
Шишкин, 2012. Системная обусловленность формообразования и ее проявления в палеонтологической летописи
Шишкин М. А. Системная обусловленность формообразования и ее проявления в палеонтологической летописи // Палеонтологический журнал, 2012, № 4, с. 3–15.
СИСТЕМНА ЗУМОВЛЕНІСТЬ ФОРМОУТВОРЕННЯ І ЇЇ ПРОЯВИ В ПАЛЕОНТОЛОГІЧНОМУ ЛІТОПИСІ
© 2012 р. М. А. Шишкін
Загальною основою механізмів формоутворення є забезпечення спрямованості індивідуального розвитку як цілого до дорослої норми. Стійкість цього цілепокладання є головною властивістю живої організації, що потребує пояснення. Кожен крок еволюції є вимушеною реорганізацією онтогенезу як системи в напрямку нової кінцевої мети (рівноважного стану), тобто нової норми. Зростання стійкості її здійснення в послідовних життєвих циклах під дією добору веде до перебудови розвитку даного еволюційного новшества, що поширюється в напрямку ранніх стадій. У підсумку відмінність між записом розвитку новшества і вихідним анцестральним онтогенезом усе більше перетворюється з пролонгуючого (рекапітулюючого) на дивергентне, що відповідає закону Бера. Зміни, пов'язані з цим стабілізаційним процесом, фіксуються в палеонтологічному літописі у вигляді низки характерних проявів — вихідної нестійкості новопридбань, перебудови їхнього морфогенезу з плином часу і переважного збереження рекапітуляцій анцестральних рис у найближчих нащадків.
I. Цілісність розвитку як контролюючий фактор морфогенезу
Пошук пояснення будь-якого феномену можливий на різних рівнях розгляду, що відповідають його розчленуванню на все більш часткові компоненти. Вибір кожного з них може бути виправданий для певних дослідницьких завдань. Але змістовні закони, що описують сутність явища як цілого, належать до найбільш верхніх із цих рівнів. Лише в цьому світлі може бути оцінена природа причинних зв'язків, що розкриваються при більш часткових редукціоністських підходах. Суть наукового пізнання якраз і полягає в тому, що для явищ, які здавалися незалежними, виявляється взаємозв'язок і співпідпорядкування загальним законам.
Усвідомлення такого порядку речей неминуче для будь-якої галузі досліджень, і теоретична біологія є тому яскравим прикладом. Так, спроби звести ембріональний розвиток до мозаїки шляхів клітинної детермінації, характерні для другої половини XIX століття, змінилися уявленням про єдине морфогенетичне поле, що контролює розвиток на всіх його етапах. У наш час подібну ж перебудову зазнає панівне еволюційне мислення, що ще сорок років тому вірило у всесилля редукціоністських принципів неодарвінізму. На зміну цьому приходить усвідомлення, що “макроеволюція не може бути зведена до теорії, де вихідними поняттями є властивості індивідуальних генів”, і що належна теорія повинна “враховувати вимоги верхніх рівнів розгляду” (Maderson et al., 1982).
Сказане визначає належний підхід до розуміння дії і становлення формоутворювальних механізмів індивідуального розвитку — чи йдеться про процеси, що спостерігаються в сучасних організмів, чи про їхні сліди в палеонтологічному літописі. Усе різноманіття формативних актів, що здійснюються в ході нормального розвитку, так чи інакше гармонізоване в його рамках, тобто підпорядковане закономірностям, що керують онтогенезом як цілим. Серед останніх найочевиднішою характеристикою є спрямованість усього процесу до заданого кінцевого підсумку, тобто присутність у ньому цілепокладального (телеономічного) контролю, як це було вперше підкреслено К. Бером. Ця інтегральна властивість розвитку і повинна служити відправною точкою для аналізу всіх його часткових процесів.
Цілепокладальна поведінка є властивістю системних об'єктів, що виражає їхню здатність до саморегуляції. Немає сумніву, що індивідуальний розвиток є цілісною динамічною системою, що послідовно коригує свій стан через зворотні зв'язки в бік кінцевої рівноваги, тобто дорослої норми (Waddington, 1957; Свєтлов, 1972; Шишкін, 2006).
Усі прояви цілісності розвитку, що виявляються ембріологією, зрештою є вираженням його цілеспрямованості. Передусім, це зменшення розмаху мінливості в ході розвитку, встановлене ще Бером. По-друге, це явище еквіфінальності або стійкості підсумку процесу щодо способів його здійснення (Roux, 1896; Driesch, 1908). Дана властивість має безліч проявів, що стосуються як дорослих, так і “установочних” провізорних стадій розвитку, наприклад, способу гаструляції в багатьох первинноротих. Ці властивості можуть виявлятися як експериментально (різні шляхи регенерації особини, розвиток повноцінних зародків із розділених бластомерів або ж самозбирання останніх в одне ціле), так і в нормальному розвитку. До останнього випадку передусім належать альтернативні шляхи розмноження (статеве і вегетативне) або типи розвитку (прямий або з метаморфозом) в одних і тих самих або близьких видів, причому вихідні стадії онтогенетичного циклу можуть бути гранично різні.
У ширшому сенсі феномен еквіфінальності проявляється на рівні історичних змін, коли одна й та сама властивість організації може здійснюватися в розвитку у предка і нащадків на різній формативній основі (див. нижче).
Якщо говорити про часткові процеси, що складають розвиток, то їхній цілепокладальний характер найвиразніший у морфогенезах регуляційного типу, проявляючись, наприклад, у таких феноменах як множинне забезпечення формоутворювальних актів або здатність компетентного зачатка до однієї й тієї самої реакції у відповідь на широкий спектр експериментальних індукційних впливів.
Головне узагальнення ембріології, що стосується спрямованості онтогенезу, водночас виражає притаманне їй розуміння механізму цього процесу. Це уявлення про цілісну детермінацію розвитку, присутню вже в зиготі і таку, що визначає формування всіх рис організації за принципом від загального до часткового (Baer, 1828; Гертвіг, 1895; Свєтлов, 1964). Весь хід подій, за висловом П.Г. Свєтлова, розглядається тут як “вмальовування” послідовних актів диференціації в рамки контролюючого їх загального плану розвитку. Розвиток розуміється як епігенез, тобто наростання різноманіття складових його факторів у рамках їхньої інтегральної зумовленості. Цей принцип передбачає відсутність прямих причинних залежностей між властивостями організму і частковими структурами зародкової клітини.
Як відомо, пошуки таких залежностей складали суть ранніх гіпотез про механізм розвитку, заснованих на “целюлярному” преформізмі, тобто зведенні всього онтогенезу до мозаїчної сукупності типів клітинної диференціації. Передбачалося, що вони контролюються окремо тими чи іншими клітинними частинками — цитоплазматичними, або ядерними (теорія Рой Ланкастера або мозаїчна теорія Ру-Вейсмана, див. Свєтлов, 1978). Цей редукціоністський підхід, виявившись несумісним із фактами ембріології, надалі здобув новий ґрунт у хромосомній генетиці — у період, коли її головне завдання складало пряме проєктування генів на ознаки дорослого організму за допомогою гібридного аналізу.
