
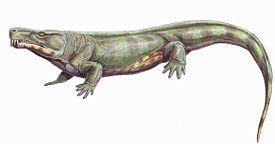
Subclass Parareptilia
Parareptilia is a large group of terrestrial tetrapods [4], a composite, possibly artificial group [3] that probably reached the reptilian level of organization independently of the rest of the vertebrates. Also, according to some views, turtles trace their origin to the parareptiles. [4] Systematics The term Parareptilia was proposed by E. Olson (an American palaeontologist) in 1947. Different classifications give a different composition for this group. [3] To begin with, it can be said that nowadays seymouriamorphs and diadectes are usually assigned to the amphibians, and most modern authors also bring seymouriamorphs closer to the pareiasaurs. [1] Carroll (an American scientist, doctor of science in the field of vertebrate palaeontology, specializing in Palaeozoic and Mesozoic amphibians and reptiles) is inclined to regard parareptiles as the sister group of amniotes, placing them above the Westlothiana. Watson (an English palaeontologist, author of many works on the evolution of fossil vertebrates) divided the most primitive reptiles without temporal fenestrae, or cotylosaurs (Cotylosauria), into three suborders: Seymouriamorpha, Diadectomorpha and Captorhinomorpha. Into the diadectomorphs he included diadectes and procolophons, which had an otic notch, and pareiasaurs, which had secondarily lost the otic notch. The diadectes were placed at the base of the diadectomorph cotylosaurs, which later made up the core of the parareptiles when they were distinguished. [1] According to E. Olson, the parareptiles initially united seymouriamorphs, diadectes, procolophons, pareiasaurs and turtles. The seymouriamorphs, raised to the rank of subclass, were joined with the diadectes; the two other groups of diadectomorph cotylosaurs — procolophons and pareiasaurs — were distinguished separately. Olson considered turtles closer to the diadectes than to the pareiasaurs. The parareptiles were contrasted with the eureptiles, which included all other reptiles. The seymouriamorphs, according to Olson, were considered ancestors of the diadectes. [1] Later Olson separated the seymouriamorphs into a special class, Batrachosauria, and regarded as parareptiles only the procolophons and pareiasaurs. Later still he abandoned the concept of parareptiles altogether, from which, successively, following the turtles, the seymouriamorphs and diadectes were excluded. [1] F. Huene (a German palaeontologist who studied fossil reptiles) united all the parareptiles with the anthracosaurs as a special branch of tetrapod evolution — Reptiliomorpha. Into this branch he also includes turtles, which he derives from the diadectomorphs. On the whole he derived all reptiles from the reptiliomorphs, but he separated the parareptiles into a branch of evolution distinct from the reptiles. [1] Romer (an American palaeontologist and morphologist, author of textbooks on palaeontology and vertebrate anatomy) regards seymouriamorphs together with diadectes within the amphibians, and unites procolophons and pareiasaurs in the suborder Procolophonia of the order of cotylosaurs. He assigns the millerosaurs there as well. [1] M. Heaton reunited the seymouriamorphs with the diadectes in a common order, Cotylosauria, and transferred it to the class of amphibians. He stressed the circumstance that the order was originally derived on the basis of the genus Diadectes=Empedocles. Procolophons, pareiasaurs and captorhinomorphs Heaton left among the reptiles, but excluded them from the cotylosaurs. The cotylosaurs, distinguished as the subclass Batrachosauria, he divides into seymouriamorphs and diadectomorphs. Heaton excludes from the seymouriamorphs the family Discosauriscidae, whose representatives led an amphibian way of life. Lanthanosuchus is not considered among the batrachosaurs. Among the batrachosaurs Heaton includes Nycteroleteridae. [1] Gauthier (a palaeontologist of Yale University), Kluge and Rowe (a professor of palaeontology from the University of Texas) distinguish parareptiles, with the batrachosaurs and diadectes transferred back to the