
Регіони найпоширеніших знахідок
Найбільша кількість знахідок знайдена в Європейській і Східній частині Росії (Оренбурзька область, Башкирія, Архангельська обл., Кіровська обл., Татарія), Північній Америці (Колорадо, Катлер Юра, Оклахома), Західній Європі (Німеччина, Шотландія, Англія), Південній і Східній Африці (Нігерія, Морадіс), також є деякі знахідки в Китаї (провінція Шаньсі), Бразилії (один рід і один вид), Мадагаскарі й Антарктиді.
Морфологічні особливості
Багато представників групи мозаїчно поєднують у своїй будові ознаки, властиві як амфібіям, так і ящерам. Однією з архаїчних особливостей багатьох парарептилій є відсутність у будові черепа скроневих вікон (порожнин позаду очниць, у яких у багатьох рептилій розташовуються потужні щелепні м'язи). [4]
Короткий параболічний череп із дахом ангуститабулярного типу (табличасті кістки стикаються з тім'яними) зі сплощеним закритим піднебінням. Радіальна структура даху черепа ямчасто-гребінчаста. Уздовж усіх швів даху черепа є гладкі смуги, оточені пористим валиком. Кістки щічного комплексу з'єднуються з тім'яним щитом на всьому протязі контакту зубчастим швом без різкого перегину між тім'яним щитом і щокою. На потиличному краї щоки по всій висоті розташовується глибока (вушна) вирізка. Добре виражені парафенестральні і базиптеригоїдні крила парасфеноїда (довга кістка, що лежить в основі черепа). У найпримітивніших форм не сформований комплекс атлант-епістофей. Найімовірніше був покрив з рогових лусок. [1]
Спосіб життя
В основній своїй масі парарептилії - порівняно невеликі (до 0,5 м завдовжки) хижаки, що живилися безхребетними (ніктеролетери, деякі проколофони); наприкінці пізньої пермі серед них з'являються великі рослиноїдні форми - парейазаври. [4]
Яскраві представники
Nyctiphruretus acudens знайдені скелети і черепи. Інші знахідки: р. Мезень в Архангельській області (Росія), Лешуконський район, Глядна Щ'єля, Козьмогородське, Усть-Няфта, Пеза-I, Мезенський район. Щелепні зуби з високими, трохи розширеними коронками, овальними в основі (рис. 1, 2). [2]

Рис. 1. Реконструкція
![Рис. 2. Череп Limnoscelis padulis – у черепі було невелике задньоскроневе вікно між верхньо-зовнішнім кінцем парокципетального відростка і tabulare або нижче і всередині від потиличного фланга squamosum (рис. 3, 4). [1]](https://s3.prozo.re/batrachos/images/nyctifruretus_acudens_ZqOyyHI.original.jpg)
Рис. 2. Череп
Limnoscelis padulis – у черепі було невелике задньоскроневе вікно між верхньо-зовнішнім кінцем парокципетального відростка і tabulare або нижче і всередині від потиличного фланга squamosum (рис. 3, 4). [1]
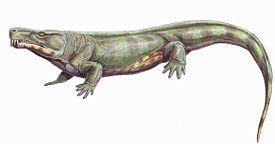
Рис. 3. Реконструкція
Tseajaia campi мав довжину черепа 12,5 см. Череп помірно високий. Є велике supraoccipetale; задньоскроневих вікон немає. Щелепне зчленування розташоване трохи нижче потиличного. Задній край даху черепа утворений парними табличастими і задньотім'яними кістками. Тім'яний отвір дуже великий, поздовжньо видовжений. Є довга вушна вирізка. Добре розвинена міжптеригоїдна ямка піднебіння. Поперечні фланги птеригоїдів добре розвинені, зубів не мають. Піднебінні зуби не розвинені. Щелепні зуби конічні, загострені, примаксилярно видовжені. Посередині максилярного зубного ряду 1 зуб має подобу ікла. Клубова кістка з довгим заднім відростком (Рис. 3; 4). [1]

Рис. 3. Комп'ютерна реконструкція
![Рис. 4. Скелет Tokosaurus perforatus - Дуже маленька форма, довжина черепа не більше 3 см. Окремі округлі остеродерми (вторинні шкірні окостеніння, що розташовуються в мезодермальному шарі шкіри) тільки в щічній області черепа [2]. Мав зароджуван](https://s3.prozo.re/batrachos/images/tseajaia588_ycWL7aD.original.jpg)
Рис. 4. Скелет
Tokosaurus perforatus - Дуже маленька форма, довжина черепа не більше 3 см. Окремі округлі остеродерми (вторинні шкірні окостеніння, що розташовуються в мезодермальному шарі шкіри) тільки в щічній області черепа [2]. Мав зароджуване скроневе вікно (Рис. 5; 6) [1]. Щелепні зуби тонкі, гостроконічні, передні зуби верхньощелепної кістки не збільшені [2].