За мірою залучення генетики до аналізу процесів розвитку ці погляди трансформувалися (у теоретичному плані) у бік їхньої більшої адаптації до реальної картини онтогенетичних подій1. Нині вони знайшли вираження в уявленнях про генні мережі, де дія регуляторних генів вищого порядку запускає послідовні етапи каскадної експресії генів-мішеней (напр., Гілберт та ін., 1997). Але за всієї ускладненості таких схем порівняно з прямолінійним преформізмом минулого, вони залишаються в рамках того, що Р. Гольдшмідт (Goldschmidt, 1940) називав принципом “інтегрованої генної дії”, тобто уявленням, за яким доросла організація виникає із взаємодії незалежних факторів. Те, що роль пускового механізму відводиться в цих схемах лише невеликому числу ініціюючих генів, не виводить їх за рамки ідеї мозаїчної зумовленості розвитку. Остання залишається антагоністом ідеї цілісного управління онтогенетичним процесом. Справді, сам постулат про незалежність факторів, що координовано запускають розвиток, за визначенням виключає відповідь на питання — що ж їх координує?
Однак у реальному житті науки спосіб осмислення фактів дослідником може суттєво відхилятися від прийнятих ним теоретичних постулатів (пор. Шишкін, 2010). З цієї причини в сьогоднішньому багажі генетики можна знайти чимало узагальнень, що виражають альтернативну для неї ідею цілісної детермінації. Уже Т. Морган (1924) припускав, що дискретність спадкових факторів не обов'язково суперечить зумовленості кожної властивості організму всією зародковою плазмою. Йому ж належить думка, що диференціальна активність генів у різних типах клітин пов'язана з потраплянням еквіпотенціальних ядер у різну цитоплазму — судження, що поділяється нині більшістю генетиків (Корочкін, 2002). По суті, це визнання, що поведінка геному клітини контролюється її цілісними властивостями — названі вони цитоплазматичним оточенням, клітинним середовищем чи якось інакше.
Підтвердженням цього висновку служать багато експериментальних узагальнень, що стосуються впливу клітини на будову і функцію свого генетичного апарату. Показано, що всі щаблі матричного синтезу потенційно неоднозначні, включаючи як різні можливості редагування транскриптів, так і їхньої подальшої трансляції і посттрансляційних змін білкових молекул — залежно від фізіологічних умов у клітині (Інге-Вечтомов, 1976; Голубовський, 2001). У підсумку результат синтезу завжди визначається характером його внутрішньоклітинної регуляції в ході всього процесу.
Далі, багатьма дослідами, особливо на одноклітинних організмах, було встановлено, що клітина здатна локально перебудовувати свій геном стосовно завдань її функціонування в конкретній ситуації (Cairns et al., 1988; Hall, 1990). Ці явища відображають еволюційно вироблену здатність клітини до адаптивних модифікацій на генетичному рівні (хоча для генетиків, що її відкрили, вона зазвичай постає вражаючим свідченням “адаптивних мутацій”, що спрямовано здійснюються організмом). Таким чином, клітина “здатна відповісти на виклик середовища активним генетичним пошуком”, тобто “діяти цілеспрямовано” відповідно до наявних умов (Голубовський, 2001).
Усе це показує, що генетична активність клітини в ході її життя керується в принципі таким же системним регуляторним механізмом (що задає доцільний хід подій), як і онтогенез у цілому. Це здається нам ще одним свідченням на користь того, що вся сукупність узагальнень генетики буде з часом неминуче переосмислена у світлі законів, що керують індивідуальним розвитком (пор. Шишкін, 2006, 2010).
Стосовно наших завдань ми маємо право зробити висновок, що в обох зіставлюваних випадках споконвічно присутній контролюючий фактор цілісності. На рівні клітин його специфіка, очевидно, відповідає їхнім конкретним генераціям, а в цілому організмі він постає як послідовність станів, що характеризують спадкоємні фази онтогенезу. Останнє уявлення відповідає ідеї А. Гурвіча (Gurwitsch, 1922) про перетворюване біологічне поле, кожен зі станів якого є інваріантою, що визначає хід розвитку на найближчий його відрізок. У підсумку зміна характеристик поля в розвитку є функцією від шляху, пройденого його субстратом.
Ідея поля видається для ембріології “єдиною концепцією цілісного причинного вивчення ходу онтогенезу”, що дозволяє пояснити його спрямованість (Свєтлов, 1964). Очевидно, саме тут слід шукати механізм онтогенетичного цілепокладання, що визначає хід і координацію окремих формоутворювальних процесів у нормальному розвитку.
II. Еволюція як зміна цілепокладання в системі онтогенетичного здійснення
“Доцільність не є споконвічною властивістю живого” (Шмальгаузен, 1982) і повинна бути предметом еволюційного пояснення. Якщо еволюція є трансформацією онтогенезу як цілого, то кожен її крок означає перебудову системи розвитку в бік нової кінцевої рівноваги (“мети”, або адаптивної норми) замість утраченого. Система тим самим відновлює тимчасово порушену стійкість. Згідно з епігенетичною теорією еволюції, що постулює такий хід подій, це відбувається на основі “наявних можливостей” вихідної норми реакції, тобто шляхом збереження добором однієї з термінальних лабільних аберацій, здійсненних у рамках цієї норми (Шмальгаузен, 1982; Шишкін, 1988а, б, 2006, 2010). Ця аберація і є моделлю нового найближчого стану рівноваги (рис. 1, а). Вона визначає напрямок корекції властивостей системи розвитку в ході добору.
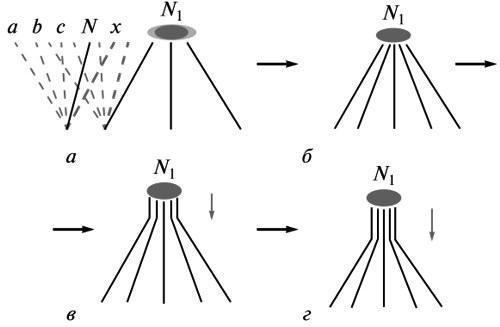
Рис. 1. Еволюційна зміна як стабілізація нової “мети” розвитку: а — простір можливих шляхів розвитку в рамках вихідної норми N (зліва) і в рамках початкової зміни в бік нової норми N1 (у центрі); формування нової норми на основі однотипних індивідуальних варіацій, що зберігаються добором (справа); б, в, г — послідовні стадії стабілізації розвитку нової норми в ході поширення процесу (у поколіннях) від підсумкової стадії до все більш ранніх. Позначення: а, Ь, с — потенційні термінальні аберації вихідної дорослої норми; N — вихідна норма; N1 — нова норма на різних етапах її становлення; х — одна з аберацій вихідної норми, що зберігається добором у нових умовах. Горизонтальні стрілки — етапи стабілізації зміни; вертикальні стрілки — напрямок історичних стабілізаційних перетворень.
Таким чином, шлях становлення кожного нового типу цілепокладання (зміненої норми) є історією зростання точності “наведення” розвитку на нову обрану мету (рис. 1, а-г). І навпаки, відсуваючись до ранніх етапів цього процесу, ми повинні спостерігати все більший розкид індивідуальних варіантів розвитку навколо цієї майбутньої мети. Початковому етапу повинна відповідати максимальна лабільність аберації, що зберігається добором. Це виражається в граничній неоднозначності відтворення її фенотипу в безпосереднього потомства.
Звідси зрозумілий вектор змін в еволюції морфогенезу. Кожен крок у стабілізації нової мети — це переважне збереження особин, що успішно повторили фенотип своїх раніше відібраних батьків. Тим самим отримують перевагу ті індивідуальні “еквіфінальні” варіанти потомства, у яких безпосередня формоутворювальна основа збережуваного фенотипу виявилася найменш чутливою до коливань факторів розвитку (процес, описаний Шмальгаузеном як перехід від залежного здійснення новшества до все більш автономного). Серед нащадків цих варіантів знову отримають селективну перевагу ті, у яких завадостійкість новшества виявилася найбільшою — за рахунок залучення в його інтегрований морфогенез усе більш ранніх онтогенетичних процесів (рис. 1, в, г).