amphibians. The authors exclude turtles from the core composition. As the first offshoot from the common stem they consider the mammal-like reptiles. [1] Solenodonosaurus is placed between the seymouriamorphs and the diadectomorphs. [1] M. F. Ivakhnenko (a Russian palaeontologist, doctor of biological sciences) returned to the name and original scope of the group and reworked the systematics of the parareptiles on the basis of studying their various representatives from the Upper Permian and the Triassic. He described new rich faunas of Late Permian and Early Triassic procolophons, and showed the belonging of Chalkosaurus to the lanthanosuchids. Ivakhnenko included the seymouriamorphs within the parareptiles. The parareptiles were also united with the other reptiles in the class Reptiliomorpha. Ivakhnenko establishes a new order, Nycteroleteromorpha, in which the nycteroleterids and lanthanosuchids are united. The nycteroleterids stand at the base of the diadectosaurs, before the diadectomorphs. In 2001 Ivakhnenko split the parareptiles into 3 suborders: Seymouriamorpha, Procolophonomorpha and Cheloniamorpha. [1] Thus, according to M. F. Ivakhnenko (2001), the parareptiles are within the class Reptiliomorpha. In this scheme the parareptiles include seymouriamorphs, procolophons (superorder Procolophonomorpha) and cheloniamorphs (superorder Cheloniamorpha). The latter superorder includes nycteroleterids, tokosaurs, lanthanosuchids, rhipaeosaurs, pareiasaurs and turtles. On the whole, in this classification the parareptiles look like a natural group that arose from amphibian (anamniote) seymouriamorph ancestors, independently reached the amniote level, and ultimately led to the appearance of modern turtles. [1] Since we use the systematics compiled by M. F. Ivakhnenko, the systematics of the subclass Parareptilia look as follows: Superorder Procolophonomorpha Order Procolophonida Suborder Nyctiphruretia Family Nyctiphruretidae Suborder Procolophonia Family Spondylolestidae Family Procolophonidae Superorder Cheloniamorpha Order Pareiasaurida Suborder Nycteroleterina Superfamily Nycteroleteroidea Family Nycteroleteridae Subfamily Nycteroleterinae Subfamily Bashkyroleterinae Family Tokosauridae Family Rhipaeosauridae Superfamily Lanthanosuchoidea Family Lanthaniscidae Family Lanthanosuchidae Superfamily Elginioidea Family Elginiidae Suborder Pareiasaurina [2] Family Bradysauridae Family Pareiasauridae Time of existence If one considers only the extinct forms of parareptiles, the time of their existence can be assigned to such periods as the Triassic (252.17 – 201.3 million years ago) and the Permian (298.9 – 252.2 million years ago) [1, 4]. But if one takes into account the fact that modern turtles belong to this group, then their existence continues to this day. Regions of the most common finds The largest number of finds has been found in the European and Eastern parts of Russia (Orenburg region, Bashkiria, Arkhangelsk region, Kirov region, Tatarstan), North America (Colorado, Cutler Formation, Oklahoma), Western Europe (Germany, Scotland, England), Southern and Eastern Africa (Nigeria, Moradi), and there are also some finds in China (Shanxi province), Brazil (one genus and one species), Madagascar and Antarctica. Morphological features Many representatives of the group combine in a mosaic fashion features characteristic of both amphibians and reptiles. One of the archaic features of many parareptiles is the absence in the skull structure of temporal fenestrae (cavities behind the orbits in which, in many reptiles, powerful jaw muscles are located). [4] A short parabolic skull with a roof of the angustitabular type (the tabular bones contact the parietals) with a flattened, closed palate. The radial structure of the skull roof is pit-and-ridge. Along all the sutures of the skull roof there are smooth strips surrounded by a porous ridge. The bones of the cheek complex join the parietal shield along the entire length of contact by a serrate suture, without a sharp bend between the parietal shield and the cheek. On the occipital edge of the cheek, along its entire height, a