Рис. 5. Реконструкція
![Рис. 6. Череп Lanthaniscus efremovi – знайдений в Архангельській області [1] і описаний за скелетом [2] (Рис. 7; 8).](https://s3.prozo.re/batrachos/images/macroleter_ZfM5GSY.original.jpg)
Рис. 6. Череп
Lanthaniscus efremovi – знайдений в Архангельській області [1] і описаний за скелетом [2] (Рис. 7; 8).

Рис. 7. Реконструкція
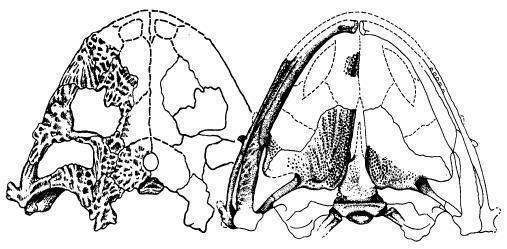
Рис. 8. Череп
Lanthaniscus watsoni має широкий і різко сплощений череп, довжиною близько 19 см (рис.). Дах черепа вкритий дрібнокомірчастою скульптурою, у щічній області ускладненою поздовжніми гребенями. Щічна область виступає назад від рівня потиличного і щелепного зчленувань, задній край щоки орнаментований виростами, розташованими головним чином на верхньоскроневих і квадратно-виличних кістках; місцями вирости є і на нижніх краях черепа – в області щелепного зчленування, під очницями і трохи позаду рівня ніздрів. Трохи позаду очниць є велике скроневе вікно, обмежене, як нижнє скроневе вікно діапсид, заочноямковою, виличною, квадратно-виличною і лускатою кістками. Очниця по нижньому краю переходить у велику вирізку, у якій могла розміщуватися залоза, яка могла мати функцію сольової, що виділяє надлишки катіонів. Заочноямкові кістки дуже великі і широко з'єднуються з надскроневими кістками. Вушна вирізка не розвинена, слухова кісточка не виявлена (Рис. 9; 10). [1]
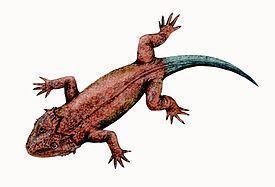
Рис. 9. Реконструкція
![Рис. 10. Череп Nanoparia luckhoffi мав довжину черепа 15 см (Рис. 11; 12). [1]](https://s3.prozo.re/batrachos/images/lanthaniscus_watsoni_E5Qynng.original.jpg)
Рис. 10. Череп
Nanoparia luckhoffi мав довжину черепа 15 см (Рис. 11; 12). [1]