Таким чином, у міру стабілізації нової адаптивної норми, перебудова її морфогенезу поширюється в поколіннях у бік попередніх стадій розвитку. Разом із поглибленням цього процесу, у тому ж напрямку неминуче поширюється і його морфологічне вираження (Шишкін, 1988а, б, 2006, 2010).
У сумі, історія становлення будь-якого новшества — це ланцюг послідовних випробувань добором його носіїв на здатність відтворювати самих себе незалежно від збурень ходу розвитку. Процес добору виступає тут як послідовність затухаючих циклів корекції, що ведуть до стабілізації нової норми.
Те, що перетворення морфогенезу тривало існуючих структур протікають описаним вище шляхом, захоплюючи все більш ранні стадії розвитку, добре відомо з досвіду порівняльної ембріології і палеонтології (хоча тут зазвичай доступні для оцінки не елементарні еволюційні події, а їхній сумарний підсумок, об'єднаний загальним вектором змін).
Найочевидніша закономірність, що виявляється в цих випадках, полягає в тому, що властивості організації, які виникли в умовах дорослого життя предка і, безсумнівно, морфогенетично залежали від них, у нащадків проявляються значно раніше і більш автономно — задовго до настання умов, що відповідають функціонуванню цих властивостей. Наприклад, черепашка і нога в молюсків, що виникли як адаптації дорослої тварини до проживання на дні, у сучасних форм зазвичай з'являються вже на стадії пелагічної личинки, поза зв'язком з умовами їхнього еволюційного виникнення. Так само в сучасних птахів особливості скелета крила, що склалися в ході пристосування до польоту у предків, проявляються вже ембріонально, поза зв'язком з польотом.
Стабілізаційні перетворення такого роду, що виражають поглиблену перебудову морфогенезу дорослої структури при збереженні підсумкового плану її будови, описані в літературі під безліччю назв — таких як акселерація, конденсація, адюльтація, ембріоналізація2 тощо (Шишкін, 2006) .
Більш непрямим, але настільки ж наочним свідченням такого шляху еволюції морфогенезу є численні паралелізми між варіаціями в одних таксонів і діагностичними ознаками в їхніх найближчих родичів. Очевидно, другий стан може бути витлумачений як результат історичної стабілізації першого (пор. нижче).
Зазначена спрямованість ходу подій у стабілізаційних процесах проливає світло на справжній сенс закону Бера (наростання онтогенетичної дивергенції між спорідненими формами по ходу розвитку). Безліч авторів вважають його “фундаментально несумісним” із біогенетичним законом Е. Геккеля, гадаючи, що геккелівські рекапітуляції дорослих анцестральних станів є насправді лише проявами зародкової схожості предка і нащадка, що описуються правилом Бера (de Beer, 1971; Gould, 1977).
Насправді змістовними є обидва ці узагальнення, характеризуючи в гранично ідеалізованому вигляді дві сторони еволюційного процесу (Шишкін, 1988б). Закон Бера не викликає питань лише щодо типів розвитку, що дивергували від спільного предка; але при порівнянні онтогенезів, що належать до однієї філетичної лінії, можливість їхнього розходження (а не пролонгації одного іншим) сама по собі не очевидна і потребує пояснення.
Еволюція починається з термінальних змін організації. Перші кроки цього селективного процесу, що закріплює одну з пізніх аберацій наявної норми, спершу створюють тим самим лише нову модифікацію останньої, реалізовану тільки в даних змінених умовах (Шмальгаузен, 1982; пор. вище). Тобто, у згоді з геккелівським принципом, колишня норма тут значною мірою зберігається як субтермінальна стадія розвитку, що служить основою для нової адаптивної реакції (рис. 2, С–D(DI)). Але, у міру перетворення цієї реакції на безумовну стійку норму, наростає її автономізація від факторів середовища, якими спочатку індукувався її морфогенез. Тим самим вихідна для нової норми анцестральна стадія, що більш чи менш повно відтворювалася на попередньому етапі, повинна бути перетворена, щоб забезпечити формативний механізм для цієї зміни (рис. 2, D(DII)). У міру подальшої стабілізації нової норми в низхідних нащадків ця перебудова морфогенезу буде посилюватися, одночасно поширюючись на все більш ранні стадії (див. вище). У результаті рекапітуляція анцестрального стану буде стиратися все більше, а дивергенція між вихідним і похідними варіантами онтогенезу — поглиблюватися (рис. 2, С–D (DIV)).
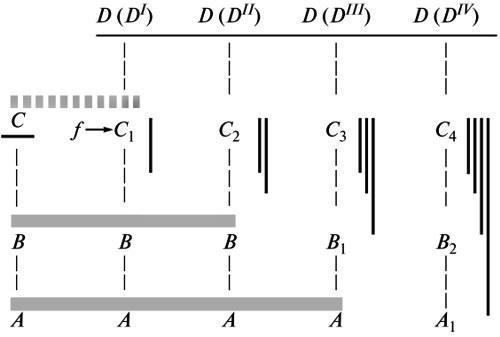
Рис. 2. Поглиблення дивергенції між розвитком у предка і низхідних нащадків у процесі перебудови морфогенезу еволюційного новшества. Позначення: А, А1; В, В1, В2; С, С1-4 — стадії розвитку; D(DI)-D(DIV) — історичні етапи стабілізації новшества; f— зовнішній фактор, що спочатку впливав на формоутворення. Сірі горизонтальні лінії — послідовні рівні дивергенції; темні вертикальні лінії — накопичення стабілізаційних змін (за Шишкіним, 1988б, зі змінами).
Таким чином, еволюція нових придбань є історією безперервного перетворення їхнього онтогенетичного запису з подовжуючого (за Геккелем) на дивергентний (за Бером) порівняно з онтогенезом вихідної предкової форми (Шишкін, 1988б).
Зі сказаного зрозуміло, що чим менша філетична дистанція між предком і нащадком, з тим більшою ймовірністю в пізньому онтогенезі нащадка можуть зберегтися сліди анцестрального дорослого стану, і навпаки. Ця закономірність, добре знайома з досвіду вивчення викопних решток (див., наприклад, Руженцев, 1960), пояснює, чому основна критика біогенетичного закону виходить від фахівців із рецентних організмів, тоді як палеонтологи ставляться до нього набагато позитивніше. Справді, при аналізі, що включає сучасні форми, об'єктами порівняння є організми, віддалені один від одного величезними інтервалами еволюційних перетворень, не кажучи вже про переважно нелінійний характер їхніх споріднених відносин. Навпаки, палеонтолог нерідко має справу з більш чи менш спадкоємними ланками в рамках одного філетичного стовбура, де повторення предкових рис у пізньому онтогенезі нащадка в багатьох випадках виступає як очевидний факт.
III. Свідчення еволюції формоутворення, що реєструються на викопних рештках
Зі сказаного вище вимальовується коло закономірностей еволюції морфогенезу, прояви
яких можна з великою ймовірністю очікувати при зіставленні викопних організмів. До їхнього числа передусім входять наступні.
(1) Виникнення новшеств у вигляді нестійких варіацій — не лише з точки зору частоти і ступеня їхнього прояву, але й співвідношень з іншими рисами організації.
(2) Історична стабілізація цих придбань, що виражається (крім власне зростання їхньої стійкості) у перебудові способу їхнього розвитку і в усе більш ранньому видимому прояві в онтогенезі3.
(3) Висока ймовірність збереження дорослих рис предка в пізньому онтогенезі порівняно близьких нащадків, і тим самим — можливість виявлення в них давніх рекапітуляцій, стертих у пізніших типах розвитку.