deep (otic) notch is located. The parafenestral and basipterygoid wings of the parasphenoid (a long bone lying at the base of the skull) are well expressed. In the most primitive forms the atlas-axis complex is not formed. Most likely there was a covering of horny scales. [1] Way of life In their main mass the parareptiles are comparatively small (up to 0.5 m long) predators that fed on invertebrates (nycteroleterids, some procolophons); at the end of the Late Permian large herbivorous forms appear among them — the pareiasaurs. [4] Notable representatives Nyctiphruretus acudens — skeletons and skulls have been found. Other finds: the Mezen River in the Arkhangelsk region (Russia), Leshukonsky district, Glyadnaya Shchelya, Kozmogorodskoye, Ust-Nyafta, Peza-I, Mezensky district. The jaw teeth have high, slightly widened crowns, oval at the base (Figs. 1, 2). [2] Fig. 1. Reconstruction Fig. 2. Skull Limnoscelis padulis — in the skull there was a small post-temporal fenestra between the upper-outer end of the paroccipital process and the tabulare, or below and inward from the occipital flange of the squamosum (Figs. 3, 4). [1] Fig. 3. Reconstruction Tseajaia campi had a skull length of 12.5 cm. The skull is moderately high. There is a large supraoccipitale; there are no post-temporal fenestrae. The jaw articulation is located slightly below the occipital one. The posterior edge of the skull roof is formed by the paired tabular and postparietal bones. The parietal foramen is very large, longitudinally elongated. There is a long otic notch. The interpterygoid vacuity of the palate is well developed. The transverse flanges of the pterygoids are well developed and bear no teeth. Palatal teeth are not developed. The jaw teeth are conical, pointed, premaxillarily elongated. In the middle of the maxillary tooth row 1 tooth resembles a canine. The ilium has a long posterior process (Figs. 3; 4). [1] Fig. 3. Computer reconstruction Fig. 4. Skeleton Tokosaurus perforatus — a very small form, the skull length no more than 3 cm. Separate rounded osteoderms (secondary skin ossifications located in the mesodermal layer of the skin) only in the cheek region of the skull [2]. It had an incipient temporal fenestra (Figs. 5; 6) [1]. The jaw teeth are thin, sharply conical; the anterior teeth of the maxilla are not enlarged [2]. Fig. 5. Reconstruction Fig. 6. Skull Lanthaniscus efremovi — found in the Arkhangelsk region [1] and described from a skeleton [2] (Figs. 7; 8). Fig. 7. Reconstruction Fig. 8. Skull Lanthaniscus watsoni has a broad and sharply flattened skull, about 19 cm long (fig.). The skull roof is covered with a fine-celled sculpture, complicated in the cheek region by longitudinal ridges. The cheek region projects backward beyond the level of the occipital and jaw articulations; the posterior edge of the cheek is ornamented with outgrowths located mainly on the supratemporal and quadratojugal bones; in places the outgrowths are also on the lower edges of the skull — in the area of the jaw articulation, beneath the orbits, and slightly behind the level of the nostrils. Slightly behind the orbits there is a large temporal fenestra, bounded, like the lower temporal fenestra of diapsids, by the postorbital, jugal, quadratojugal and squamosal bones.

The orbit, along its lower edge, passes into a large notch in which a gland could be located that may have had the function of a salt gland, excreting excess cations.
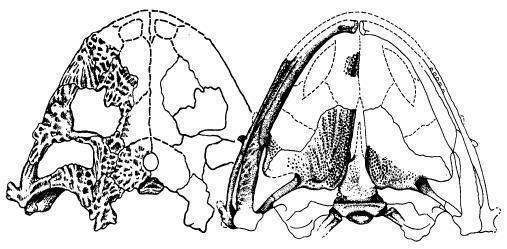
![Рис. 2. Череп Limnoscelis padulis – у черепі було невелике задньоскроневе вікно між верхньо-зовнішнім кінцем парокципетального відростка і tabulare або нижче і всередині від потиличного фланга squamosum (рис. 3, 4). [1]](https://s3.prozo.re/batrachos/images/nyctifruretus_acudens_ZqOyyHI.original.jpg)
The postorbital bones are very large and join broadly with the supratemporal bones. The otic notch is not developed; the auditory ossicle has not been found (Figs.
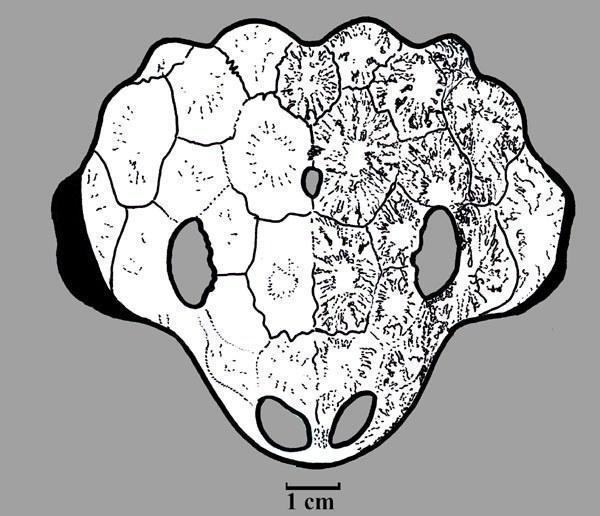
9; 10). [1] Fig. 9. Reconstruction Fig. 10. Skull Nanoparia luckhoffi had a skull length of 15 cm (Figs. 11; 12). [1] Fig.