Рис. 11. Реконструкція
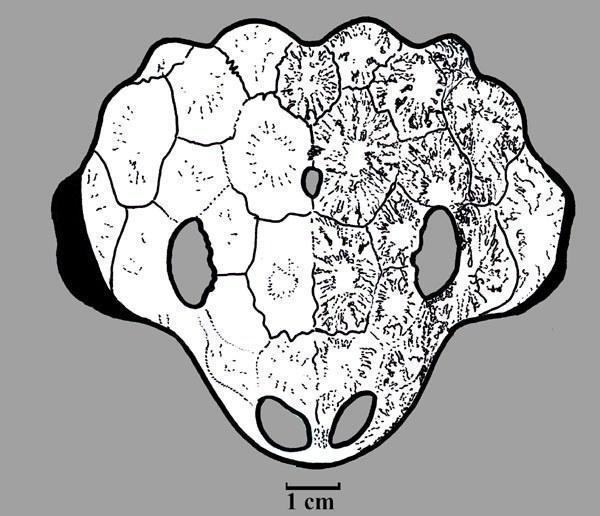
Рис. 12. Череп
Scutosaurus karpinskii мав дуже тонку підошву stapes. Сезамовидні кістки прилягали до заднього краю supratemporale або squamosum. Івахненко вказує, що ювенільні зуби мали плоску коронку з округлим верхнім краєм, що несе до 12 зубців. Зуби поступово набували кореня, що вростає в кістку. Спинний панцир був відсутній (Рис. 13; 14). [1]
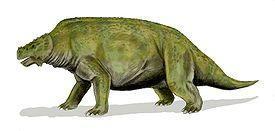
Рис. 13. Реконструкція
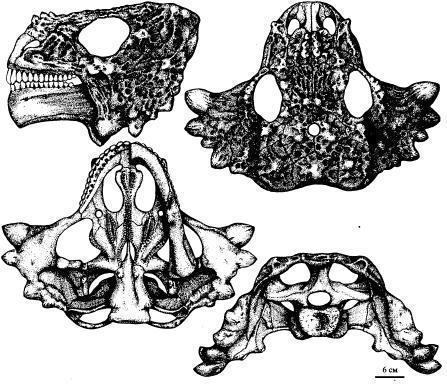
Рис. 14. Череп
Elginia mirabilis у довжину має близько 90 см. Менші серединні вирости черепа елгінії розташовані не на тім'яних кістках, а на розташованих позаду них додаткових елементах, що відповідають, скоріше за все, переднім панцирним пластинкам, що приросли до черепа. Піднебіння характеризується відносно слаборозвиненими птеригоїдними поперечними відростками, спрямованими, косо вперед і назовні. Зубів на птеригоїдних флангах в елгінії немає. Хоани позаду характерно вигнуті всередину. Зубний ряд укорочений, до 12 дев'ятизубцевих зубів у верхній щелепі. Спинні ребра не розширені, клубова кістка з добре розвиненим переднім відростком (Рис. 15; 16). [1]
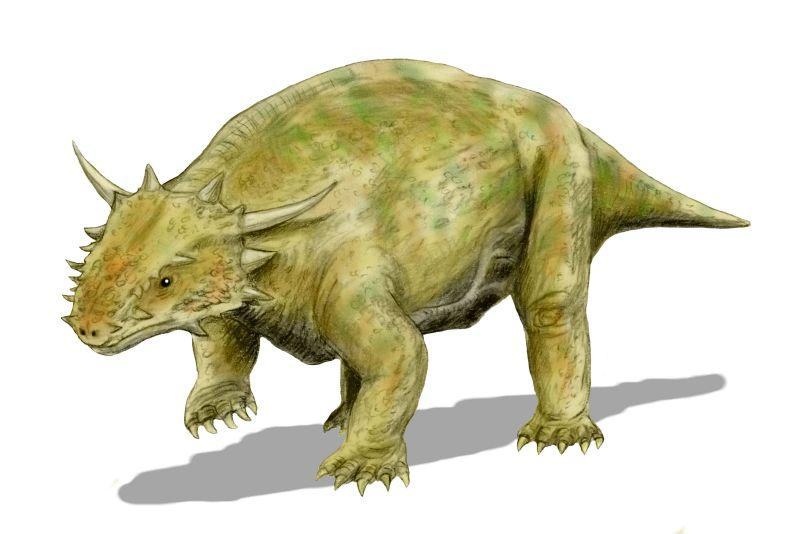
Рис. 15. Реконструкція
![Рис. 16. Череп. Owenetta kitchingorum задньотім'яні утворюють вузенький і короткий задній виступ (Рис. 17; 18). [1]](https://s3.prozo.re/batrachos/images/eliginia_mirabilis_2_NW7NVWG.original.png)
Рис. 16. Череп.
Owenetta kitchingorum задньотім'яні утворюють вузенький і короткий задній виступ (Рис. 17; 18). [1]

Рис. 17. Реконструкція
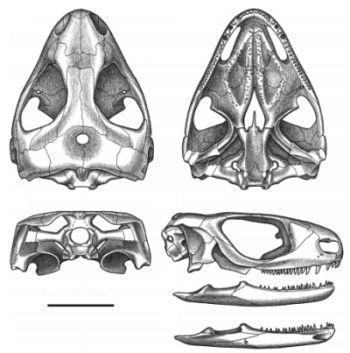
Рис. 18. Череп і нижня щелепа.
Список використаної літератури:
1. Л.П. Татарінов "Нариси з еволюції рептилій" - М. ГЕОС. - 2006 р. - 234 стор.
2. М.Ф. Івахненко, Є.М. Курочкін "Викопні хребетні Росії і суміжних країн" - М., ГЕОС - 2008 р. - 374 стор.
3. https://ru.wikipedia.org/wiki/Парарептилии
4. https://files.school-collection.edu.ru/dlrstore/4a71b57f-702d-448c-b206-0bd08b12b002/046_Parareptilii.html