Як уже сказано, в останньому з цих випадків викопний матеріал за своєю інформативністю має беззаперечну перевагу перед рецентним; але щодо закономірностей (1) і (2) ця відмінність не настільки очевидна. Палеонтологічний літопис зберігає в основному лише усталені морфотипи, тобто ті, що змогли тривало персистувати в часі й просторі. Водночас ранні етапи їхнього становлення, що характеризувалися швидкою трансформацією і протікали в локальних популяціях на обмежених територіях, залишають мало прямих свідчень. Ці етапи вловлюються, головним чином, опосередковано — у вигляді згасаючих слідів анцестральних варіацій, що збереглися в розвитку більш стабілізованих нащадків (особливо в педоморфних лініях).
Такі ж застереження необхідні й щодо стабілізаційної перебудови розвитку нових придбань. Свідчення цього процесу, що вловлюються на викопному матеріалі, зводяться в основному до його морфологічних проявів і поширення останніх на більш ранні стадії. Власне ж зміни факторів морфогенезу (доступні для порівняльного вивчення в рецентних форм) можуть простежуватися лише в рідкісних випадках. Крім того, як уже сказано вище, матеріалом для оцінки стабілізаційних процесів на викопному матеріалі служать зазвичай не елементарні еволюційні зрушення, а їхні сумарні підсумки в рамках односпрямованих послідовностей філетичних змін.
Джерела даних про хід розвитку, доступні на викопному матеріалі, можуть бути узагальнені у вигляді кількох головних категорій (Макрідін, Шишкін, 1988). Це, по-перше, вікові серії або їхні окремі ключові стадії (наприклад, ларвальні морфотипи в комах або ембріони в амніот). Грубими моделями вікових серій, що дають у ряді випадків інформацію про якісні перетворення (часто недоступну на природних ростових послідовностях), можуть служити зіставлення споріднених форм з різним ступенем педоморфного недорозвитку (Шишкін, 1973). Наступним обширним джерелом є зміни, пов'язані з акреційним ростом скелета, тобто таким, що йде шляхом приросту нових його частин без резорбції попередніх. Такий тип скелетоутворення тією чи іншою мірою відомий у більшості груп безхребетних. Досить згадати розвиток черепашки в головоногих молюсків — джерело хрестоматійних прикладів гетерохроній і рекапітуляцій у давніх організмів. Близьким до акреції є ріст скелета шляхом додавання нових елементів, як це властиво голкошкірим.
Окремим варіантом акреції є ріст колоніальних організмів (коралів, моховаток, граптолітів та ін.), де об'єктом вивчення можуть бути як життєві цикли особин, що складають колонію (гістерогенез), так і її розвиток у цілому (астогенез). При цьому хід історичних змін астогенезу може оцінюватися в тих же категоріях — зокрема, таких, як гетерохронії — що й еволюція індивідуального розвитку.
Спеціальним предметом аналізу можуть бути аномалії розвитку, що являють собою природні аналоги експериментальних порушень формоутворення, досліджуваних у сучасних організмів. Для кожної системи розвитку такі відхилення в сукупності виражають її специфіку (“фазовий портрет”), або простір потенційно здійсненних онтогенетичних траєкторій. Воно завжди обмежене, структуроване і є опосередкованим продуктом еволюції нормального розвитку, складаючи його більш широку характеристику (Waddington, 1957; Шишкін, 1988а, 2006, 2010). Еволюційні зміни цього простору передбачають більшу чи меншу подобу рисунка можливих траєкторій розвитку в споріднених таксонів, причому кожна з них у принципі може виявитися в одному випадку стійкою (нормальною), а в іншому — аберантною4.
У викопних організмів приклади аномалій або просто варіацій, що повторюють нормальні особливості споріднених таксонів, так само часті, як і в сучасних (див. наприклад, Шишкін, Макрідін, 1988, рис. 50). Зокрема, в амонітів відомі випадки, коли скульптура черепашки на одному її боці розвинена нормально, а на іншому — за типом, відомим у близького виду (Tasnadi-Kubacska, 1962, фіг. 82). Крім демонстрації прихованих можливостей розвитку, такі випадки проливають також світло на особливості формоутворення, що не вловлюються у слідах типового онтогенезу. Так, у даному прикладі самоочевидне автономне функціонування мантійного епітелію (що формує орнамент) по двох боках від осі симетрії черепашки.
Іноді аномалії дозволяють фіксувати ранні особливості нормального морфогенезу, які в типовому випадку вже не виражені на стадії формування твердого скелета. Наприклад, у тріасових амфібій-тупілакозаврид компоненти ди-плоспондильних хребців окостенівають у вигляді дисків, що не зберігають слідів розділення на два антимери. Однак ця вихідна парність може виявлятися при неправильному злитті хребців — у випадках, коли окремі диски представлені лише однією половиною.
Усі ці сліди формоутворювальних процесів дають основу для документування еволюційних закономірностей, перелічених вище. Розглянемо деякі приклади.
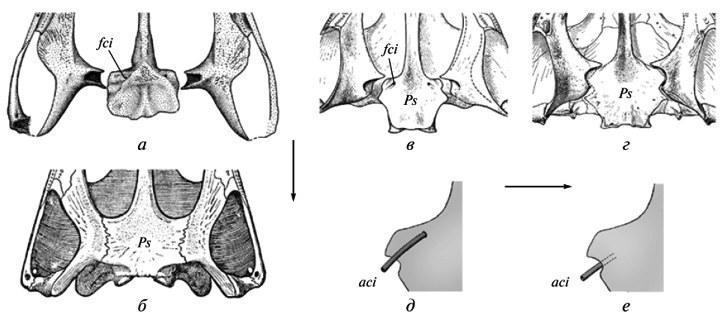
Рис. 3. Перехід від поверхневого розташування внутрішніх сонних артерій (а, в, д) до їхнього поздовжнього вростання в основу черепа (б, г, е) в еволюції темноспондильних амфібій. Основа черепа (а-г) і частина парасфеноїда (д, е) з піднебінного боку: а — примітивний стан з поверхневим розташуванням слідів артерій (Onchiodon); б — похідний стан (Benthosuchus); в, г — примітивний стан у тримерорахоїда Dvinosaurus primus і просунутий у D. еgregius; д — реконструкція ділянки артерії в області її входження в череп у Dvinosaurus primus; е — те саме у D. еgregius. Позначення: aci — внутрішня сонна артерія; fci — вхідний отвір артерії; Ps — парасфеноїд. Стрілки показують напрямок перетворень (а — за: Boy, 1990, зі змінами; б — за: Бистров, Єфремов, 1940; в-д — за: Шишкін, 1973).
Вихідна нестійкість еволюційних змін. Цей феномен проявляється в різних аспектах еволюції черепа давніх темноспондильних амфібій. Зокрема, консолідація його піднебінних покривних окостенінь призвела до зміни топографії внутрішніх сонних артерій. Вихідно, перед впровадженням в ендокраній, вони розташовувалися відкрито на піднебінні (рис. 3, а, д); але потім їхні канали виявилися цілком включеними в парасфеноїд у передвушній області — стан, типовий для тріасових форм (рис. 3, б, е). Перехід від першого плану будови до другого протікав у різних лініях швидко; тому він зазвичай не залишає слідів на доступних для вивчення вікових стадіях. Але в розвитку в ранніх форм другого типу (пермський Acanthostomatops: Witzmann, Schoch, 2006, фіг. 5) варіабельні рекапітуляції вихідних співвідношень можуть зберігатися. Мінливість перехідного дорослого стану (асиметрія вхідних отворів артерій) відома в пермського мелозаврида Tryphosuchus. Нарешті, непряме вираження нестійкості переходу до просунутого типу відзначається в неотенічних двинозаврид, де має місце неоднозначність дорослого стану на родовому рівні. А саме, у роду Dvinosaurus два альтернативних типи розташування артерій, що в нормі характеризують різні еволюційні етапи, виступають у ранзі лише одного з видових відмінностей (рис. 3, в-е) — на тлі збереження примітивної конструкції черепа. Ці співвідношення складають типовий приклад “архаїчного різноманіття” (Мамкаєв, 1968) — феномену, що характеризує ранні, не усталені етапи становлення нових планів організації. Хоча власне двинозавриди є термінальною пізньопермською групою, вони, найімовірніше, дали початок деяким лавразіатським тріасовим “брахіопідам”.