11.
![Рис. 4. Скелет Tokosaurus perforatus - Дуже маленька форма, довжина черепа не більше 3 см. Окремі округлі остеродерми (вторинні шкірні окостеніння, що розташовуються в мезодермальному шарі шкіри) тільки в щічній області черепа [2]. Мав зароджуван](https://s3.prozo.re/batrachos/images/tseajaia588_ycWL7aD.original.jpg)
Reconstruction Fig. 12. Skull Scutosaurus karpinskii had a very thin footplate of the stapes.

The sesamoid bones adjoined the posterior edge of the supratemporale or squamosum.
![Рис. 6. Череп Lanthaniscus efremovi – знайдений в Архангельській області [1] і описаний за скелетом [2] (Рис. 7; 8).](https://s3.prozo.re/batrachos/images/macroleter_ZfM5GSY.original.jpg)
Ivakhnenko indicates that the juvenile teeth had a flat crown with a rounded upper edge bearing up to 12 cusps.

The teeth gradually acquired a root growing into the bone.
A dorsal carapace was absent (Figs. 13; 14). [1] Fig. 13. Reconstruction Fig. 14. Skull Elginia mirabilis is about 90 cm in length. The smaller median outgrowths of Elginia's skull are located not on the parietal bones, but on additional elements positioned behind them, corresponding most likely to anterior armour plates fused to the skull. The palate is characterized by relatively weakly developed pterygoid transverse processes directed obliquely forward and outward. There are no teeth on Elginia's pterygoid flanges. The choanae are characteristically curved inward at the back.
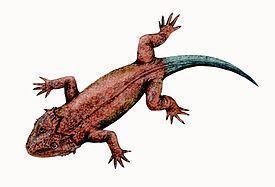
The tooth row is shortened, up to 12 nine-cusped teeth in the upper jaw.
![Рис. 10. Череп Nanoparia luckhoffi мав довжину черепа 15 см (Рис. 11; 12). [1]](https://s3.prozo.re/batrachos/images/lanthaniscus_watsoni_E5Qynng.original.jpg)
The dorsal ribs are not widened; the ilium has a well-developed anterior process (Figs.

15; 16).
[1] Fig. 15.
Reconstruction Fig.
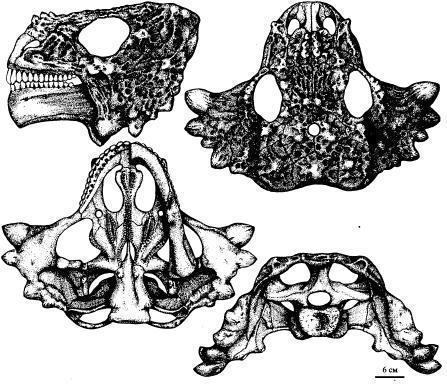
16. Skull. Owenetta kitchingorum — the postparietals form a narrow and short posterior projection (Figs. 17; 18). [1] Fig. 17. Reconstruction Fig.
18.
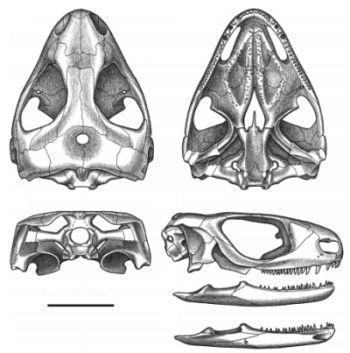
![Рис. 16. Череп. Owenetta kitchingorum задньотім'яні утворюють вузенький і короткий задній виступ (Рис. 17; 18). [1]](https://s3.prozo.re/batrachos/images/eliginia_mirabilis_2_NW7NVWG.original.png)
Skull and lower jaw.

List of references used: 1.
L. P. Tatarinov, "Essays on the evolution of reptiles" — M., GEOS — 2006 — 234 pp. 2. M. F. Ivakhnenko, E. N. Kurochkin, "Fossil vertebrates of Russia and adjacent countries" — M., GEOS — 2008 — 374 pp. 3. https://ru.wikipedia.org/wiki/Парарептилии 4. https://files.school-collection.edu.ru/dlrstore/4a71b57f-702d-448c-b206-0bd08b12b002/046_Parareptilii.html