Інший подібний приклад — це втрата в різних лініях пермокарбонових темноспондилів між-скроневої кістки в черепному даху (шляхом злиття її в нормі із задньолобовою). Проміжні етапи цього процесу не виявлені. На найбільш ранніх стадіях онтогенезу, доступних для вивчення (початок окостеніння), його нормальні рекапітуляції вже втрачені — навіть у форм, вивчених за досить повними віковими серіями, таких як бранхіозаври. Проте, як рідкісна варіація ці кістки можуть іноді зберігати самостійність у ювенільних особин — на одному або обох боках черепа (рис. 4, а, б). Іншими словами, їхні мезенхімні закладки зберігалися в ранньому розвитку, іноді досягаючи окостеніння при випадкових затримках або порушеннях процесів злиття (Boy, 1988, фіг. 2B; Schoch, 2004, фіг. 1А, “po”).
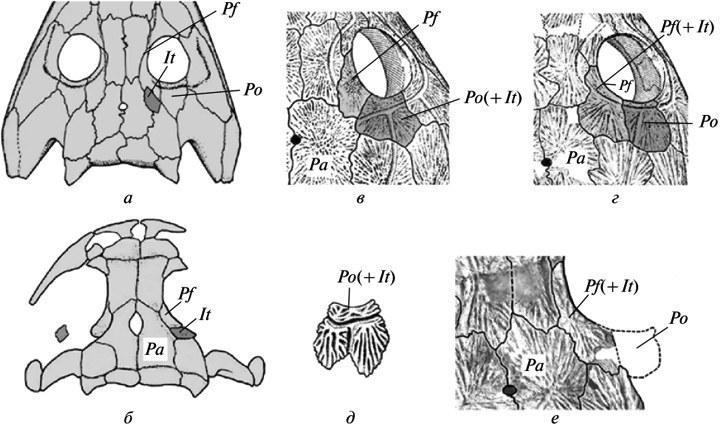
Рис. 4. Нестійкість ранніх етапів втрати міжскроневої кістки в еволюції даху черепа в темноспондильних амфібій: а — збереження кістки у вигляді варіації в еріопоїда Sclerocephalus; б — у бранхіозавра Apateon; в — злиття із заочноямковою кісткою в одного з видів Dvinosaurus (D. primus); г — злиття із задньолобовою кісткою у виду D. еgregius; д — збереження слідів відокремленості intertemporale у складі комплексної заочноямкової кістки в молодої особини D. primus; е — злиття intertemporale із задньолобовою кісткою як варіація в того ж виду. Позначення: It — intertemporale; Pa — parietale; Pf — postfrontale; Pf(+It) — злиті postfrontale і intertemporale; Po — postorbitale; Po(+It) — злиті postorbitale і intertemporale (а — за: Boy, 1988, зі змінами; б — за: Schoch, 2004, зі змінами; в, г — за: Шишкін, 1973; д — за: Bystrow, 1938).
Ця вихідна нестійкість еволюційної зміни, затемнена неповнотою її документації на тлі швидких перетворень, знову набагато виразніше демонструється в неотенічного Dvinosaurus, де зміни протікали повільніше. Збереження тут intertemporale на ранніх стадіях розвитку видно з присутності слідів його окремого центру окостеніння в пізніх ювенільних особин, навіть незважаючи на втрату самостійності кістки (рис. 4, д). Особливо цікаво те, що у вихідного виду D.primus остання, як правило, асимілюється незвичайним для Temnospondyli шляхом, за рахунок злиття із заочноямковою кісткою (рис. 4, в). При цьому як варіація трапляється й альтернативне злиття із задньолобовою кісткою, яке стає нормою в наступного виду D.egregius (рис. 4, г, е). Усе це змушує гадати, що різноспрямованість і нестабільність подій на шляху до кінцевої втрати intertemporale була насправді типова для даного перетворення в історії групи Temnospondyli в цілому. Очевидно, такий же шлях становлення є закономірним для будь-яких еволюційних новшеств. Останнє, власне, і передбачається принципом “архаїчного різноманіття”.
Стабілізація еволюційних придбань. Прояви підвищеної мінливості в ранній історії еволюційних новопридбань уже самі по собі вказують на поступовість їхньої стабілізації. Це стосується як прикладів, наведених вище, так і багатьох інших, відомих на викопному матеріалі (Шишкін, 1988а, с. 168; 1988б, с. 193). Ще частіше спостережуваним наслідком стабілізаційних процесів є прискорення розвитку нових особливостей, що приводить до появи їх на більш ранніх онтогенетичних стадіях (див. розділ II).
Обидві названі закономірності передбачають перебудову формоутворення, але можливості прямої оцінки її механізмів за викопними рештками досить обмежені. Один із нечастих випадків, що дозволяють побачити автономізацію морфогенетичного процесу від екзогенного фактора, що первинно його індукував, був наведений у XIX ст. А. Хаєттом для еволюції черепашки палеозойських наутилоідей. Було показано, що поява в зростаючої черепашки вдавленості на дорсальному боці вихідно викликалася її щільним контактом із попереднім обертом. Але в пізніших форм ця риса вже не виявляє такої залежності і фіксується безпосередньо з початкових стадій росту, ще не зачеплених змиканням обертів (рис. 5). Подібні прояви автономізації розвитку від вихідних індукуючих факторів добре відомі й для еволюції сучасних морфогенезів (Шишкін, 1988б, с. 176).
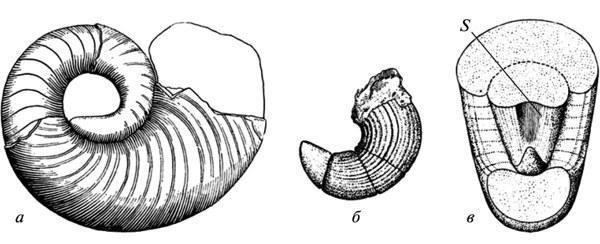
Рис. 5. Автономізація розвитку поздовжньої вдавленості в стінці черепашки в палеозойських наутилоідей: а — рання форма (ознака з'являється в результаті змикання нового оберту черепашки з попереднім); б, в — пізніша форма (поява ознаки випереджає змикання). Позначення: s — поздовжня вдавленість на дорсальній стінці початкового оберту черепашки (за: Hyatt, 1893 з Gould 1977, зі змінами).
Зміна формоутворювальних залежностей може бути також проілюстрована змінами в розвитку черепних кісток, що несуть органи бічної лінії, при переході від риб до давніх амфібій. У кистеперих предків тетрапод, як і в більшості сучасних кісткових риб, плакоди органів бічної лінії, безсумнівно, служили індукторами для закладки “канальних” покривних кісток, що розвивалися навколо них. Відповідно, ланцюжки сейсмосенсорних органів голови розташовувалися в цих риб у каналах, що проходять крізь центри формування відповідних кісток. У палеозойських амфібій цей зв'язокбув утрачений, тобто кістки закладалися вже незалежно від системи бічної лінії. Остання формувалася більш поверхнево в епідермісі і могла вступати в контакт із кістками лише на більш пізніх стадіях розвитку. Координація цих двох морфогенезів почасти зберігалася, оскільки жолобки бічної лінії все ще в основному проходили над центрами розвитку колишніх “канальних” кісток. Однак ця залежність була вже не настільки жорсткою, допускаючи зміщення окремих жолобків на інші кістки (пор. Шишкін, 1973, рис. 65, с. 170).
Рекапітуляції анцестральних рис (палінгенези), збережені в примітивних типах розвитку. Серед особливостей дорослої організації, властивих давнім хребетним, багато які або взагалі не залишили слідів у сучасних онтогенезах, або ці сліди не піддаються однозначному тлумаченню. Але в ближчих нащадків давніх форм такі особливості могли закарбовуватися в розвитку набагато виразніше. Це, зокрема, видно з таких даних щодо розвитку давніх амфібій.
Як показано Е. Ярвіком, парасфеноїд у тетрапод виник шляхом об'єднання власне парасфеноїда їхніх кистеперих предків (що підстилав трабекулярну область черепа) з позадулежачими парними привушними пластинками, що належали екзоскелету гіоїдної дуги (рис. 6, а). У розвитку сучасних тетрапод сліди парності заднього відділу комплексного парасфеноїда втрачені, включаючи й примітивних Urodela (пор. Лебєдкіна, 1979). У палеозойських темноспондильних амфібій до початку окостеніння цей стан у нормі також не залишав слідів; але він, безсумнівно, зберігався на стадії мезенхімних закладок піднебінного екзоскелета. Це доводиться варіаціями, при яких фаза розщепленості заднього відділу парасфеноїда індивідуально пролонгувалася, зберігаючись на стадіях окостеніння піднебіння. Такі приклади нерідкі в розвитку в бранхіозаврів (рис. 6, б, в). Серед еріопоїдів аналогічна затримка в одному випадку зафіксована вже у цілком сформованої ювенільної особини з окостенілим ендокранієм (рис. 6, г).
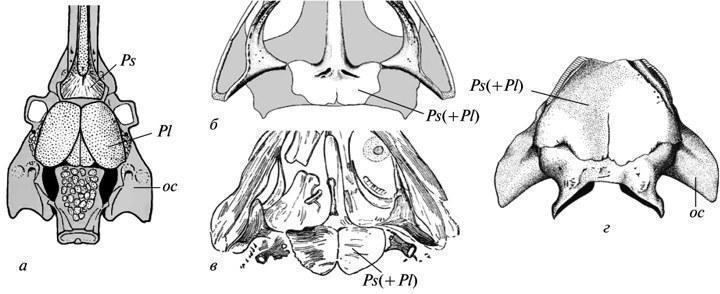
Рис. 6. Сліди рекапітуляції привушних пластин піднебіння кистеперих риб у розвитку парасфеноїда в темноспондильних амфібій. Черепні структури знизу (а-г): а — кросоптеригія Eusthenopteron (потилично-вушний і частина етмосфеноїдного відділів ендокранія); б, в — бранхіозавр Apateon і близька до нього форма (задня частина піднебіння); г — ювенільний еріопоїд “Actinodon” (потилично-вушний відділ ендокранія). Позначення: oc — вушна капсула; Pl — привушна пластина; Ps — парасфеноїд; Ps(+Pl) — комплексний парасфеноїд тетрапод (а — за: Jarvik, 1954; б — за: Boy, 1978; в — за: Watson, 1963; г — за: Boy, 1971; усі зі змінами).
До кола давніх проблем еволюційної морфології, що обговорюються в контексті даних розвитку, належить питання про гомологію слухової кісточки в безхвостих амфібій. На відміну від рептильного stapes, що розвивається з гіоїдної дуги, у безхвостих ця кісточка виникає зі стінки вушної капсули (або самостійно), поза зв'язком із гіоїдом. Тому багатьма дослідниками вона оцінювалася як новоутворення (“otostapes”), що виникло незалежно від “hyostapes” рептилій. Тим часом, у прямих предків безхвостих — палеозойських батрахоморф (Temnospondyli) картина розвитку stapes, будучи набагато архаїчнішою, ясно вказує на гіоїдне походження цього елемента. Останній у дорослих форм за будовою і топографією цілком подібний до свого гомолога в Anura (рис. 7, а, б, к, л); гіостапедіальний зв'язок міг тут зберігатися лише у вигляді рудиментарного лігамента. Але на ранніх стадіях, а в неотенічних форм — іноді й у дорослому стані, вона була представлена потужним хрящовим інфрастапедіальним відростком stapes, що мав широку основу і рекапітулював стовбур гіомандибули кистеперих предків (рис. 7, в—ж).
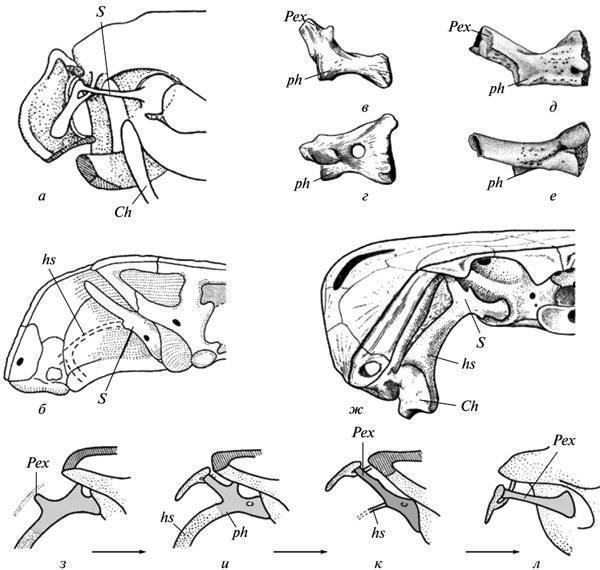
Рис. 7. Рекапітуляція гіостапедіального зв'язку в онтогенезі давніх батрахоморф (б—к): а — область середнього вуха в сучасних безхвостих (Rana); б — вушна область потилиці в темноспондилів (Lydekkerina); в—е — слухова кісточка в цій же групі в неотенічних форм (Dvinosaurus: в, г) і на ранніх стадіях розвитку (Sclerocephalus: д, е), вид ззаду (в, д) і зверху (г, е); ж — реконструкція гіостапедіального зв'язку в черепі Dvinosaurus; з—л — реконструкція шляху перетворення гіомандибули в слухову кісточку в батрахоморф — від найдавніших амфібій (з, и) до типово темноспондильної конструкції (к) і співвідношень у безхвостих (л). Позначення: Ch — ceratohyale; hs — гіостапедіальний хрящ або лігамент; Pex — processus externus (extrastapedialis); ph — processus hyoideus (infrastapedialis); S — stapes (a — за: Eaton, 1959; б — за: Parrington, 1948; д, е — за: Boy, 1988; в, г, ж—л — за Шишкіним, 1973, зі змінами).
Однією з дискусійних особливостей ембріогенезу сучасних амніот є ресегментація їхнього хребта. Закладки тіл хребців виникають тут із двох суміжних половин склеротомів сусідніх сегментів, у результаті чого кожен хребець розвивається у зв'язку з двома послідовними міотомами (рис. 8, а—в). Походження цієї перебудови не піддається тлумаченню на основі порівняння розвитку в сучасних тетрапод. У рецентних амфібій вона затемнена пізнім формуванням склеротомів і тому часто ставиться під сумнів.
Ключ до розуміння природи названого феномену майже напевно дають співвідношення в давніх апсидоспондильних (складнохребцевих) амфібій, а саме, у все тієї ж їхньої батрахоморфної гілки — Temnospondyli. У цих форм напівцентри тіл хребців могли об'єднуватися у функціональні одиниці двома альтернативними шляхами — внутрішньосегментально і транссегментально, тобто шляхом інтеграції напівцентрів із сусідніх метамерів (рис. 8, г-ж). Перехід від першого типу до ресегментованого другого відбувався в історії Temnospondyli неодноразово, здійснюючись у пізньому розвитку; при цьому в неотенічного Dvinosaurus присутні обидва варіанти, представлені різними видами. Іноді (у тріасових метопозаврів) може спостерігатися перехід від вихідного варіанта до похідного в межах одного й того ж хребетного стовпа (Шишкін, 1987).
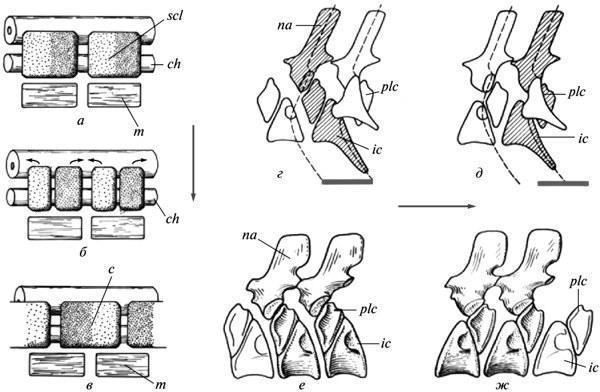
Рис. 8. Ембріональна ресегментація склеротомів в амніот як відображення рекомбінації напівцентрів хребця в давніх апсидоспондильних амфібій: а—в — стадії ресегментації в сучасних амніот (схема); г, д — внутрішньосегментальний і транссегментальний хребцеві метамери в темноспондильних амфібій (схема); е — внутрішньосегментальні хребці в Dvinosauгus egregius; ж — транссегментальні хребці в D. purlensis. Позначення: с — закладка тіла хребця; ch — хорда; ic — інтерцентр; m — міотом; na — невральна дуга; plc — плевроцентр; scl — склеротом; стрілки показують напрямок ресегментації; сірі лінії — протяжність функціонального сегмента (а—в — за: Romer, 1968; г—ж — за: Шишкін, 1987).
Зважаючи на величезний пробіл між сучасними амніотами і давніми амфібіями, а також на той факт, що Temnospondyli не є предками амніот, можна, звичайно, поставити під сумнів спадкоємність двох розглянутих типів ресегментації. Однак більш обґрунтованим є припущення, що в ранніх наземних рептиліоморфних амфібій (предків амніот) ресегментація починалася тим же шляхом, що й у батрахоморф, але потім прискорення цього процесу призвело до його ембріоналізації ще в палеозої. Опосередковано на користь цього погляду говорить також присутність обох “батрахоморфних” типів інтеграції хребців у кистеперих предків тетрапод (Шишкін, 1987).
У рамках цієї точки зору ми повинні зробити висновок, що якщо вихідно об'єктом ресегментації служили кісткові або хрящові компоненти дефінітивного апсидоспондильного хребця, то її прискорення і поглиблення привели до стану, де її прямим субстратом став уже матеріал ембріональних склеротомів. Ця перебудова зробила весь процес незалежним від формування хребцевих напівцентрів, з яких в амніот практично зберігся лише один (плевроцентр). У сумі це дає наочний приклад еквіфінальності стабільного еволюційного придбання, коли, залишаючись історично незмінним за своїм головним результатом, воно докорінно змінює спосіб свого здійснення.
Усі розглянуті приклади морфогенезів у давніх організмів (оцінені шляхом зіставлення між собою або із сучасними типами розвитку) дозволяють висвітлити деякі головні закономірності історичної стабілізації формоутворення. Остання простежується тут як у вигляді слідів початкової нестійкості майбутніх типових рис організації, так і на прикладах великомасштабних історичних змін, що приводять до якісної перебудови механізму розвитку таких рис. Як показано вище, ці феномени є характерними й очікуваними проявами цілісної еволюції онтогенезу як системи.
СПИСОК ЛІТЕРАТУРИ
Бистров А.П., Єфремов І.А. Benthosuchus sushkini Efr.. — лабіринтодонт з еотріасу р. Шарженги. // Тр. Палеонтол. ін-ту АН СРСР. 1940. Т. 10. Вип. 1. 152 с.
Гертвіг О. Сучасні спірні питання біології. Вип. 1. Еволюція чи епігенез. М., 1895. 125 с.
Гілберт С.Ф., Опіц Д.М., Реф Р.А. Новий синтез еволюційної біології і біології розвитку // Онтогенез. 1997. Т. 28. № 5. С. 325-343.
Голубовський М.Д. Неканонічні спадкові зміни // Природа. 2001. № 8. С. 3-9.
Інге-Вечтомов С.Г. Система генотипу // Фізіологічна генетика. Л.: Медицина, 1976. С. 57-113.
Корочкін Л.І. Біологія індивідуального розвитку. М.: Вид-во МДУ, 2002. 263 с.
Лебєдкіна Н.С. Еволюція черепа амфібій. М.: Наука, 1979. 283 с.
Макрідін В.П., Шишкін М.А. Вивчення онтогенезу вимерлих організмів // Сучасна палеонтологія / За ред. В.В. Меннера, В.П. Макрідіна. М.: Надра, 1988. Т. 1. С. 197-211.
Мамкаєв Ю.В. Порівняння морфологічних відмінностей у нижчих і вищих групах одного філогенетичного стовбура // Журн. загальної біол. 1968. Т. 29. С. 48-55.
Мейєн С.В. Проблеми філогенетичної класифікації організмів // Сучасна палеонтологія / За ред. В.В. Меннера, В.П. Макрідіна. М.: Надра, 1988. Т. 1. С. 497-511.
Морган Т. Структурні основи спадковості. М.-Л.: Держвидав, 1924. 310 с.
Руженцев В.Є. Принципи систематики, система і філогенія палеозойських амоноідей // Тр. Палеонтол. ін-ту АН СРСР. 1960. Т. 83. С. 1-331.
Свєтлов П.Г. Про цілісний і елементаристичний методи в ембріології // Арх. анат. гістол. ембріол. 1964. Т 46. № 4. С. 3-26.
Свєтлов П.Г. Онтогенез як цілеспрямований (телеономічний) процес // Арх. анат. гістол. ембріол. 1972. Т. 63. № 8. С. 5-16.
Свєтлов П.Г. Фізіологія (механіка) розвитку. Л.: Наука, 1978. Т. 1. 279 с.; Т. 2. 262 с.
Шишкін М.А. Морфологія давніх земноводних і проблеми еволюції нижчих тетрапод // Тр. Палеонтол. ін-ту АН СРСР. 1973. Т. 137. 260 с.
Шишкін М.А. Еволюція давніх амфібій // Тр. Палеонтол. ін-ту АН СРСР. 1987. Т. 225. 142 с.
Шишкін М.А. Еволюція як епігенетичний процес // Сучасна палеонтологія / За ред. Меннера В.В., Макрідіна В.П. М.: Надра, 1988а. Т. 2. С. 142-169.
Шишкін М.А. Закономірності еволюції онтогенезу // Сучасна палеонтологія. / За ред. Меннера В.В., Макрідіна В.П. М.: Надра, 1988б. Т. 2. С. 169-209.
Шишкін М.А. Індивідуальний розвиток і уроки еволюціонізму // Онтогенез. 2006. Т. 37. № 3. С. 179-198.
Шишкін М.А. Еволюційна теорія і наукове мислення // Палеонтол. журн. 2010. № 6. С. 3-17.
Шишкін М.А., Макрідін В.П. Палеопатологічні і палеотератологічні дослідження // Сучасна палеонтологія / За ред. Меннера В.В., Макрідіна В.П. М.: Надра, 1988. Т. 1. С. 219-229.
Шмальгаузен І.І. Організм як ціле в індивідуальному та історичному розвитку. М.: Наука, 1982. 228 с. Alberch P. Developmental constraints in evolutionary processes // Life Sci. Res. Rep. 1982. № 22. P. 313-332.
Baer K. Ueber Entwicklungsgeschichte der Thiere. T I. Koenigsberg: Kupfertaf, 1828. 271 S.
Beer G.R. de. Embryos and ancestors. Oxford: Clarendon Press, 1971.191 p.
Boy J. Ein bemerkenswerter Schädelrest eines unterpermischen Labyrinthodontiers (Amphibia) aus dem Saargebiet // Abh. hess. Land.-Amt. Bodenforsch. 1971. Bd 60. S. 31-43. Boy J. Die Tetrapodenfauna (Amphibia, Reptilia) des saar- pfalzischen Rothliegenden (Unter-Perm, SW Deutschland). 1.Branchiosaurus // Mainzer geowiss. Mitt. 1978. Bd 7. S. 27-76.
Boy J.A. Über einige Vertreter der Eryopoidea (Amphibia: Temnospondyli) aus dem europäischen Rotliegend (?höchstes Karbon - Perm). 1. Sclerocephalus // Paläontol. Z. 1988. Bd 62. Hf. 1/2. S. 107-132.
Boy J. Über einige Vertreter der Eryopoidea (Amphibia: Temnospondyli) aus dem europäischen Rotliegend (?höchstes Kabon-Perm). 3. Onchiodon // Paläontol. Z. 1990. Bd 64. S. 287-312.
Bystrow A.P. Dvinosaurus als neotenische Form der Stego- cephalen // Acta zool. 1938. Bd 19. S. 209-295.
Cairns J, Overbaugh J, Miller S. The origin of mutants // Nature. 1988. V. 335. P. 142-145.
Driesch H. The science and philosophy of the organism. L.: Black, 1908. V. 1. 329 p.; V. 2. 381 p.
Eaton T.H. The ancestry of modern Amphibia. A review of the evidence // Univ. Kansas Publs, Mus. Natur. History. 1959. V. 12. P. 155-180.
Goldschmidt R. The material basis of evolution. New Haven: Yale Univ. Press, 1940. 436 p.
Gould S.J. Ontogeny and phylogeny. Cambridge: Belknap Press, 1977. 501 p.
Gould S.J. Change in developmental timing as a mechanism of macroevolution // Evolution and development / Ed. Bonner J.T Berlin: Springer, 1982. P. 337-346.
Gurwitsch A.G. Über den Begriff des embryonalen Feldes // Arch. Entw. Mech. 1922. Bd 51. S. 388-415.
Hall B.G. Spontaneous point mutations that occur more often when advantageous than when neutral // Genetics. 1990. V. 126. № 1. P. 5-16.
Jarvik E. On the visceral skeleton in Eusthenopteron with a discussion of the parasphenoid and palatoquadrate in fishes // Kungl. Svenska Vbtenskaapsakad. Handl. 5 Ser. 1954. Bd 5. № 1. P. 1-104.
Maderson P.F.A., Alberch P., Goodwin B.C. et al. The role of development in macroevolutionary change // Evolution and Development. Ed. Bonner J.T. Berlin: Springer, 1982. P. 279-312.
Parrington F.R. The Problem of the Classification of Reptiles // J. Linnean Soc. London, Zool. 1958. V. 44. № 295. P. 99-115.
Romer A.S. The Vertebrate Body, 3rd ed. Philadelphia: W.B. Saunders, 1962.
Roux W. Gesammelte Abhandlungen über Entwicklungsmechanik der Organismen. Leipzig: Engelmann, 1896. vols. 1 and 2.
Schoch R. Skeleton Formation in the Branchiosauridae: A Case Study in Comparing Ontogenetic Trajectories // J. Vertebr. Paleontol. 2004. V. 24. № 2. P. 309-319.
Tasnadi-Kubacska A. Paläopatologie. Jena: Fischer, 1962.
Waddington C.H. The strategy of the Genes: A Discussion on Some Aspects of Theoretical Biology. London: Allen and Unwin, 1957.
Watson D.M.S. On the Growth Stages in Branchiosaurus // Palaeontology. 1963. V. 6. P. 540-553.
Witzmann F., Schoch R. Skeletal Development of the Temnospondyl Acanthostomatops vorax from the Lower Permian Döhlen Basin of Saxony // Trans. Roy. Soc. Edinburgh. 2006. V. 96. P. 365-385.
Systemic Grounds of Morphogenesis and Their Manifestation in the Fossil Record
M. A. Shishkin
Morphogenetic mechanisms form an integrity purposed to secure the due course of ontogeny towards the adult norm. This purposefulness is a principal feature of living organization that requires an explanation. Each step in evolution is a compelled reorganization of the ontogenetic system towards a new ultimate goal (system’s equilibrium state), i.e., a new norm. An increase in stability of realization of the latter, caused by natural selection, results in progressive remodeling of ontogeny of the evolutionary novelty which spreads in succeeding life cycles towards the earlier stages. As a result, the difference between current ontogenetic record of the novelty and the ancestral developmental pattern tends to change more and more from the prolonging (recapitulative) to divergent one, corresponding to Baer’s law. In the fossil record, the changes caused by these stabilizing processes result in a number of manifestations, such as initially labile expression of new acquisitions, the remodeling of their morphogenesis with time and the predominant maintenance of ancestral traits in late ontogeny of the closest descendants.
Keywords: morphogenesis, ontogeny, purposefulness, evolution, stabilization, fossils.
1 Але в способі тлумачення еволюційних подій класична преформістська ідеологія генетики по суті не зазнала змін. Про це говорять незліченні приклади пояснення нових ознак у сучасних і викопних організмів ефектами відповідних мутацій. Назад до тексту
2 Подібні явища, як і гетерохронії взагалі, вважаються в еволюційній літературі одним із головних механізмів макроеволюції (див., наприклад, Gould, 1977). Це звичайний приклад змішування причин і наслідків в еволюційному мисленні. Насправді йдеться лише про один із проявів системної перебудови онтогенезу, який вловлюється дослідником у вигляді неузгодженості в часі окремих процесів розвитку. Назад до тексту
3 Останнє стосується рівною мірою і стабілізації педоморфних змін, тобто прогресуючої втрати дорослих анцестральних рис при їхньому недорозвитку (Шишкін, 1988б). Назад до тексту
4 Спроби осмислення різних аспектів цього феномену (не пов'язані з розумінням розвитку як системного об'єкта еволюції) відображені в безлічі узагальнень. Це паралелізм мутаційної і модифікаційної мінливості (або зв'язок між нормою і аномалією в різних таксонів), гомологічна мінливість, а також поняття, формалізовані С.Мейєном (1988) як “рефрени”, “транзитивний поліморфізм” або “правило Кренке”. Цей же феномен під назвою “розвиткових обмежень” (developmental constraints) нині привертає зростаючу увагу як одна з ключових закономірностей, що визначають хід еволюційного процесу (Gould, 1982; Alberch, 1982). При цьому він нерідко розуміється як щось самостійне щодо природного добору. Назад до тексту